- ДНК секвенирлеу әдістері

Содержание
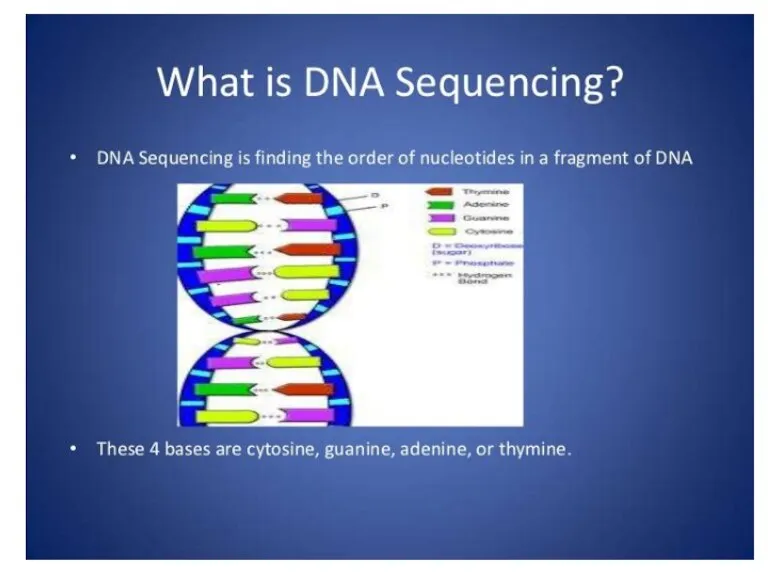
- 2. Биополимерлердің бірінші реттілігін анықтайтын әдісте секвенирлеу деген арнайы термин қолданылады. Белгілі ДНК молекуласындағы нуклеотидтердің санын мен
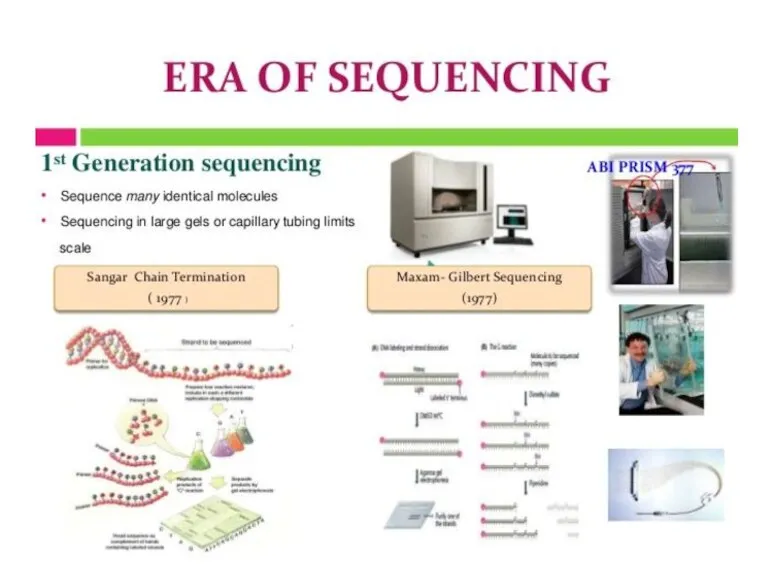
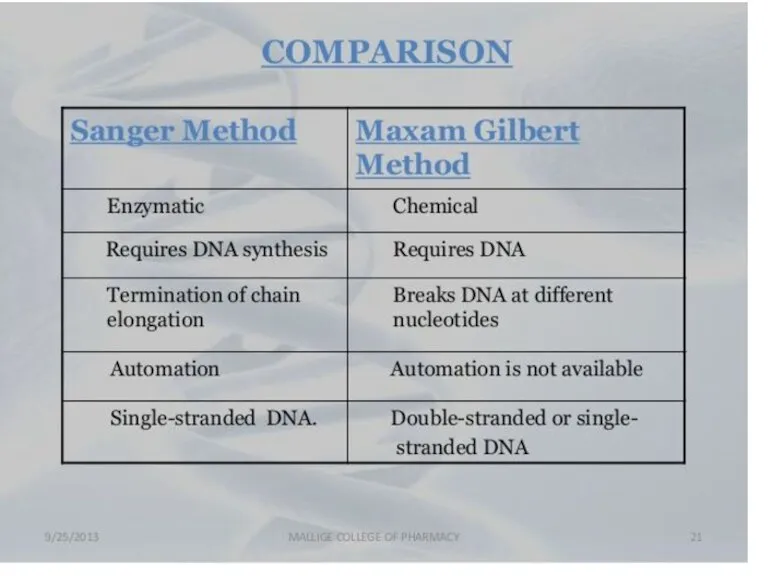
- 10. Сэнгер әдісі: «терминатор» әдісі. 4 кезең: Денатурация Синтез Терминация Электрофорез
- 11. Сэнгер әдісі: «терминатор» әдісі.
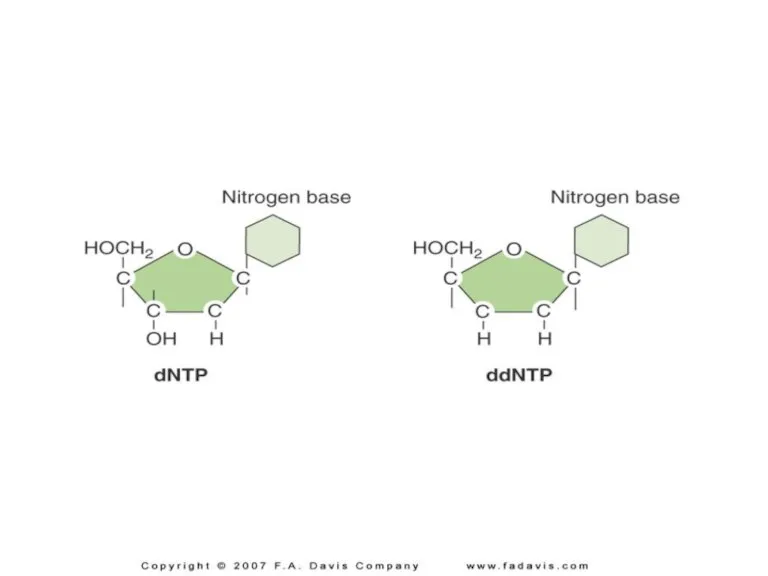
- 15. O H H O = P – O O O 2’, 3’ dideoxy nucleotide Can not
- 16. Сэнгер әдісі Белгіленген праймер мен бастапқы матрицаны қосу. ddNTP-ді 4 пробирканың біріне қосуы.
- 17. A C G T Сэнгер әдісі
- 20. Automated Version of the Dideoxy Method
- 22. Overview of Sanger sequencing method
- 23. ДНК + полимераза + праймер + dCTP dTTP dGTP dATP ddATP ddGTP ddTTP ddCTP полимеразная реакция
- 27. Метод Максама-Гилберта В основе метода секвенирования ДНК путем химической деградации лежит ограниченное расщепление меченого фрагмента ДНК
- 30. Стадии Двухцепочечный фрагмент, подлежащий секвенированию, изолирован и радиоактивно помечен на 5’-концах 32P. 2. Затем фрагмент разрезают
- 34. 9. The labeled fragments in the gel are visualized by autoradiography. 10. The sequence is read
- 35. Химическая модификация азотистых основание заключается в: Депуринизации (A+G) с использованием муравьиной кислоты Метилировании гуанина с использованием
- 37. (Next generation sequencing), NGS
- 41. Общая схема работы NGS: от исходной ДНК до «букв» на экране приготовление библиотеки (дробление ДНК и
- 46. 454 – принцип метода ДНК фрагментируют и лигируют к фрагментам адаптеры Фрагмент закрепляется на микро-шарике, покрытой
- 47. 454 – принцип метода Носитель – плашка с множеством (> 1 000 000) лунок В лунки
- 48. 454 – принцип метода Через лунки в пропускают заданном реагенты порядке (dNTP, люциферин, аденозинфосфосульфат) При присоединении
- 49. Секвенирование Illumina - принцип метода 1. ДНК фрагментируют и лигируют к фрагментам адаптеры 2. ДНК ячейки,
- 50. Секвенирование Illumina - принцип метода 7. Через ячейку пропускают реагенты (флуоресцентно меченые терминированные dNTP и полимеразу)
- 51. Преимущества: •высокая точность •универсальность •доступность ПО для обработки и анализа результатов •наименьшая цена получаемых данных (в
- 52. Секвенирование путем лигирования (SOLiD) Преимущества: высокая точность возможность использовать часть дорожек на ячейке Недостатки очень короткие
- 55. Полупроводниковое секвенирование Самая новая из технологий cеквенирования 2 поколения Сходно с 454-секвенированием, но регистрируется не свет,
- 56. Бисульфитті секвенирлеу Бисульфитті секвенирлеу - бисульфитпен өңдеу арқылы ДНҚ метилдеу паттернасын зерттеуге бағытталған әдістер тобының жалпы
- 57. Бисульфит цитозинді урацилға айналдыра отырып, бір тізбекті ДНҚ-ға әсер етеді. Егер осы цитозин метилденген болса, яғни
- 59. Бисульфитті секвенирлеудің бірінші әдісі 1992 жылы сипатталған. Метилдеу паттернін анықтау үшін ПЦР қолданылды, ол үшін праймерлер
- 62. Бисульфитті секвенирлеу әдістері бірқатар шектеулерге ие. Сүтқоректілерде ДНҚ 5-гидроксиметилцитозин модификациясы кең таралған. Бисульфитпен өңдеу кезінде 5-гидроксиметилцитозин
- 67. Скачать презентацию
Слайд 2Биополимерлердің бірінші реттілігін анықтайтын әдісте секвенирлеу деген арнайы термин қолданылады.
Белгілі ДНК молекуласындағы нуклеотидтердің санын
Биополимерлердің бірінші реттілігін анықтайтын әдісте секвенирлеу деген арнайы термин қолданылады.
Белгілі ДНК молекуласындағы нуклеотидтердің санын


