- Биология рака

Содержание
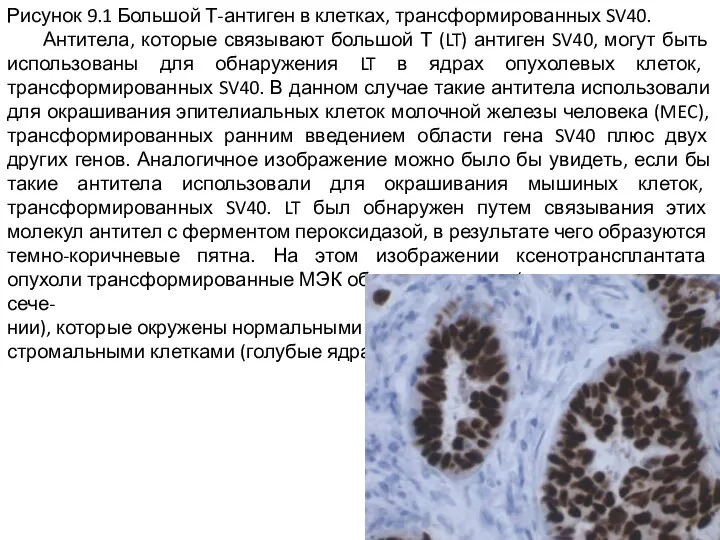
- 2. Рисунок 9.1 Большой Т-антиген в клетках, трансформированных SV40. Антитела, которые связывают большой Т (LT) антиген SV40,
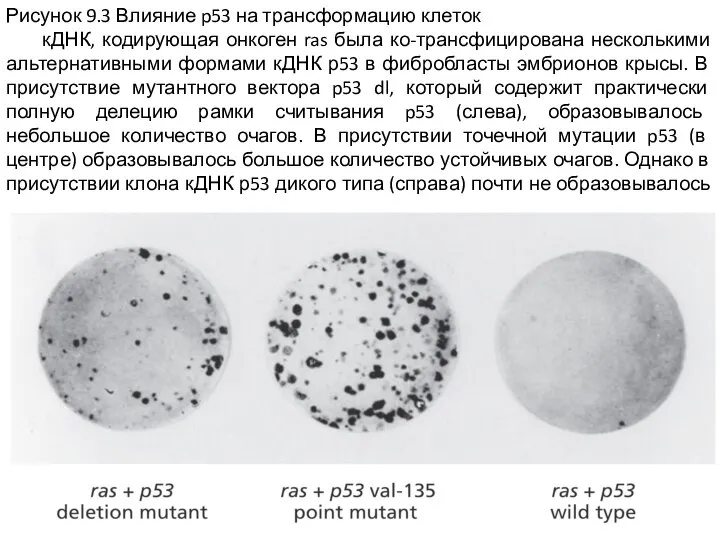
- 3. Рисунок 9.3 Влияние p53 на трансформацию клеток кДНК, кодирующая онкоген ras была ко-трансфицирована несколькими альтернативными формами
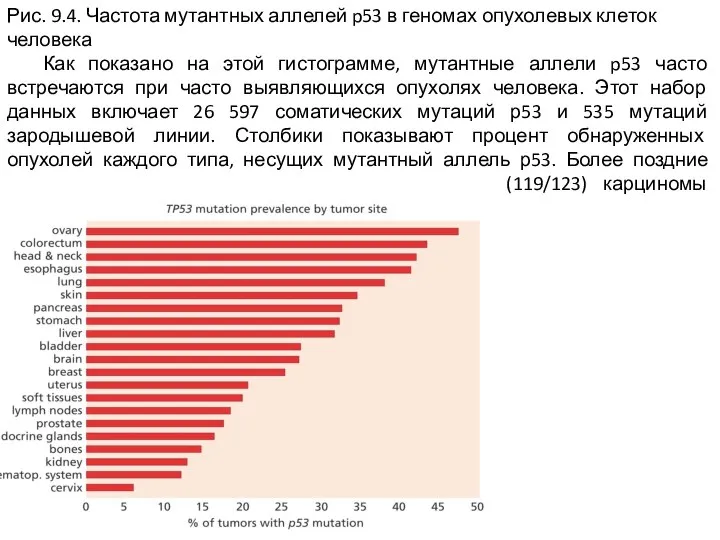
- 4. Рис. 9.4. Частота мутантных аллелей p53 в геномах опухолевых клеток человека Как показано на этой гистограмме,
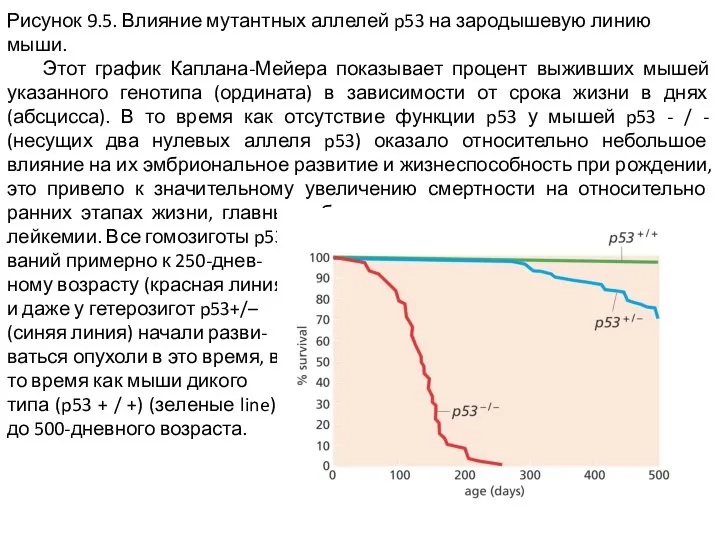
- 5. Рисунок 9.5. Влияние мутантных аллелей p53 на зародышевую линию мыши. Этот график Каплана-Мейера показывает процент выживших
