- Структура ЦНС. Свойства нервных центров

Содержание
- 2. Нервным центром называют совокупность нейронов, участвующих в реализации определенного рефлекса, регуляции той или иной функции или
- 3. Свойства эти следующие: 1.Одностороннее проведение в нервных центрах можно доказать при раздражении передних корешков и отведении
- 4. Центральное облегчение. Возникновение временной и особенно пространственной суммации способствуют и особенности организации синаптического аппарата в нервных
- 5. 7. Последействие. Рефлекторные акты, в отличие от потенциалов действия, заканчиваются не одновременно с прекращением вызвавшего их
- 6. 6.2. МЕТОДЫ ИССЛЕДОВАНИЯ ФУНКЦИЙ ЦНС. Современная физиология располагает большим арсеналом методов, направленных на изучение физиологических свойств
- 7. Электроэнцефалография (ЭЭГ) является современным и достаточно распространенным в клинике методом исследования функций мозга. Метод этот позволяет
- 9. Скачать презентацию
Слайд 2
Нервным центром называют совокупность нейронов, участвующих в реализации определенного рефлекса, регуляции
Нервным центром называют совокупность нейронов, участвующих в реализации определенного рефлекса, регуляции

Слайд 3Свойства эти следующие:
1.Одностороннее проведение в нервных центрах можно доказать при раздражении
Свойства эти следующие:
1.Одностороннее проведение в нервных центрах можно доказать при раздражении

Слайд 4Центральное облегчение. Возникновение временной и особенно пространственной суммации способствуют и особенности организации
Центральное облегчение. Возникновение временной и особенно пространственной суммации способствуют и особенности организации

Слайд 57. Последействие. Рефлекторные акты, в отличие от потенциалов действия, заканчиваются не одновременно
7. Последействие. Рефлекторные акты, в отличие от потенциалов действия, заканчиваются не одновременно

Слайд 66.2. МЕТОДЫ ИССЛЕДОВАНИЯ ФУНКЦИЙ ЦНС.
Современная физиология располагает большим арсеналом методов, направленных
6.2. МЕТОДЫ ИССЛЕДОВАНИЯ ФУНКЦИЙ ЦНС.
Современная физиология располагает большим арсеналом методов, направленных

Слайд 7 Электроэнцефалография (ЭЭГ) является современным и достаточно распространенным в клинике методом исследования
Электроэнцефалография (ЭЭГ) является современным и достаточно распространенным в клинике методом исследования

 Органы чувств человека
Органы чувств человека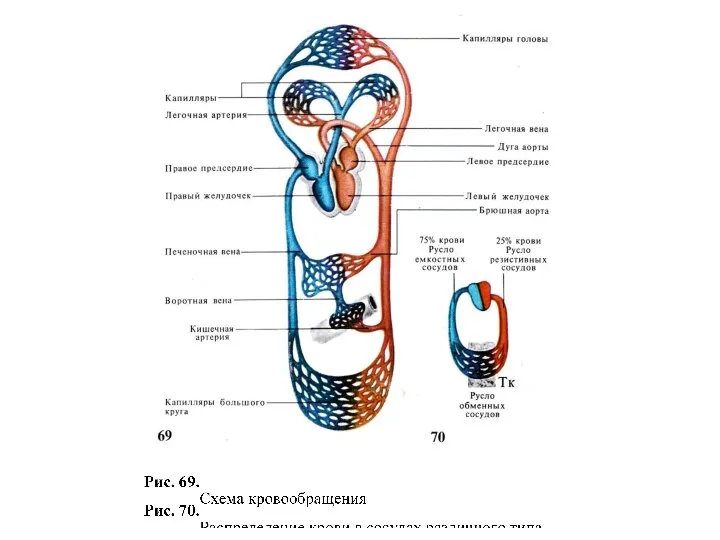 Гемодинамика. Объём крови в различных отделах сердечно-сосудистой системы
Гемодинамика. Объём крови в различных отделах сердечно-сосудистой системы Витамины. Классификация. Группы витаминов
Витамины. Классификация. Группы витаминов Съедобные и несъедобные грибы
Съедобные и несъедобные грибы Катаболиз органических веществ на примере углеводов и жиров
Катаболиз органических веществ на примере углеводов и жиров Лиственница
Лиственница Пчелы - общественные насекомые
Пчелы - общественные насекомые Реакции окисления и восстановления биоорганических соединений. Структура и функции биолекул
Реакции окисления и восстановления биоорганических соединений. Структура и функции биолекул Искусственный отбор
Искусственный отбор Опора и движение
Опора и движение Modern genetics
Modern genetics Типы плодов
Типы плодов Саванна
Саванна Органы дыхания
Органы дыхания Витамины
Витамины Взаимодействие генов
Взаимодействие генов Класс млекопитающие
Класс млекопитающие Расчетная таблица по определению гумусового баланса в севообороте
Расчетная таблица по определению гумусового баланса в севообороте Память, молекулярные механизмы памяти
Память, молекулярные механизмы памяти Воздушное питание, хлоропласты, фотосинтез, растения-паразиты. Питание животных
Воздушное питание, хлоропласты, фотосинтез, растения-паразиты. Питание животных Цікаві факти про черепах
Цікаві факти про черепах Витамин D
Витамин D Едят ли птички сладкое? Всезнайка
Едят ли птички сладкое? Всезнайка Молекулярная биология
Молекулярная биология Опыление 6 класс - Презентация
Опыление 6 класс - Презентация Постэмбриональный период развития организмов
Постэмбриональный период развития организмов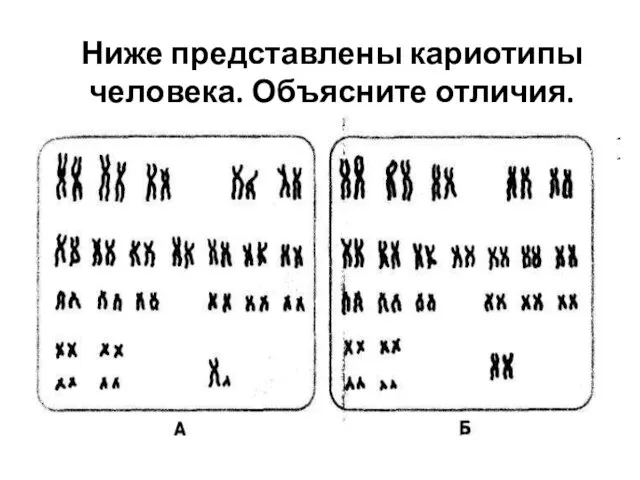 Генетика пола
Генетика пола Обмен веществ и энергии. Лекции 34,35
Обмен веществ и энергии. Лекции 34,35