- Биосинтез нуклеиновых кислот (анаболизм). Репликация

Содержание
- 2. 1. Суть, значение 2. Исходный материал, матричная схема. Фермент (ферменты). 3. Механизм. Направление синтеза дочерней цепи.
- 3. Репликация ДНК. РЕПЛИКАЦИЯ (от позднелат. replicatio - повторение) (редупликация), самовоспроизведение ДНК обеспечивающее точное копирование генетической информации
- 4. Гипотеза о механизме репликации сформулирована в 1953 Дж. Уотсоном и Ф. Криком, которые предположили, что две
- 6. Почему исходными веществами являются дНТФ?? При отщеплении дифосфата при построении новой цепи каждый раз выделяется энергия,
- 7. В процессе репликации двойная спираль ДНК, состоящая из двух комплементарных полинуклеотидных цепей, раскручивается на отдельные цепи
- 9. Репликация, как и любой процесс матричного синтеза, состоит из 3 этапов:: Инициация (зарождение цепи) 2. Элонгация
- 10. Рассмотрим процесс репликации в прокариотической клетке, состоящий из нескольких стадий. Инициация – создание репликативной вилки. На
- 11. Элонгация Дочерние нити ДНК образуются на обеих нитях материнской ДНК. Этот процесс катализирует несколько ДНК-полимераз, которые
- 13. Итак, репликационная вилка асимметрична. Из двух синтезируемых дочерних цепей ДНК одна строится непрерывно, а другая -
- 14. РНК-праймеры затем наращиваются дезоксину-клеотидами с 3'-конца ДНК-полимеразой -III, которая про-должает наращивание до тех пор, пока строящаяся
- 15. Элонгация А - ведущая цепь; Б - отстающая цепь; В - фрагмент Оказаки.
- 16. Для образования непрерывной цепи ДНК из многих таких фрагментов, в действие вступает особая система репа-рации ДНК,
- 17. Терминации нет фактически, так как она происходит при окончании молекулы матричной ДНК. Скорость построения цепи у
- 19. Скачать презентацию
Слайд 21. Суть, значение
2. Исходный материал, матричная схема. Фермент (ферменты).
3. Механизм. Направление синтеза
1. Суть, значение
2. Исходный материал, матричная схема. Фермент (ферменты).
3. Механизм. Направление синтеза

Слайд 3 Репликация ДНК.
РЕПЛИКАЦИЯ (от позднелат. replicatio - повторение) (редупликация), самовоспроизведение ДНК обеспечивающее
Репликация ДНК.
РЕПЛИКАЦИЯ (от позднелат. replicatio - повторение) (редупликация), самовоспроизведение ДНК обеспечивающее

Слайд 4 Гипотеза о механизме репликации сформулирована в 1953 Дж. Уотсоном и Ф. Криком,
Гипотеза о механизме репликации сформулирована в 1953 Дж. Уотсоном и Ф. Криком,

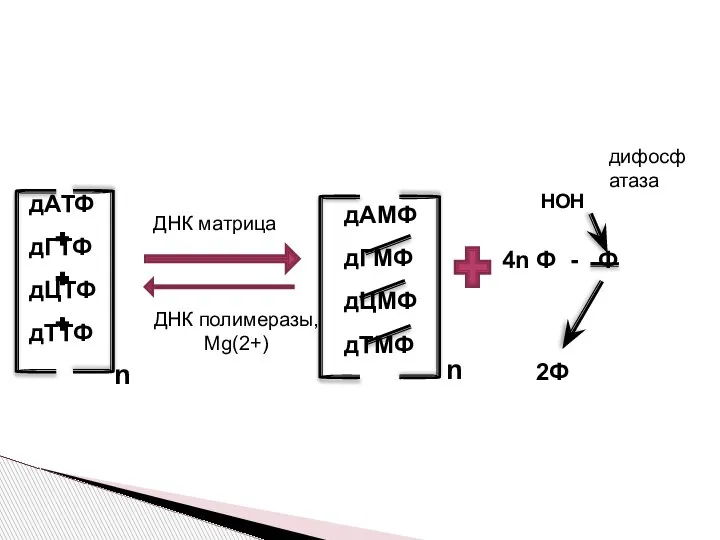
Слайд 6Почему исходными веществами являются дНТФ??
При отщеплении дифосфата при построении новой цепи
Почему исходными веществами являются дНТФ??
При отщеплении дифосфата при построении новой цепи

