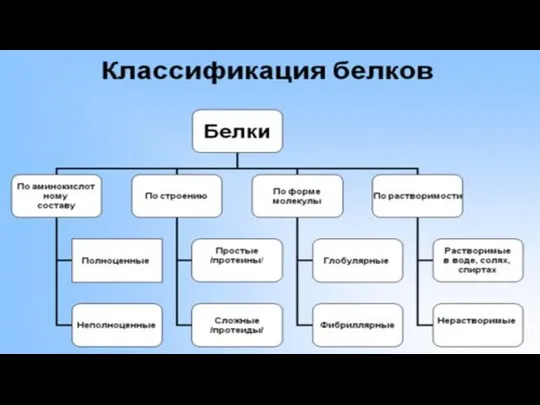
- Белки и ферменты

Содержание
- 2. План План: 1)Введение; 2)Синтез белка; а)транскрипция б)трансляция в)посттрансляционные изменения 3)Заключение; 4)Список использованной литературы.
- 3. Введение Введение Белки́ (протеи́ны, полипепти́ды]) — высокомолекулярные органические вещества, состоящие из альфа-аминокислот, соединённых в цепочку пептидной
- 7. Синтез белка — сложный многоступенчатый процесс, представляющий цепь синтетических реакций, протекающих по принципу матричного синтеза. Транскрипция
- 8. Участок ДНК, на котором происходит транскрипция (транскриптон, оперон), состоит из трех частей: промотора, гена (в случае
- 9. Трансляция (англ. translation – перевод) – это биосинтез белка на матрице мРНК. После переноса информации с
- 10. Для инициации необходимы мРНК, ГТФ, малая и большая субъединицы рибосомы, три белковых фактора инициации ,метионин и
- 11. Элонгация Для этой стадии необходимы все 20 аминокислот, Элонгация представляет собой циклический процесс. Первый цикл (и
- 12. 3 Фермент транслоказа перемещает мРНК относительно рибосомы таким образом, что первый кодон АУГ оказывается вне рибосомы,
- 13. Терминация Синтез белка продолжается до тех пор, пока рибосома не достигнет на мРНК особых терминирующих кодонов
- 14. После завершения трансляции большая часть белков подвергается дальнейшим химическим модификациям, которые называются посттрансляционными модификациями. Известно более
- 15. Посттрансляционные модификации могут быть как широко распространёнными, так и редкими, вплоть до уникальных. Примером универсальной модификации
- 16. Посттрансляционные модификации делят на: модификации главной цепи; отщепление N-концевого остатка метионина; ограниченный протеолиз — удаление фрагмента
- 17. Заключение Белки играют в питании человека чрезвычайно важную роль, так как они являются главной составной частью
- 19. Скачать презентацию
Слайд 2План
План:
1)Введение;
2)Синтез белка;
а)транскрипция
б)трансляция
в)посттрансляционные изменения
3)Заключение;
4)Список использованной литературы.
План
План:
1)Введение;
2)Синтез белка;
а)транскрипция
б)трансляция
в)посттрансляционные изменения
3)Заключение;
4)Список использованной литературы.

Слайд 3Введение
Введение
Белки́ (протеи́ны, полипепти́ды]) — высокомолекулярные органические вещества, состоящие из альфа-аминокислот, соединённых
Введение
Введение
Белки́ (протеи́ны, полипепти́ды]) — высокомолекулярные органические вещества, состоящие из альфа-аминокислот, соединённых
![Введение Введение Белки́ (протеи́ны, полипепти́ды]) — высокомолекулярные органические вещества, состоящие из альфа-аминокислот,](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/949646/slide-2.jpg)


Слайд 7Синтез белка — сложный многоступенчатый процесс, представляющий цепь синтетических реакций, протекающих по
Синтез белка — сложный многоступенчатый процесс, представляющий цепь синтетических реакций, протекающих по