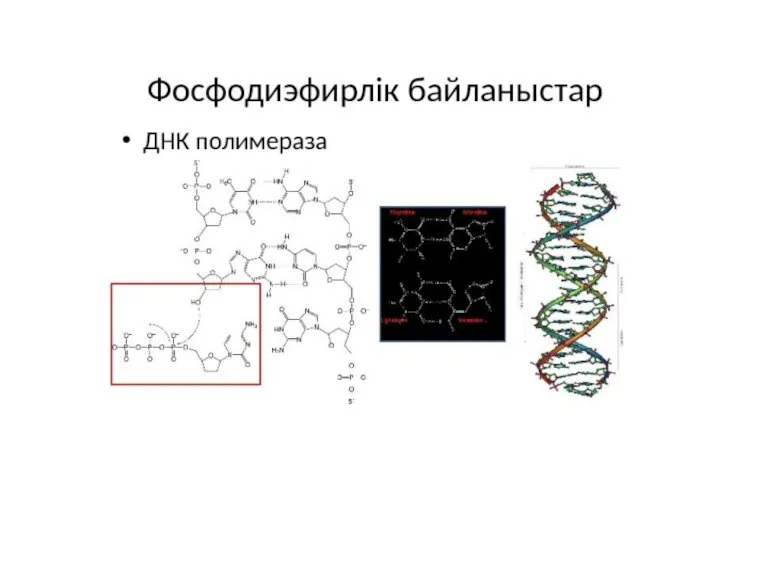



Слайд 10Сэнгер әдісі: «терминатор» әдісі.
4 кезең:
Денатурация
Синтез
Терминация
Электрофорез
Сэнгер әдісі: «терминатор» әдісі.
4 кезең:
Денатурация
Синтез
Терминация
Электрофорез

Слайд 11Сэнгер әдісі: «терминатор» әдісі.
Сэнгер әдісі: «терминатор» әдісі.