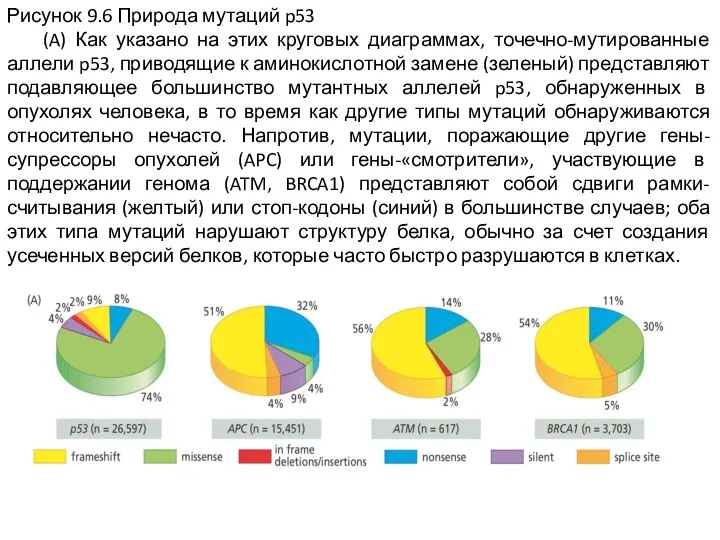
- 6. Рисунок 9.6 Природа мутаций p53 (A) Как указано на этих круговых диаграммах, точечно-мутированные аллели p53, приводящие
- 7. (B) Места в рамке считывания p53 точечных мутаций, вызывающих аминокислотные замены отмечены здесь (вверху). Очевидно, что
- 8. (C) Общая структура тетрамера р53, связанного с ДНК, показана здесь. Четыре ДНК-связывающих домена показаны зеленым и
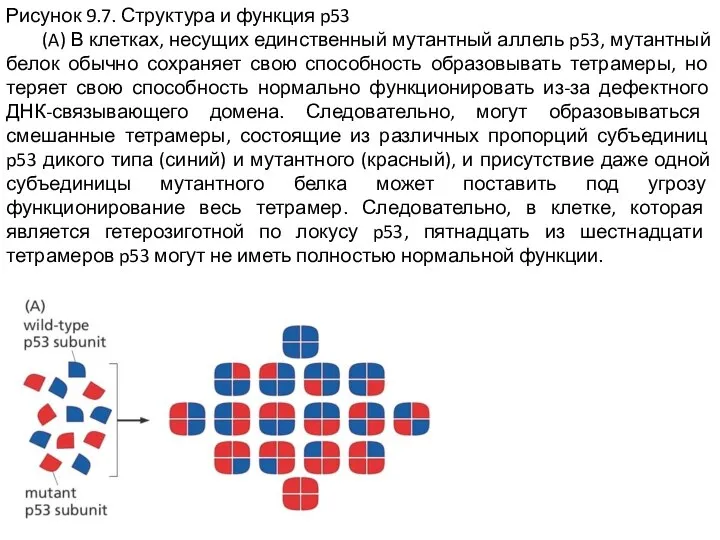
- 9. Рисунок 9.7. Структура и функция p53 (A) В клетках, несущих единственный мутантный аллель p53, мутантный белок
- 10. (B) Возможно, наиболее прямая демонстрация доминантно-негативного режима действия p53 произошла от «вбивания» мутантных аллелей p53 в