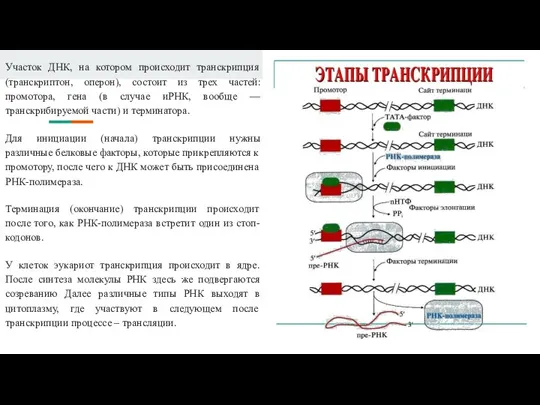
Слайд 8Участок ДНК, на котором происходит транскрипция (транскриптон, оперон), состоит из трех частей:
Участок ДНК, на котором происходит транскрипция (транскриптон, оперон), состоит из трех частей:
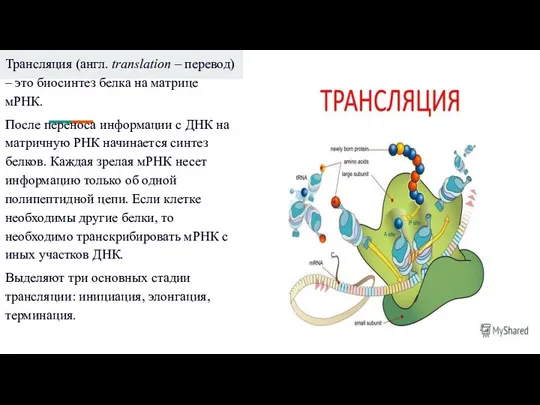
Слайд 9Трансляция (англ. translation – перевод) – это биосинтез белка на матрице мРНК.
После
Трансляция (англ. translation – перевод) – это биосинтез белка на матрице мРНК.
После
Слайд 10Для инициации необходимы мРНК, ГТФ, малая и большая субъединицы рибосомы, три белковых
Для инициации необходимы мРНК, ГТФ, малая и большая субъединицы рибосомы, три белковых

Слайд 11Элонгация
Для этой стадии необходимы все 20 аминокислот, Элонгация представляет собой циклический процесс.
Элонгация
Для этой стадии необходимы все 20 аминокислот, Элонгация представляет собой циклический процесс.

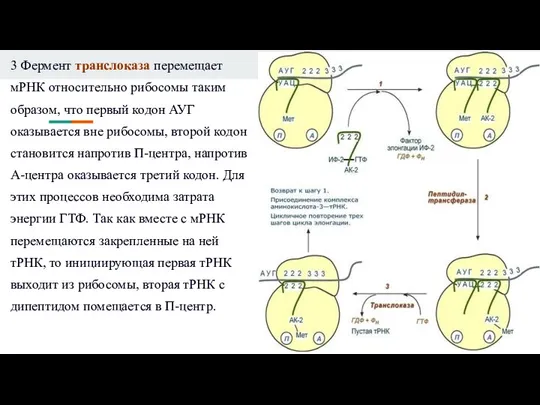
Слайд 123 Фермент транслоказа перемещает мРНК относительно рибосомы таким образом, что первый кодон
3 Фермент транслоказа перемещает мРНК относительно рибосомы таким образом, что первый кодон
Слайд 13Терминация
Синтез белка продолжается до тех пор, пока рибосома не достигнет на мРНК
Терминация
Синтез белка продолжается до тех пор, пока рибосома не достигнет на мРНК

Слайд 14После завершения трансляции большая часть белков подвергается дальнейшим химическим модификациям, которые называются
После завершения трансляции большая часть белков подвергается дальнейшим химическим модификациям, которые называются

Слайд 15Посттрансляционные модификации могут быть как широко распространёнными, так и редкими, вплоть до
Посттрансляционные модификации могут быть как широко распространёнными, так и редкими, вплоть до

Слайд 16Посттрансляционные модификации делят на:
модификации главной цепи;
отщепление N-концевого остатка метионина;
ограниченный протеолиз — удаление
Посттрансляционные модификации делят на:
модификации главной цепи;
отщепление N-концевого остатка метионина;
ограниченный протеолиз — удаление