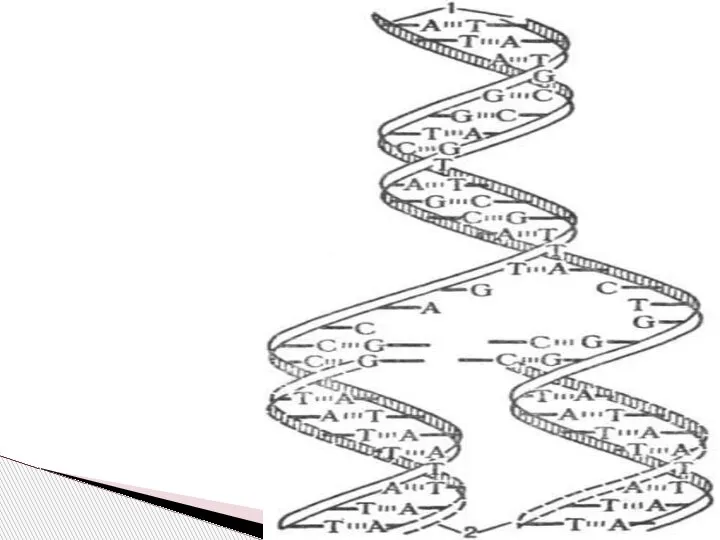
Слайд 7 В процессе репликации двойная спираль ДНК, состоящая из двух комплементарных полинуклеотидных
В процессе репликации двойная спираль ДНК, состоящая из двух комплементарных полинуклеотидных

Слайд 9 Репликация, как и любой процесс матричного синтеза, состоит из 3 этапов::
Инициация (зарождение
Репликация, как и любой процесс матричного синтеза, состоит из 3 этапов::
Инициация (зарождение

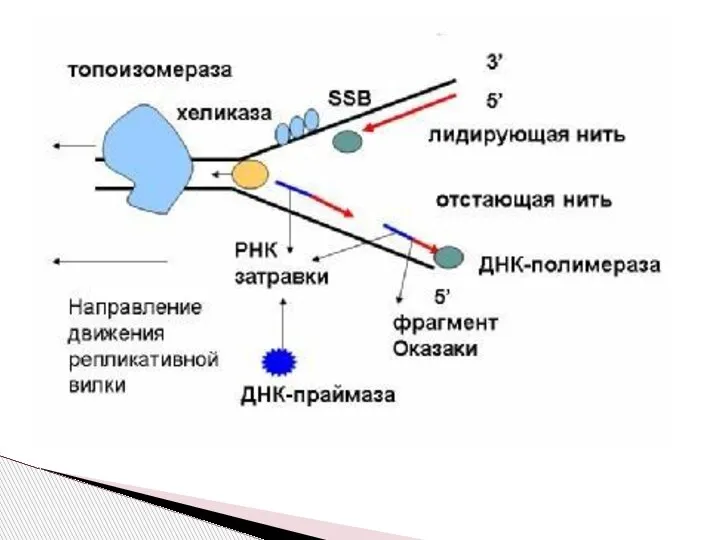
Слайд 10Рассмотрим процесс репликации в прокариотической клетке, состоящий из нескольких стадий.
Инициация – создание
Рассмотрим процесс репликации в прокариотической клетке, состоящий из нескольких стадий.
Инициация – создание