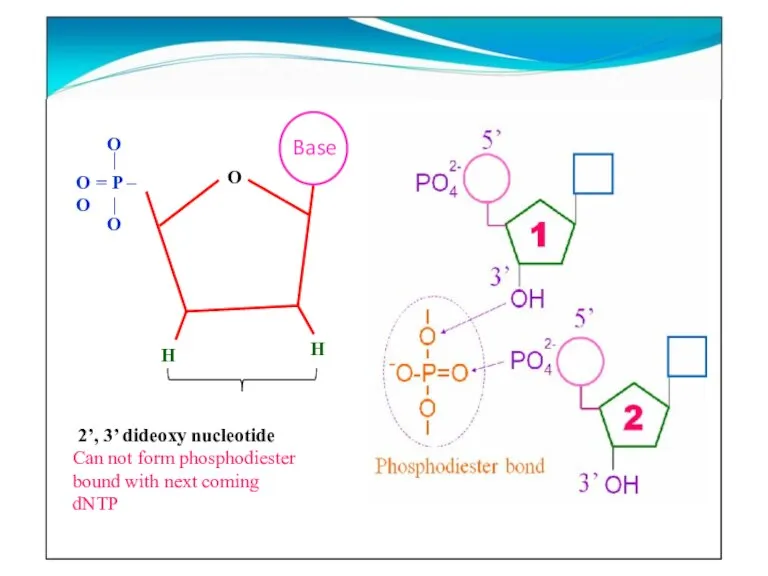
Слайд 15O
H
H
O = P – O
O
O
2’, 3’ dideoxy nucleotide Can not form phosphodiester
O
H
H
O = P – O
O
O
2’, 3’ dideoxy nucleotide Can not form phosphodiester
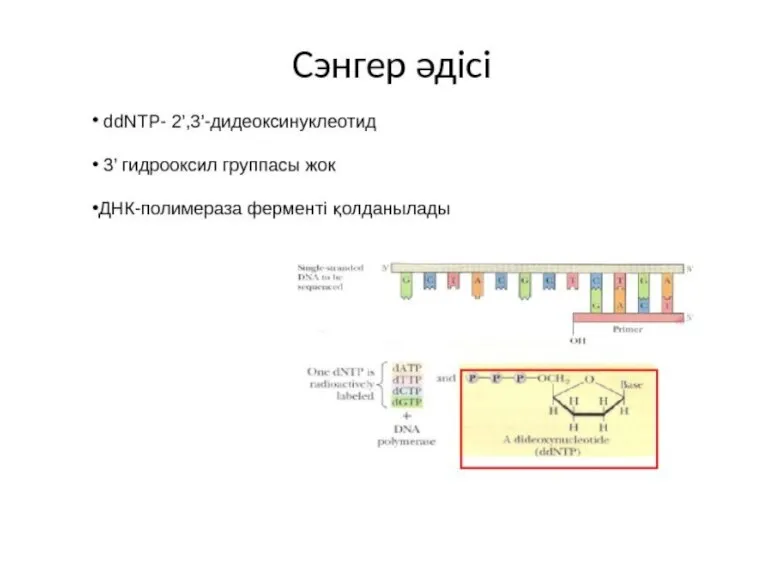
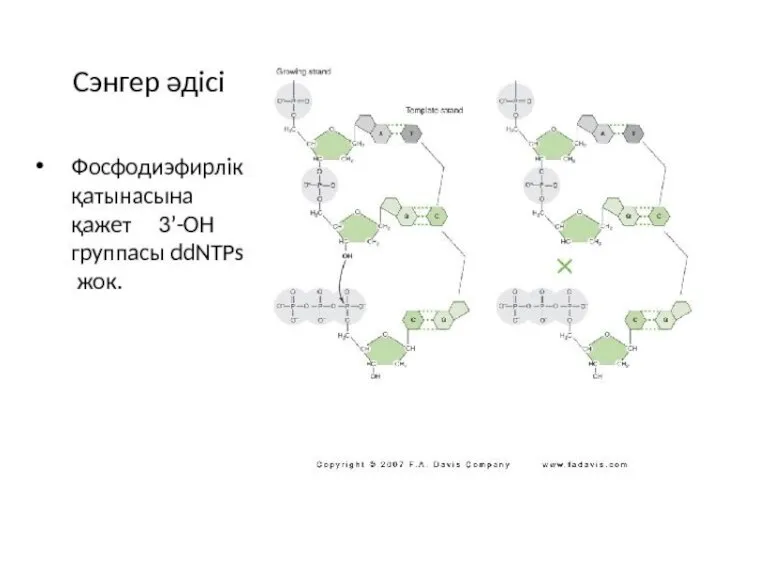
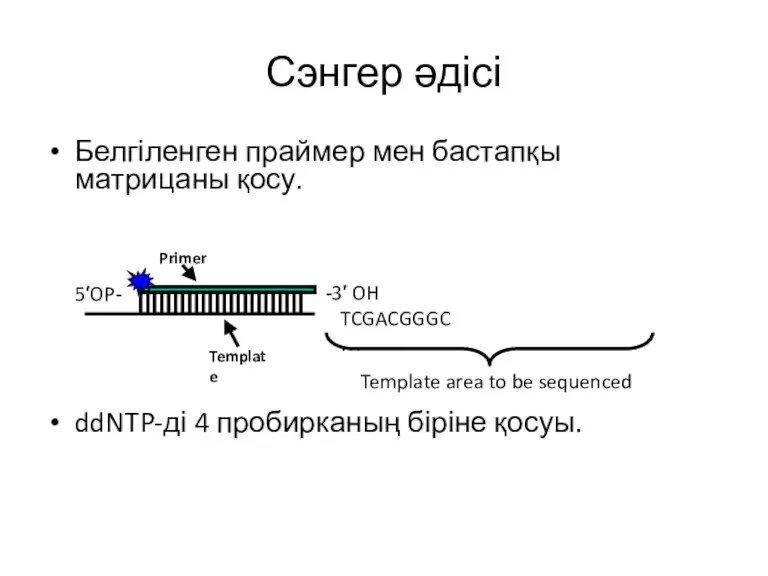
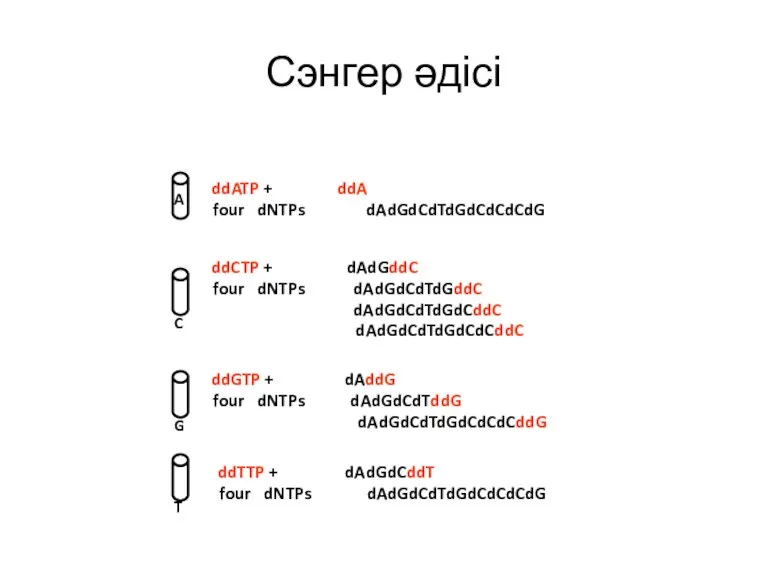
Слайд 16Сэнгер әдісі
Белгіленген праймер мен бастапқы матрицаны қосу.
ddNTP-ді 4 пробирканың біріне қосуы.
Сэнгер әдісі
Белгіленген праймер мен бастапқы матрицаны қосу.
ddNTP-ді 4 пробирканың біріне қосуы.
Слайд 17A
C
G
T
Сэнгер әдісі
A
C
G
T
Сэнгер әдісі
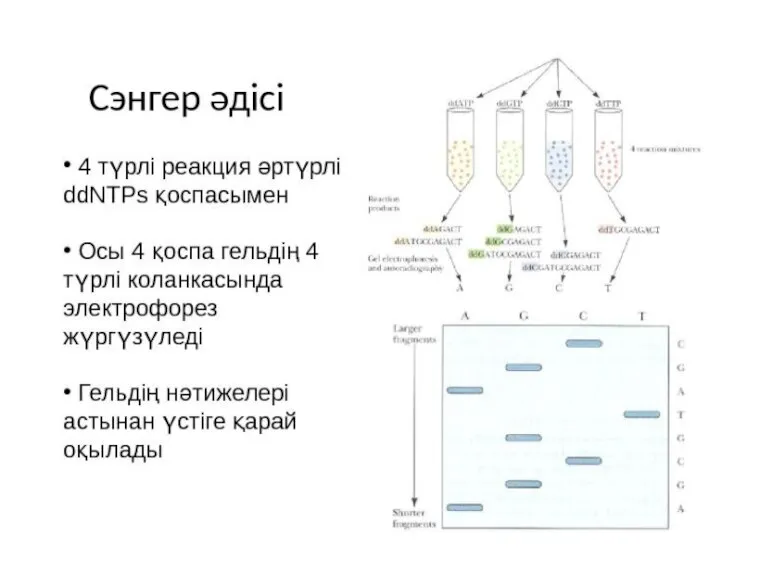