Слайд 17Заключение
Белки играют в питании человека чрезвычайно важную роль, так как они являются
Заключение
Белки играют в питании человека чрезвычайно важную роль, так как они являются

 Серое вещество спинного мозга
Серое вещество спинного мозга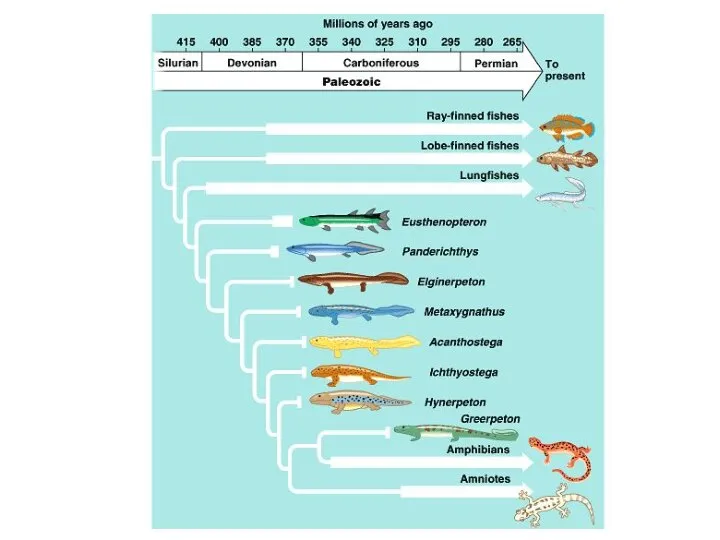 Амфибии. Переход от sarcopterygia к tetrapoda. Panderichthys
Амфибии. Переход от sarcopterygia к tetrapoda. Panderichthys Правило Глогера
Правило Глогера Меры безопасности при применении пестицидов. (Лекция 22)
Меры безопасности при применении пестицидов. (Лекция 22) Обмен веществ (метаболизм)
Обмен веществ (метаболизм) Биологические методы борьбы с вредителями
Биологические методы борьбы с вредителями Kriterii_vida1
Kriterii_vida1 Видообразование. Эволюционные процессы
Видообразование. Эволюционные процессы Формы размножения организмов. Бесполое размножение
Формы размножения организмов. Бесполое размножение Уровни организации живой материи. Клетка наименьшая структурная единица живой материи
Уровни организации живой материи. Клетка наименьшая структурная единица живой материи Жизненный цикл Лягушачьей многоустки – Polystoma integerrimum
Жизненный цикл Лягушачьей многоустки – Polystoma integerrimum Глаз (ophthalmos oculus)
Глаз (ophthalmos oculus) Породы собак
Породы собак Quiz o zwierzętach
Quiz o zwierzętach 1359898
1359898 Презентация на тему Репликация ДНК
Презентация на тему Репликация ДНК  Презентация по биологии на тему_ _Отдел Моховидные. Особенности строения мхов_ (7 клас
Презентация по биологии на тему_ _Отдел Моховидные. Особенности строения мхов_ (7 клас Мейоз
Мейоз Периферическая нервная система, спинномозговые нервы
Периферическая нервная система, спинномозговые нервы Жизнь на Земле
Жизнь на Земле Археи. Общие особенности, экология, эволюционное значение
Археи. Общие особенности, экология, эволюционное значение Папоротникообразные
Папоротникообразные Аконит
Аконит Урок по биологии для 8 класса. Для чего нам сердце?
Урок по биологии для 8 класса. Для чего нам сердце? Плод. Разнообразие и значение плодов. Распространение плодов и семян
Плод. Разнообразие и значение плодов. Распространение плодов и семян Подготовка к ЕГЭ. Задания А1
Подготовка к ЕГЭ. Задания А1 Биомембраны. Функции и общая характеристика строения биомембран. Виды мембранного транспорта
Биомембраны. Функции и общая характеристика строения биомембран. Виды мембранного транспорта Рассказы о животных
Рассказы о животных