- 11. Рисунок 9.8. Сигналы, активирующие р53, и последующие эффекты р53. Исследования функции p53 показали, что различные физиологические
- 12. Рисунок 9.10. Контроль уровней p53 с помощью Mdm2. После того, как концентрации p53 увеличиваются в ответ
- 13. Рисунок 9.15. Контроль апоптоза с помощью ARF (A) Белок p14ARF чело- века, называемый ARF на рисунке
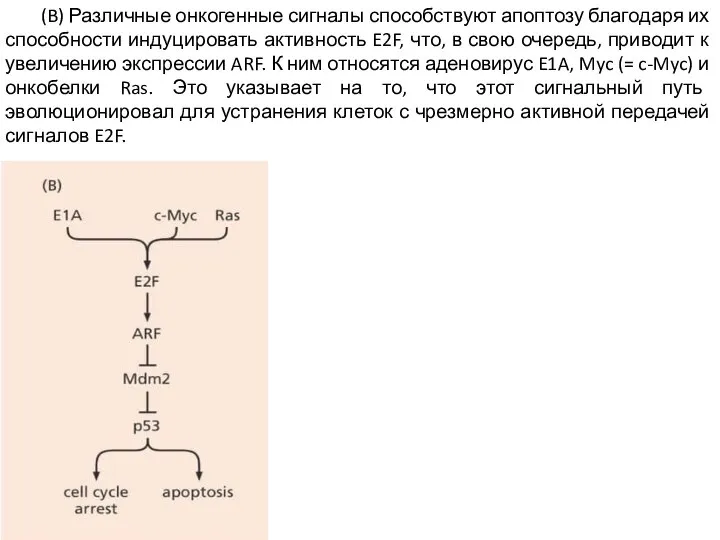
- 14. (B) Различные онкогенные сигналы способствуют апоптозу благодаря их способности индуцировать активность E2F, что, в свою очередь,
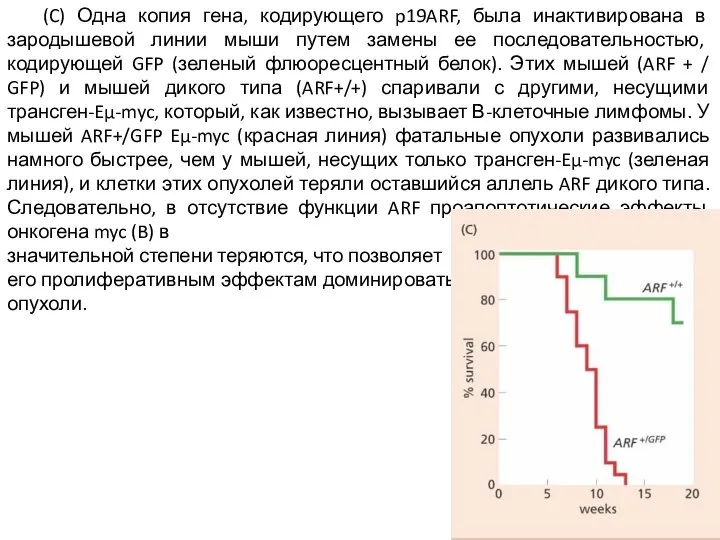
- 15. (C) Одна копия гена, кодирующего p19ARF, была инактивирована в зародышевой линии мыши путем замены ее последовательностью,