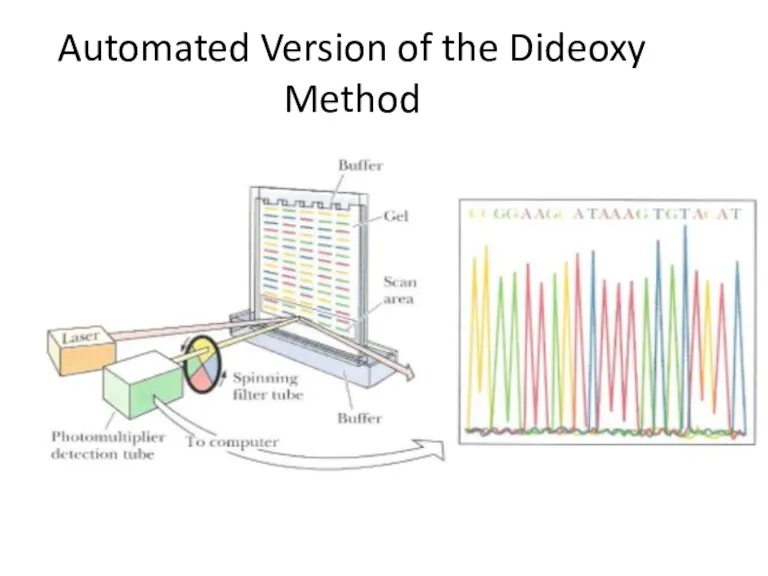
Слайд 20Automated Version of the Dideoxy Method
Automated Version of the Dideoxy Method
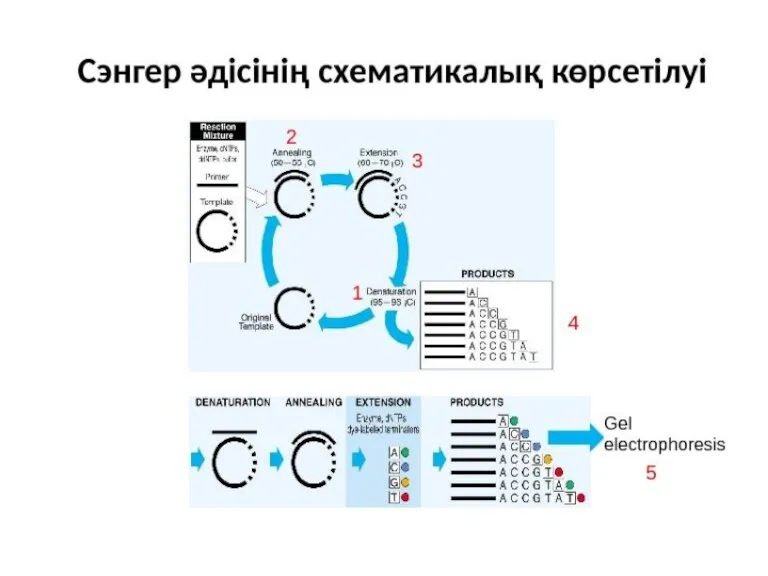
Слайд 22Overview of Sanger sequencing method
Overview of Sanger sequencing method
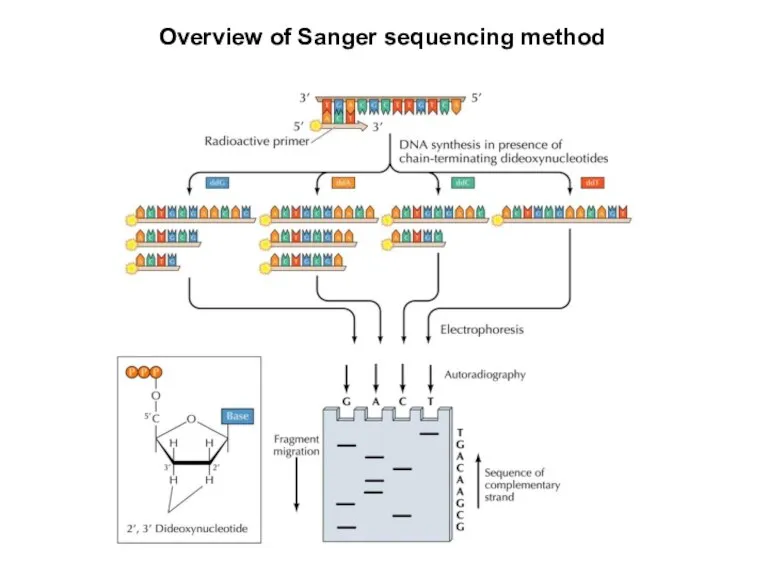
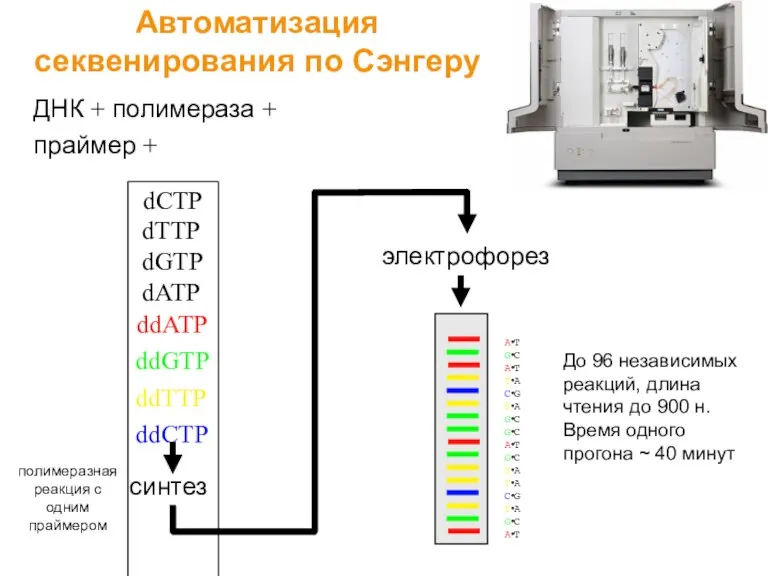
Слайд 23ДНК + полимераза +
праймер +
dCTP dTTP dGTP dATP
ddATP ddGTP ddTTP ddCTP
полимеразная реакция
ДНК + полимераза +
праймер +
dCTP dTTP dGTP dATP
ddATP ddGTP ddTTP ddCTP
полимеразная реакция



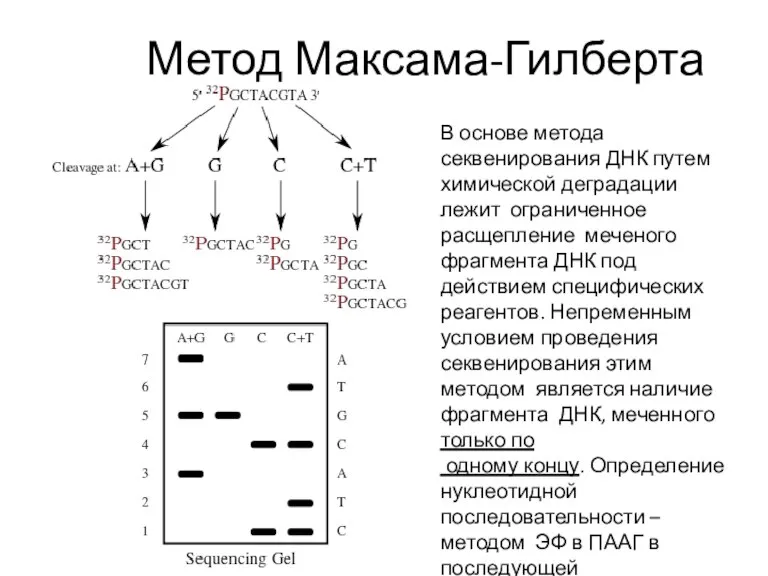
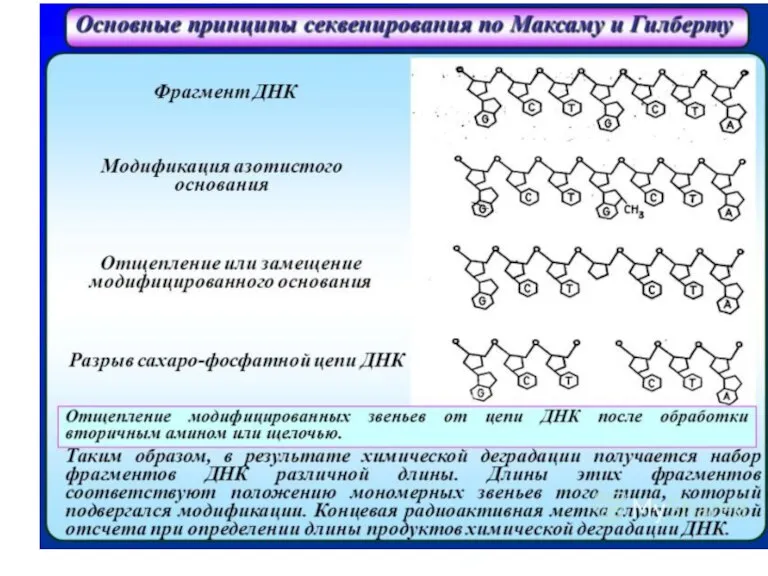
Слайд 27Метод Максама-Гилберта
В основе метода секвенирования ДНК путем химической деградации лежит ограниченное расщепление
Метод Максама-Гилберта
В основе метода секвенирования ДНК путем химической деградации лежит ограниченное расщепление

Слайд 30
Стадии
Двухцепочечный фрагмент, подлежащий секвенированию, изолирован и радиоактивно помечен на 5’-концах 32P.
2. Затем
Стадии
Двухцепочечный фрагмент, подлежащий секвенированию, изолирован и радиоактивно помечен на 5’-концах 32P.
2. Затем