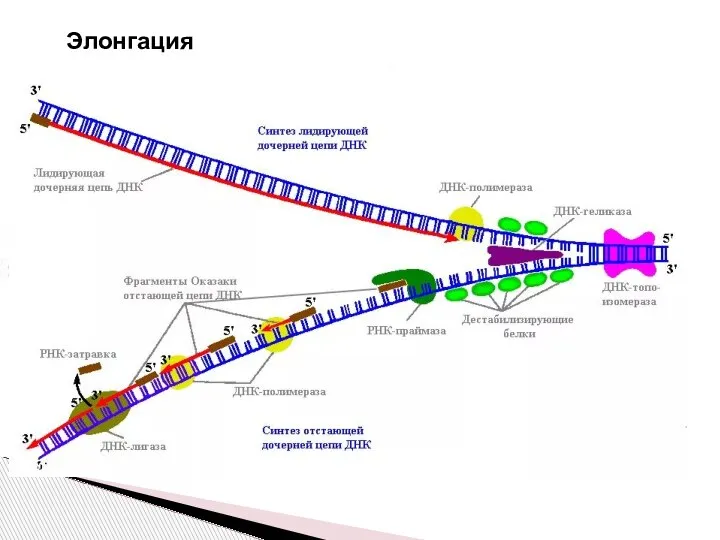
Слайд 11 Элонгация
Дочерние нити ДНК образуются на обеих нитях материнской ДНК. Этот процесс
Элонгация
Дочерние нити ДНК образуются на обеих нитях материнской ДНК. Этот процесс

Слайд 13 Итак, репликационная вилка асимметрична. Из двух синтезируемых дочерних цепей ДНК одна строится
Итак, репликационная вилка асимметрична. Из двух синтезируемых дочерних цепей ДНК одна строится

Слайд 14 РНК-праймеры затем наращиваются дезоксину-клеотидами с 3'-конца ДНК-полимеразой -III, которая про-должает наращивание до
РНК-праймеры затем наращиваются дезоксину-клеотидами с 3'-конца ДНК-полимеразой -III, которая про-должает наращивание до

Слайд 15Элонгация
А - ведущая цепь;
Б - отстающая цепь;
В - фрагмент Оказаки.
Элонгация
А - ведущая цепь;
Б - отстающая цепь;
В - фрагмент Оказаки.
Слайд 16 Для образования непрерывной цепи ДНК из многих таких фрагментов, в действие вступает
Для образования непрерывной цепи ДНК из многих таких фрагментов, в действие вступает

Слайд 17Терминации нет фактически, так как она происходит при окончании молекулы матричной ДНК.
Терминации нет фактически, так как она происходит при окончании молекулы матричной ДНК.

 Зудневые клещи
Зудневые клещи Композиция из комнатных растений Цветочная феерия
Композиция из комнатных растений Цветочная феерия Ферменты, или энзимы. Функции ферментов
Ферменты, или энзимы. Функции ферментов Состав и краткая характеристика основных компонентов пищи
Состав и краткая характеристика основных компонентов пищи Механизм дыхания
Механизм дыхания Домашние животные
Домашние животные Дарвин и теория эволюции
Дарвин и теория эволюции Augi vasarā
Augi vasarā Учение И.П. Павлова о высшей нервной деятельности и ее типологических особенностях
Учение И.П. Павлова о высшей нервной деятельности и ее типологических особенностях Вплив регулятору росту рослин АКМ на формування продуктивності рослин сафлору красильного сорту Живчик
Вплив регулятору росту рослин АКМ на формування продуктивності рослин сафлору красильного сорту Живчик Муравьиная ферма
Муравьиная ферма Агробіологічна характеристика однорічних ранніх ярих бур’янів (гостриця лежача, камеліна звичайна, біформа промениста)
Агробіологічна характеристика однорічних ранніх ярих бур’янів (гостриця лежача, камеліна звичайна, біформа промениста) Презентация на тему Сравни позвоночных
Презентация на тему Сравни позвоночных  En el bosque
En el bosque Физиология растений
Физиология растений ВПР биология 11 класс. Болезни
ВПР биология 11 класс. Болезни 20140106_zhivotnyy_mir_ro
20140106_zhivotnyy_mir_ro Iýmit siňdiris sistemasy
Iýmit siňdiris sistemasy Жизненный цикл клетки. Митоз
Жизненный цикл клетки. Митоз Найди рыбу. Игра
Найди рыбу. Игра Формирование метапредметных результатов школьников при реализации ФГОС ООО по биологии
Формирование метапредметных результатов школьников при реализации ФГОС ООО по биологии Нервная система
Нервная система Развитие жизни на Земле
Развитие жизни на Земле Лисица
Лисица Кто такие земноводные?
Кто такие земноводные? Гормоны
Гормоны lektsia_1
lektsia_1 Пути обмена отдельных аминокислот
Пути обмена отдельных аминокислот