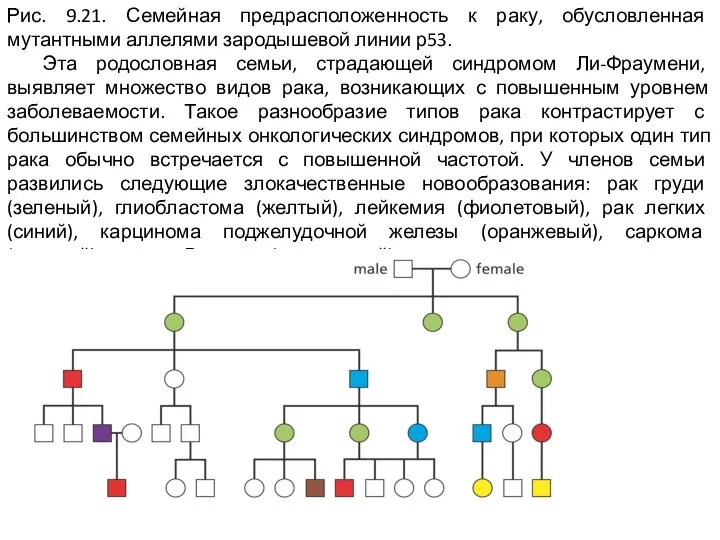
- 16. Рис. 9.21. Семейная предрасположенность к раку, обусловленная мутантными аллелями зародышевой линии р53. Эта родословная семьи, страдающей
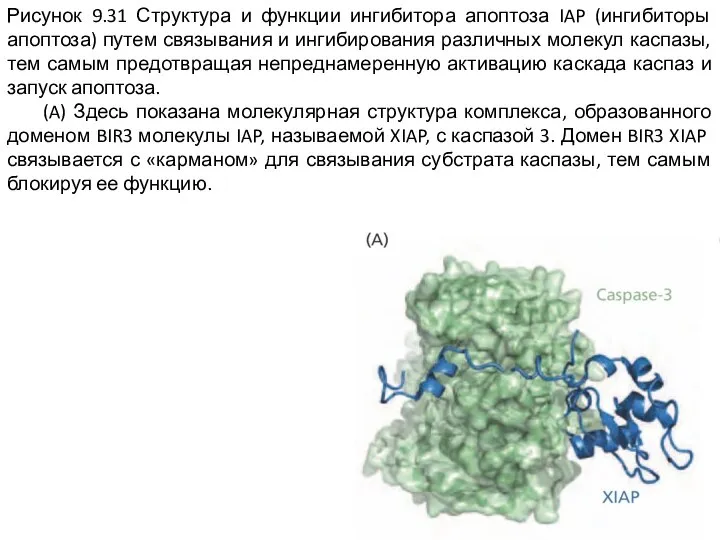
- 17. Рисунок 9.31 Структура и функции ингибитора апоптоза IAP (ингибиторы апоптоза) путем связывания и ингибирования различных молекул
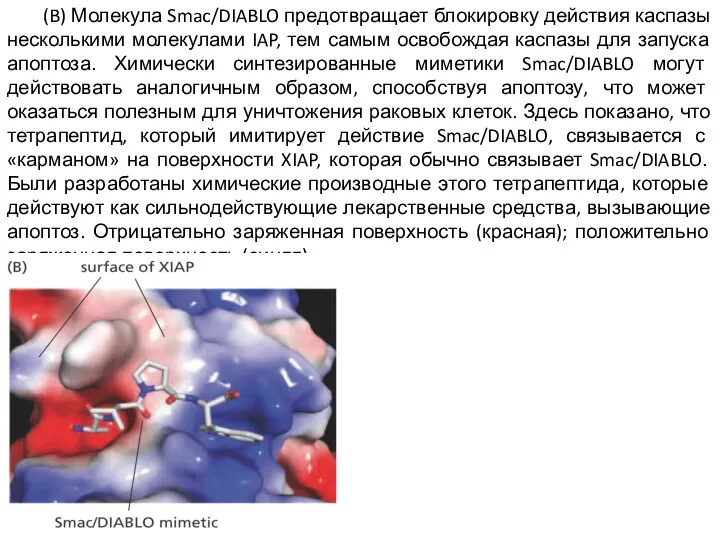
- 18. (B) Молекула Smac/DIABLO предотвращает блокировку действия каспазы несколькими молекулами IAP, тем самым освобождая каспазы для запуска
- 20. Скачать презентацию
Слайд 2Рисунок 9.1 Большой Т-антиген в клетках, трансформированных SV40.