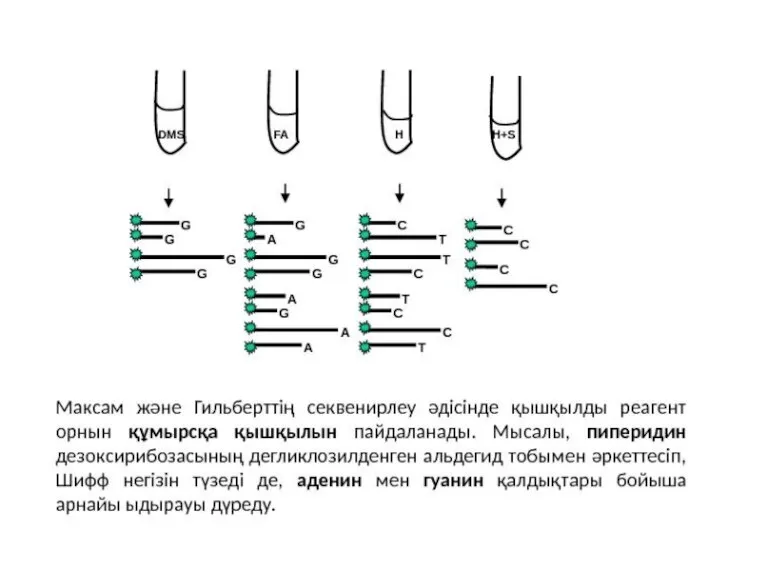
Слайд 34
9. The labeled fragments
in the gel are visualized
by
9. The labeled fragments
in the gel are visualized
by

Слайд 35Химическая модификация азотистых основание заключается в:
Депуринизации (A+G) с использованием муравьиной кислоты
Метилировании гуанина
Химическая модификация азотистых основание заключается в:
Депуринизации (A+G) с использованием муравьиной кислоты
Метилировании гуанина

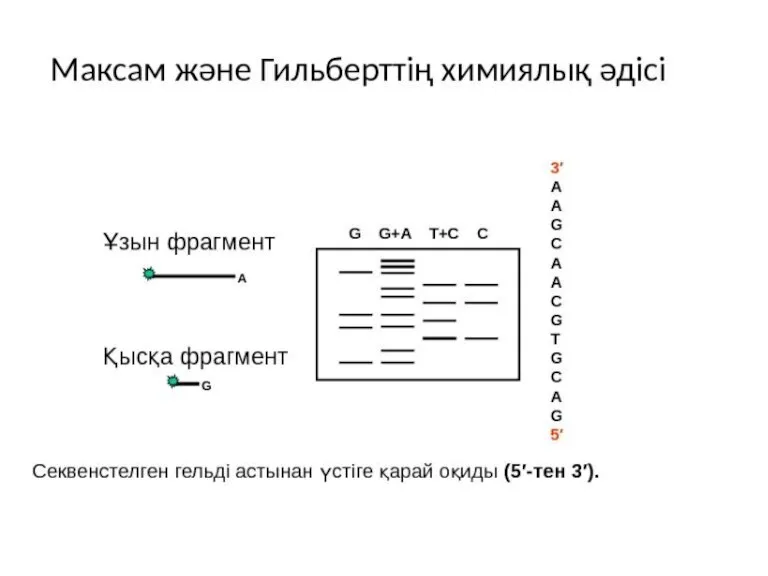
Слайд 37(Next generation sequencing),
NGS
(Next generation sequencing),
NGS




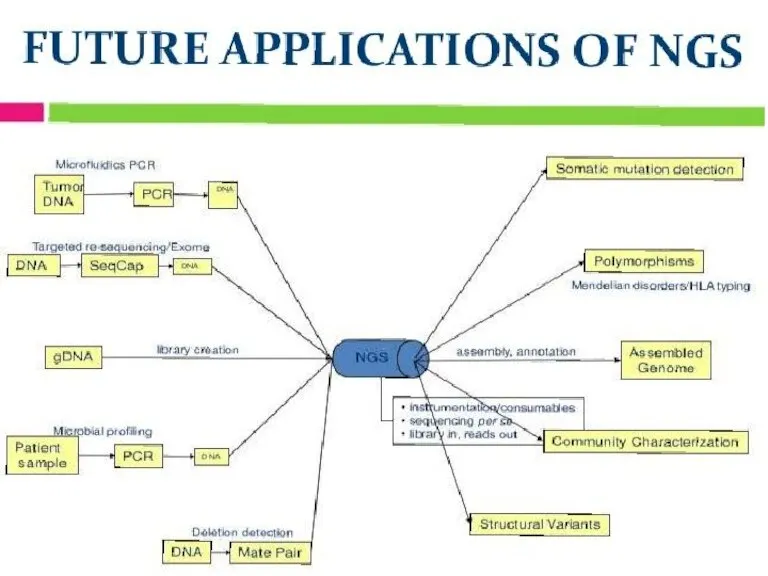
Слайд 41Общая схема работы NGS: от исходной ДНК до «букв» на экране
приготовление библиотеки
Общая схема работы NGS: от исходной ДНК до «букв» на экране
приготовление библиотеки
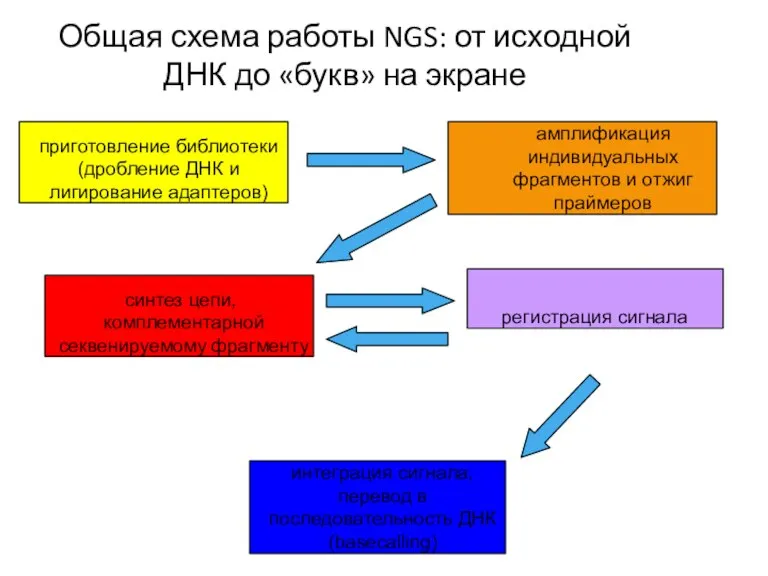



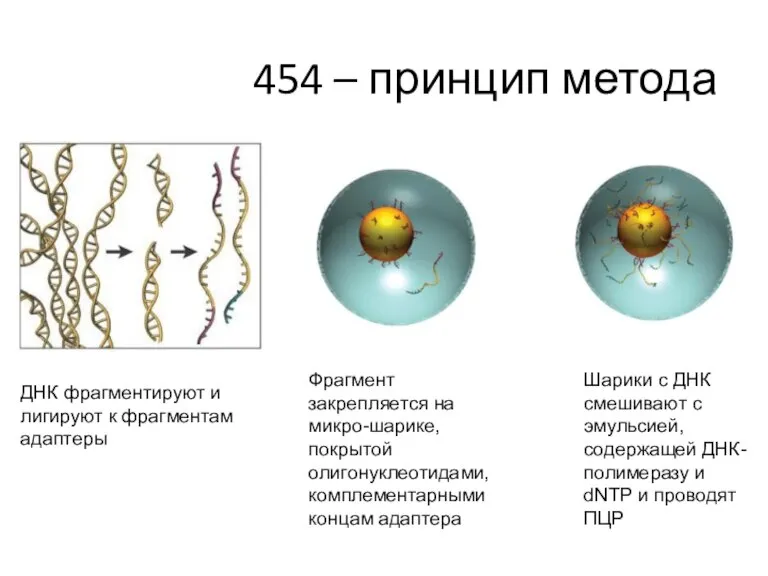
Слайд 46454 – принцип метода
ДНК фрагментируют и лигируют к фрагментам адаптеры
Фрагмент закрепляется на
454 – принцип метода
ДНК фрагментируют и лигируют к фрагментам адаптеры
Фрагмент закрепляется на
Слайд 47454 – принцип метода
Носитель – плашка с множеством (> 1 000
000) лунок
В
454 – принцип метода
Носитель – плашка с множеством (> 1 000
000) лунок
В

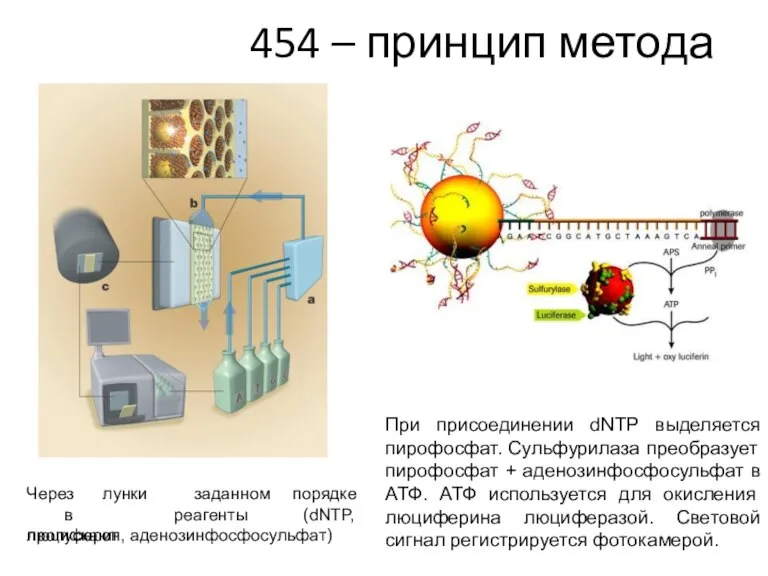
Слайд 48454 – принцип метода
Через лунки в пропускают
заданном реагенты
порядке (dNTP,
люциферин, аденозинфосфосульфат)
При присоединении dNTP выделяется пирофосфат.
454 – принцип метода
Через лунки в пропускают
заданном реагенты
порядке (dNTP,
люциферин, аденозинфосфосульфат)
При присоединении dNTP выделяется пирофосфат.
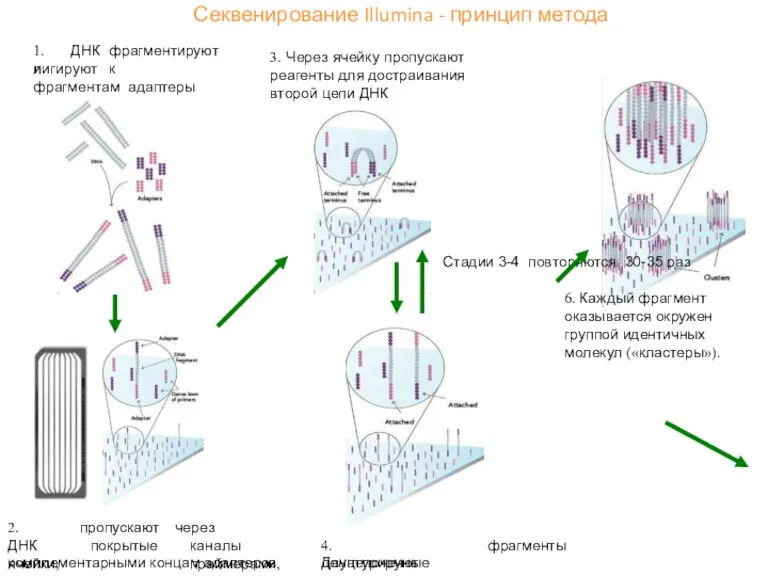
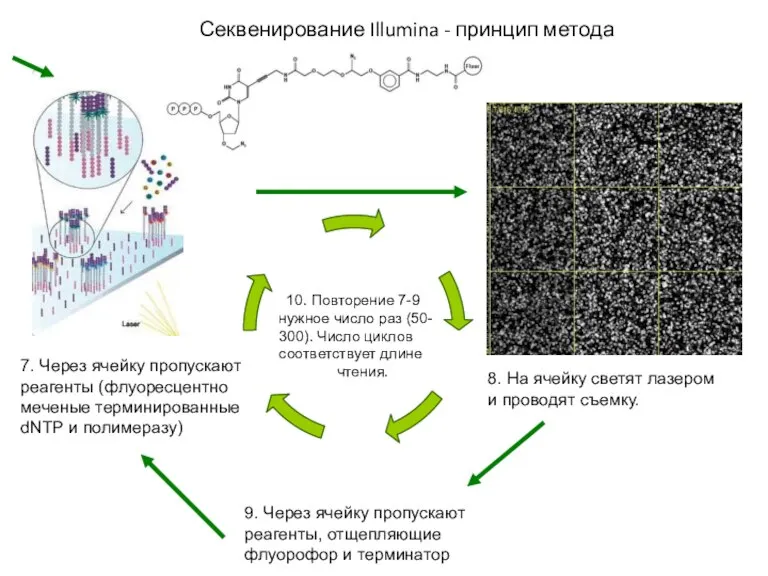
Слайд 49Секвенирование Illumina - принцип метода
1. ДНК фрагментируют и
лигируют к фрагментам адаптеры
2. ДНК ячейки,
пропускают покрытые
через каналы праймерами,
комплементарными концам адаптеров
3.
Секвенирование Illumina - принцип метода
1. ДНК фрагментируют и
лигируют к фрагментам адаптеры
2. ДНК ячейки,
пропускают покрытые
через каналы праймерами,
комплементарными концам адаптеров
3.
Слайд 50Секвенирование Illumina - принцип метода
7. Через ячейку пропускают реагенты (флуоресцентно меченые терминированные
Секвенирование Illumina - принцип метода
7. Через ячейку пропускают реагенты (флуоресцентно меченые терминированные
Слайд 51Преимущества:
•высокая точность
•универсальность
•доступность ПО для обработки и анализа результатов
•наименьшая цена получаемых данных (в
Преимущества:
•высокая точность
•универсальность
•доступность ПО для обработки и анализа результатов
•наименьшая цена получаемых данных (в

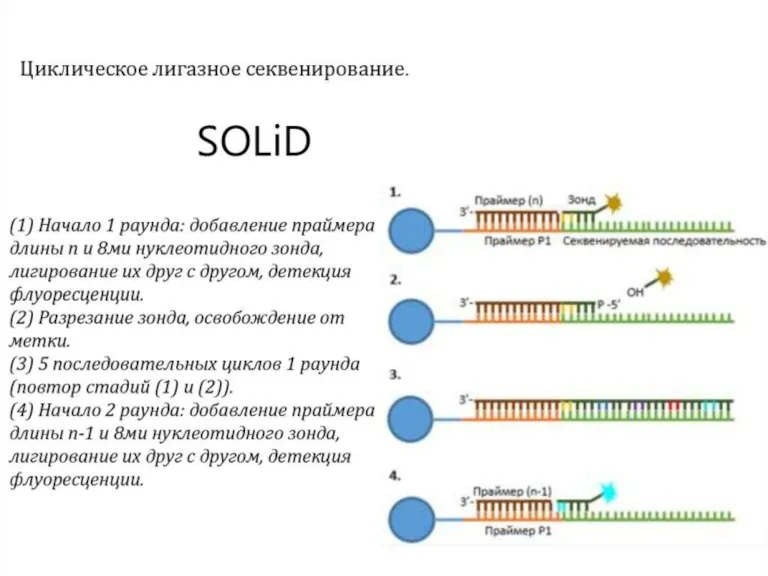
Слайд 52Секвенирование путем лигирования (SOLiD)
Преимущества:
высокая точность
возможность использовать часть дорожек на ячейке Недостатки
очень короткие
Секвенирование путем лигирования (SOLiD)
Преимущества:
высокая точность
возможность использовать часть дорожек на ячейке Недостатки
очень короткие