Антитела, которые связывают большой
Рисунок 9.1 Большой Т-антиген в клетках, трансформированных SV40.
Антитела, которые связывают большой
Слайд 3Рисунок 9.3 Влияние p53 на трансформацию клеток
кДНК, кодирующая онкоген ras была
Рисунок 9.3 Влияние p53 на трансформацию клеток
кДНК, кодирующая онкоген ras была
Слайд 4Рис. 9.4. Частота мутантных аллелей p53 в геномах опухолевых клеток человека
Как
Рис. 9.4. Частота мутантных аллелей p53 в геномах опухолевых клеток человека
Как
Слайд 5Рисунок 9.5. Влияние мутантных аллелей p53 на зародышевую линию мыши.
Этот график
Рисунок 9.5. Влияние мутантных аллелей p53 на зародышевую линию мыши.
Этот график
Слайд 6Рисунок 9.6 Природа мутаций p53
(A) Как указано на этих круговых диаграммах,
Рисунок 9.6 Природа мутаций p53
(A) Как указано на этих круговых диаграммах,
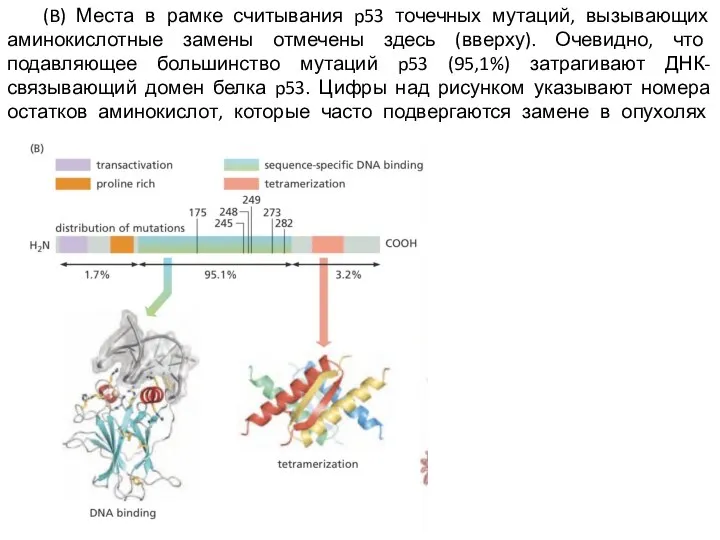
Слайд 7 (B) Места в рамке считывания p53 точечных мутаций, вызывающих аминокислотные замены отмечены
(B) Места в рамке считывания p53 точечных мутаций, вызывающих аминокислотные замены отмечены