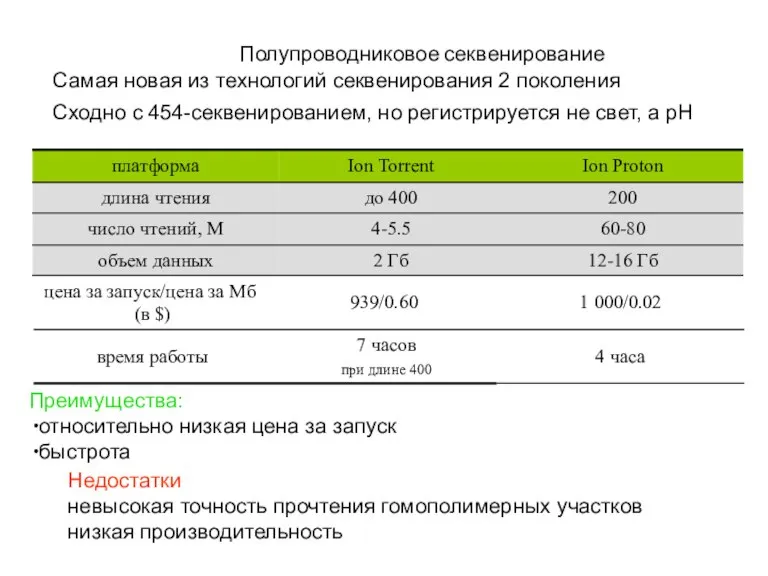
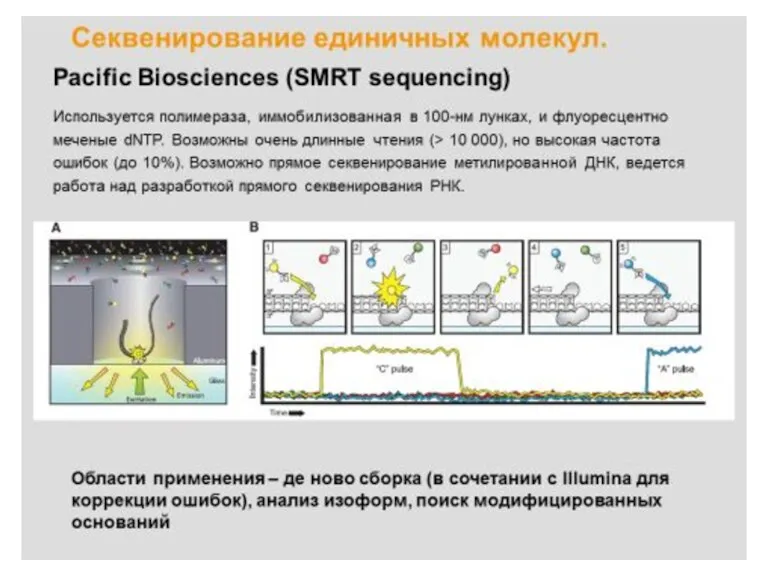
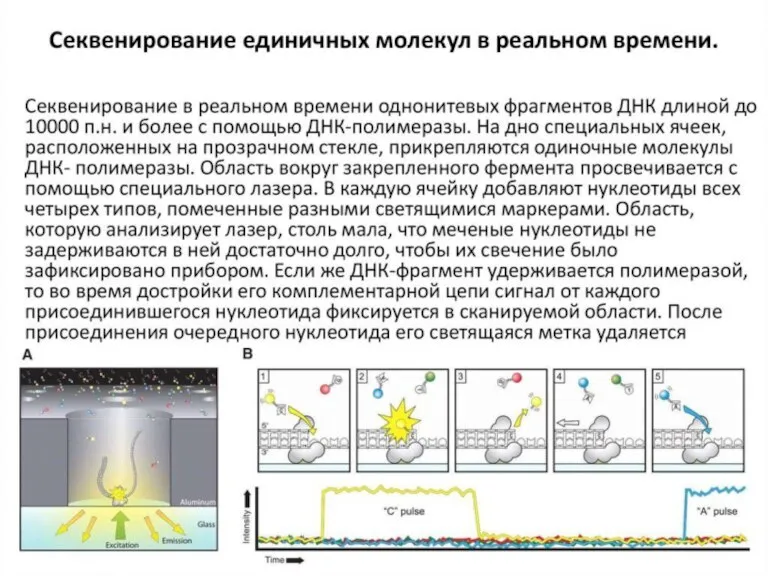
Слайд 55Полупроводниковое секвенирование
Самая новая из технологий cеквенирования 2 поколения Сходно с 454-секвенированием, но
Полупроводниковое секвенирование
Самая новая из технологий cеквенирования 2 поколения Сходно с 454-секвенированием, но
Слайд 56Бисульфитті секвенирлеу
Бисульфитті секвенирлеу - бисульфитпен өңдеу арқылы ДНҚ метилдеу паттернасын зерттеуге бағытталған
Бисульфитті секвенирлеу
Бисульфитті секвенирлеу - бисульфитпен өңдеу арқылы ДНҚ метилдеу паттернасын зерттеуге бағытталған

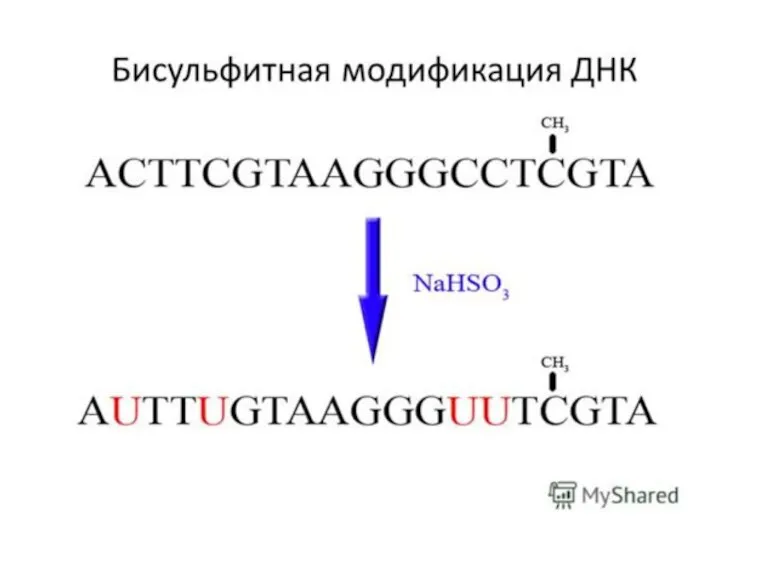
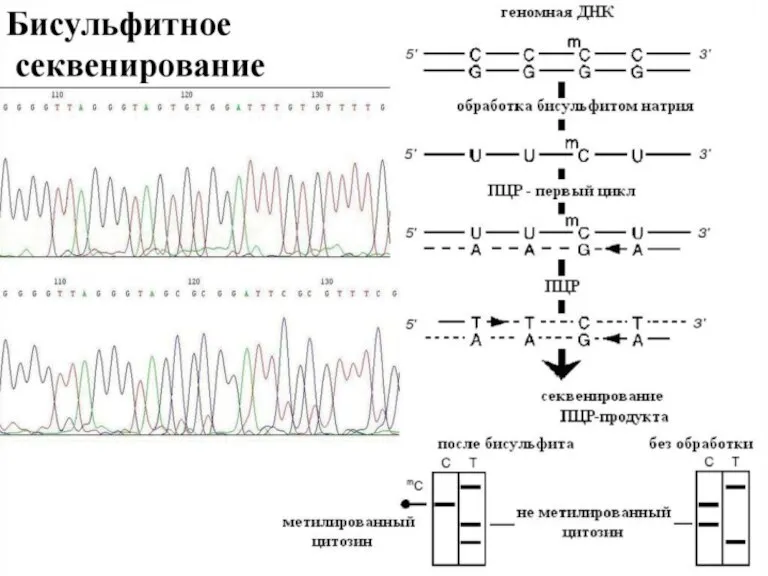
Слайд 57Бисульфит цитозинді урацилға айналдыра отырып, бір тізбекті ДНҚ-ға әсер етеді. Егер осы
Бисульфит цитозинді урацилға айналдыра отырып, бір тізбекті ДНҚ-ға әсер етеді. Егер осы