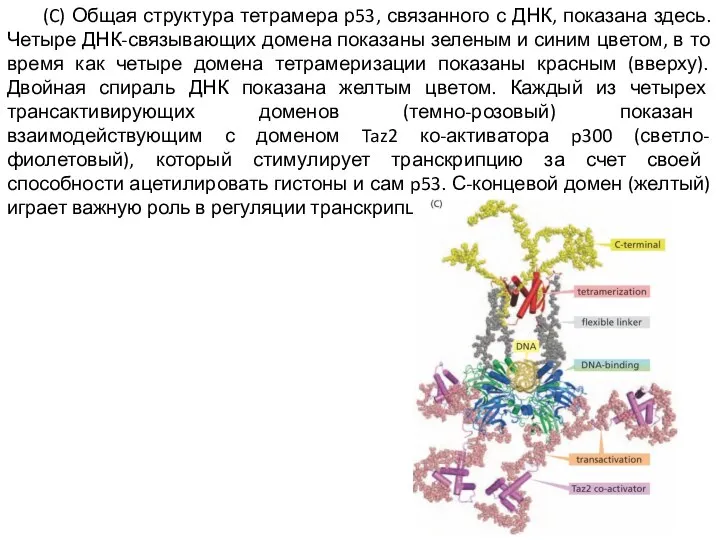
Слайд 8 (C) Общая структура тетрамера р53, связанного с ДНК, показана здесь. Четыре ДНК-связывающих
(C) Общая структура тетрамера р53, связанного с ДНК, показана здесь. Четыре ДНК-связывающих
Слайд 9Рисунок 9.7. Структура и функция p53
(A) В клетках, несущих единственный мутантный
Рисунок 9.7. Структура и функция p53
(A) В клетках, несущих единственный мутантный
Слайд 10 (B) Возможно, наиболее прямая демонстрация доминантно-негативного режима действия p53 произошла от «вбивания»
(B) Возможно, наиболее прямая демонстрация доминантно-негативного режима действия p53 произошла от «вбивания»
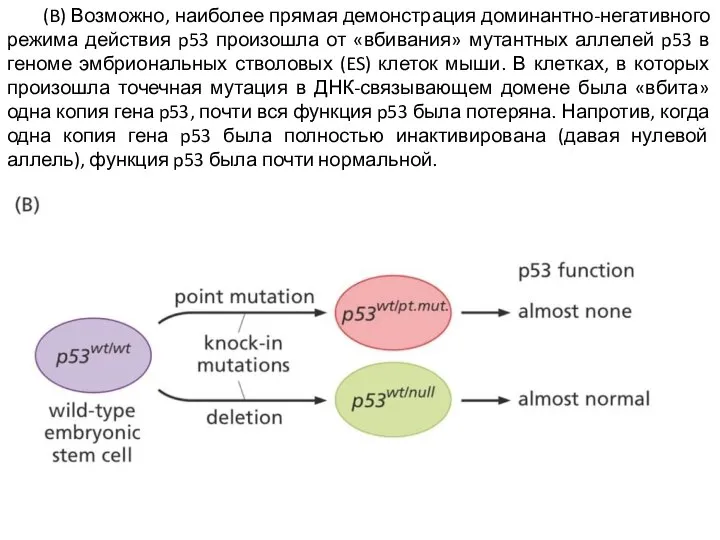
Слайд 11Рисунок 9.8. Сигналы, активирующие р53, и последующие эффекты р53.
Исследования функции p53
Рисунок 9.8. Сигналы, активирующие р53, и последующие эффекты р53.
Исследования функции p53
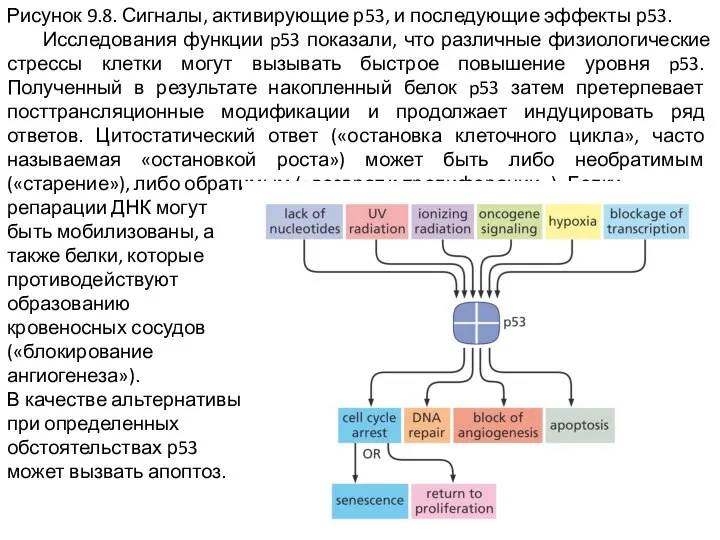
Слайд 12Рисунок 9.10. Контроль уровней p53 с помощью Mdm2.
После того, как концентрации
Рисунок 9.10. Контроль уровней p53 с помощью Mdm2.