Слайд 59Бисульфитті секвенирлеудің бірінші әдісі 1992 жылы сипатталған. Метилдеу паттернін анықтау үшін ПЦР
Бисульфитті секвенирлеудің бірінші әдісі 1992 жылы сипатталған. Метилдеу паттернін анықтау үшін ПЦР

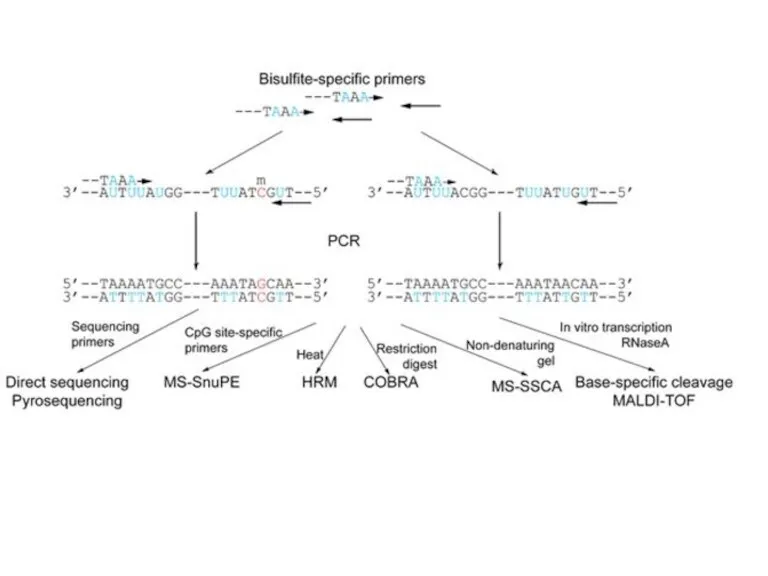
Слайд 62Бисульфитті секвенирлеу әдістері бірқатар шектеулерге ие. Сүтқоректілерде ДНҚ 5-гидроксиметилцитозин модификациясы кең таралған.
Бисульфитті секвенирлеу әдістері бірқатар шектеулерге ие. Сүтқоректілерде ДНҚ 5-гидроксиметилцитозин модификациясы кең таралған.

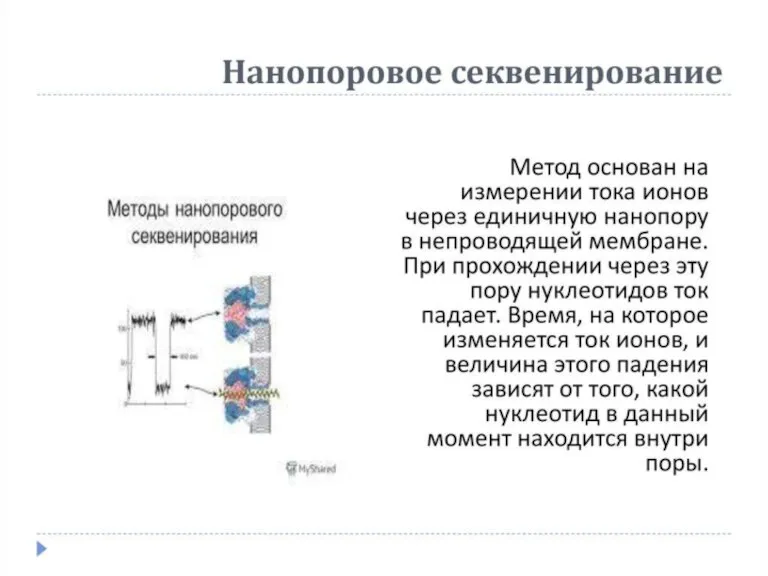
 Липиды
Липиды Физиология дыхания
Физиология дыхания Размножение организмов
Размножение организмов Видовой состав растений горы Магнитной
Видовой состав растений горы Магнитной Земноводные. Пресмыкающиеся
Земноводные. Пресмыкающиеся Theory of phyloembroyogenesis – deviation. Phylogenesis of the chord avian nervous system. Developmental disorders of
Theory of phyloembroyogenesis – deviation. Phylogenesis of the chord avian nervous system. Developmental disorders of Презентация на тему РАЗМНОЖЕНИЕ И ОПЛОДОТВОРЕНИЕ У РАСТЕНИЙ
Презентация на тему РАЗМНОЖЕНИЕ И ОПЛОДОТВОРЕНИЕ У РАСТЕНИЙ  Презентация на тему Кожно-мышечная чувствительность. Обоняние. Вкус
Презентация на тему Кожно-мышечная чувствительность. Обоняние. Вкус  Труд человека и бионика
Труд человека и бионика Презентация на тему История генетики
Презентация на тему История генетики  Виды рыб, часто использующихся в питании
Виды рыб, часто использующихся в питании В мире животных и растений
В мире животных и растений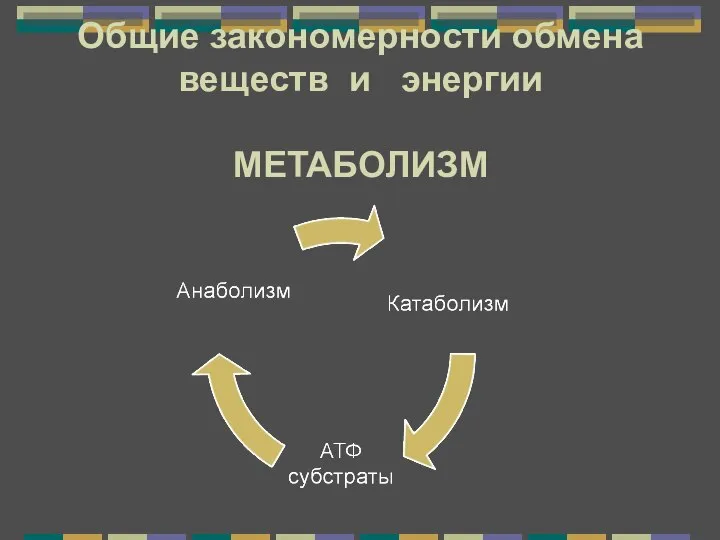 Метаболизм. Энергетический обмен
Метаболизм. Энергетический обмен Размножение организмов
Размножение организмов Основы учения о наследственности и изменчивости. Генетика
Основы учения о наследственности и изменчивости. Генетика Райские птицы. Биогеография и экология
Райские птицы. Биогеография и экология Деление клетки
Деление клетки Грибы. Тест
Грибы. Тест Зеленая аптека. Создание условий для ознакомления детей с миром целебных трав
Зеленая аптека. Создание условий для ознакомления детей с миром целебных трав Флора Ирана
Флора Ирана Многообразие организмов и их классификация
Многообразие организмов и их классификация Класс Пресмыкающиеся. Особенности внешнего и внутреннего строения. Многообразие. Происхождение
Класс Пресмыкающиеся. Особенности внешнего и внутреннего строения. Многообразие. Происхождение Деревья - очистители воздуха
Деревья - очистители воздуха Тест. Коллоквиум №3 (НМА, ЖВС, возбудимые ткани)
Тест. Коллоквиум №3 (НМА, ЖВС, возбудимые ткани) Этапы развития человека
Этапы развития человека Минеральные вещества в рационе собаки
Минеральные вещества в рационе собаки Физическая культура и организм человека
Физическая культура и организм человека Детям о комнатных растениях
Детям о комнатных растениях