После того, как концентрации
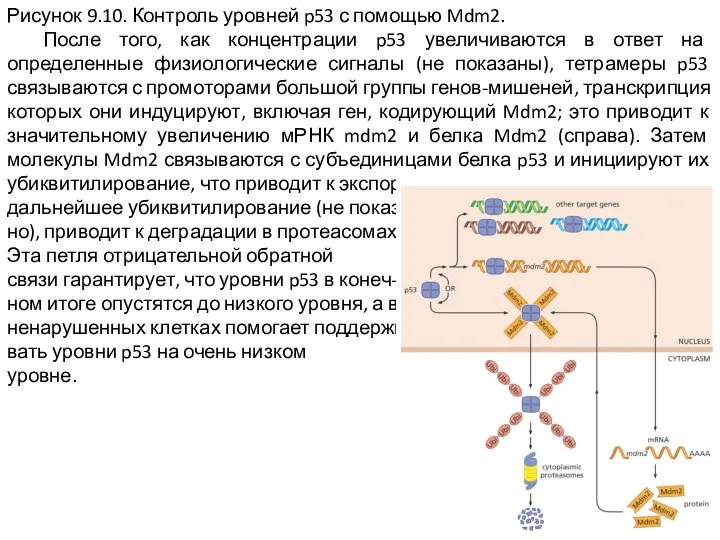
Слайд 13Рисунок 9.15. Контроль
апоптоза с помощью ARF
(A) Белок p14ARF чело-
века, называемый
Рисунок 9.15. Контроль
апоптоза с помощью ARF
(A) Белок p14ARF чело-
века, называемый
Слайд 14 (B) Различные онкогенные сигналы способствуют апоптозу благодаря их способности индуцировать активность E2F,
(B) Различные онкогенные сигналы способствуют апоптозу благодаря их способности индуцировать активность E2F,
Слайд 15 (C) Одна копия гена, кодирующего p19ARF, была инактивирована в зародышевой линии мыши
(C) Одна копия гена, кодирующего p19ARF, была инактивирована в зародышевой линии мыши
Слайд 16Рис. 9.21. Семейная предрасположенность к раку, обусловленная мутантными аллелями зародышевой линии р53.
Рис. 9.21. Семейная предрасположенность к раку, обусловленная мутантными аллелями зародышевой линии р53.
Слайд 17Рисунок 9.31 Структура и функции ингибитора апоптоза IAP (ингибиторы апоптоза) путем связывания
Рисунок 9.31 Структура и функции ингибитора апоптоза IAP (ингибиторы апоптоза) путем связывания
Слайд 18 (B) Молекула Smac/DIABLO предотвращает блокировку действия каспазы несколькими молекулами IAP, тем самым
(B) Молекула Smac/DIABLO предотвращает блокировку действия каспазы несколькими молекулами IAP, тем самым
 Комнатные растения
Комнатные растения Нервная система человека
Нервная система человека Инулин
Инулин Презентация на тему Лист. Внешнее и внутреннее строение листа
Презентация на тему Лист. Внешнее и внутреннее строение листа  Лишайники, их роль в природе и жизни человека
Лишайники, их роль в природе и жизни человека Экспрессия генов
Экспрессия генов Питомник Ясавэй представляет
Питомник Ясавэй представляет Нервная система (2)
Нервная система (2) Общая концепция механизации животноводства и роль специалистов в повышении эффективности ее функционирования. Лекция №1
Общая концепция механизации животноводства и роль специалистов в повышении эффективности ее функционирования. Лекция №1 Озеленение крыш с помощью современных материалов
Озеленение крыш с помощью современных материалов Как вырастить растение
Как вырастить растение Железо – элемент жизни
Железо – элемент жизни Двигательный анализатор человека. Развитие произвольных движений
Двигательный анализатор человека. Развитие произвольных движений Podstawy anatomiczne
Podstawy anatomiczne Генетики пола. Наследование признаков, сцепленных с полом
Генетики пола. Наследование признаков, сцепленных с полом Ярмарка знаний. Игра
Ярмарка знаний. Игра Ткани растений и животных
Ткани растений и животных Зимующие птицы
Зимующие птицы Радиационная генетика
Радиационная генетика Направления в эволюции. (9 класс)
Направления в эволюции. (9 класс) EVGENIKA_kg (2)
EVGENIKA_kg (2) Значение насекомых в природе
Значение насекомых в природе Котики-антибиотики. Ознакомление с базовыми методами создания генно-модифицированных организмов
Котики-антибиотики. Ознакомление с базовыми методами создания генно-модифицированных организмов Развитие насекомых
Развитие насекомых Органы чувств
Органы чувств Понятие о ДНК как хранителе и носителе генетического материала. 7 класс
Понятие о ДНК как хранителе и носителе генетического материала. 7 класс Многообразие грибов
Многообразие грибов Костные рыбы (Osteichthyes)
Костные рыбы (Osteichthyes)