- Двигательные системы

Содержание
- 2. Физиология мышц Поддержание позы и равновесия тела Произвольные движения Воспроизведение речи Работа сердца Регуляция тонуса сосудов
- 3. Классификация Скелетные, сердечная и гладкие мышцы. Скелетные мышцы принято классифицировать а) По количеству входящих двигательных единиц
- 4. Двигательная единица Комплекс, включающий один мотонейрон и иннервируемые им мышечные волокна, называется ДВИГАТЕЛЬНОЙ ИЛИ НЕЙРОМОТОРНОЙ ЕДИНИЦЕЙ
- 5. Два варианта классификации скелетных мышц Анатомическая По плотности иннервации Функциональная По работоспособности
- 6. Анатомическая классификация по количеству двигательных единиц 1. Много двигательных единиц на одну мышцу (плотность иннервации высокая)
- 7. Функциональная классификация по работоспособности Медленные (малоутомляемые) Быстрые (легкоутомляемые)
- 8. Функциональная характеристика
- 9. Свойства мышцы Возбудимость Проводимость Сократимость Эластичность – способность сокращаться при растягивании Пластичность − способность сохранять приданную
- 10. Возбуждение мышцы .
- 11. Особенности нервно-мышечного синапса (концевой пластинки) Большая поверхность пресинаптической мембраны Большая складчатость постсинаптической мембраны Нет суммации –
- 12. Потенциал концевой пластинки (ПКП) и потенциал действия (ПД) мышечного волокна Синапс ПКП ПД ПД Са++ СПР
- 13. Уровни организации скелетной мышцы
- 14. Микрофотография мышечного волокна
- 15. Строение саркомера Саркомер – функциональная единица сократительного аппарата мышечной клетки. Длина саркомера 2,5 мкм, поперечник –
- 16. Толстые миофиламенты
- 17. Свойства миозина Толстые миофиламенты – образованы молекулами миозина, которые имеют четыре легкие цепи миозина и две
- 18. Тонкие миофиламенты
- 19. Свойства актина Тонкие миофиламенты построены из нитей белка актина. Мономеры актина взаимодействуют между собой и образуют
- 20. Регуляторные белки В продольных бороздках актиновой спирали располагаются нитевидные молекулы белка тропомиозина. Тропомиозин закрывает активные центры
- 21. Механизм скольжения
- 22. Укорочение саркомера
- 23. Последовательность событий при сокращении I этап.ПД пресинаптического окончания приводит к выделению медиатора (электрохимическое сопряжение) Возникновение ПКП
- 24. Временная характеристика процесса сокращения
- 25. Механизм сокращения 1.Са+ выходит из цистерн ЭПР (опыт с экворином, Рюгель 1977) 2.Са+ связывается с тропонином
- 26. Кинематика сокращения
- 27. Виды и режимы мышечного сокращения Виды Изометрическое Изотоническое Ауксотоническое Режимы Одиночное сокращение Тетанус Зубчатый Гладкий Оптимум
- 28. Сопоставление во времени сокращения мышцы и возбудимости Возбудимость мышцы во время сокращения нормальна, следовательно она может
- 29. Расслабление мышцы Са+ удаляется от тропонина и уходит в ЭПР Тропомиозин блокирует актин Поперечные мостики актин-миозин
- 30. Запись мышечных сокращений
- 31. Суммация сокращений
- 32. Режимы мышечного сокращения Одиночное сокращение Тетанус – слитное сокращение без расслабления А)Зубчатый – импульс в фазу
- 33. Регуляция силы сокращения мышцы Сила сокращения зависит от числа включенных нейромоторных единиц; частоты импульсации мотонейронов; синхронизации
- 34. Законы мышечного сокращения 1. Мышечное волокно подчиняется закону «все или ничего» 2. Целая мышца подчиняется закону
- 35. Гладкие мышцы Мелкие (2-10 мкм диаметр, 50-500 мкм длина) Одноядерные, способны к делению Механо- и хемочувствительны
- 37. Скачать презентацию
Слайд 2Физиология мышц
Поддержание позы и равновесия тела
Произвольные движения
Воспроизведение речи
Работа сердца
Регуляция
Физиология мышц
Поддержание позы и равновесия тела
Произвольные движения
Воспроизведение речи
Работа сердца
Регуляция

Слайд 3Классификация
Скелетные, сердечная и гладкие мышцы.
Скелетные мышцы принято классифицировать
а) По количеству входящих
Классификация
Скелетные, сердечная и гладкие мышцы.
Скелетные мышцы принято классифицировать
а) По количеству входящих

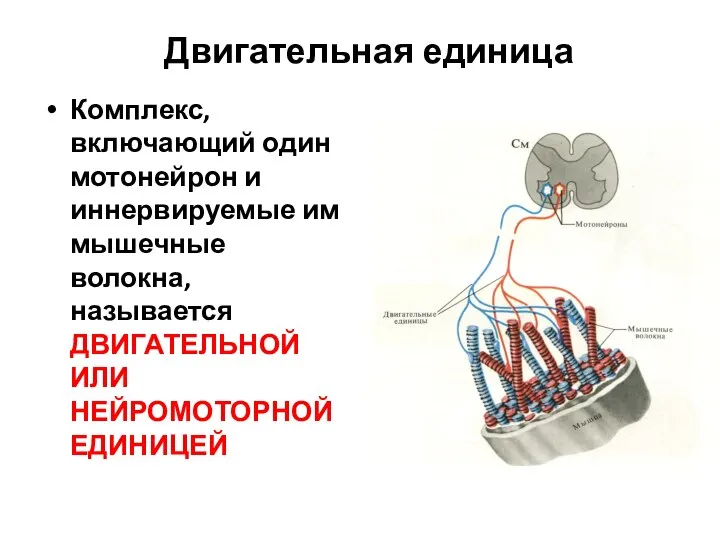
Слайд 4Двигательная единица
Комплекс, включающий один мотонейрон и иннервируемые им мышечные волокна, называется ДВИГАТЕЛЬНОЙ
Двигательная единица
Комплекс, включающий один мотонейрон и иннервируемые им мышечные волокна, называется ДВИГАТЕЛЬНОЙ
Слайд 5Два варианта классификации
скелетных мышц
Анатомическая
По плотности иннервации
Функциональная
По работоспособности
Два варианта классификации
скелетных мышц
Анатомическая
По плотности иннервации
Функциональная
По работоспособности

Слайд 6Анатомическая классификация
по количеству двигательных единиц
1. Много двигательных единиц на одну мышцу
Анатомическая классификация
по количеству двигательных единиц
1. Много двигательных единиц на одну мышцу

Слайд 7Функциональная классификация
по работоспособности
Медленные (малоутомляемые)
Быстрые (легкоутомляемые)
Функциональная классификация
по работоспособности
Медленные (малоутомляемые)
Быстрые (легкоутомляемые)

Слайд 8Функциональная характеристика
Функциональная характеристика

Слайд 9Свойства мышцы
Возбудимость
Проводимость
Сократимость
Эластичность – способность сокращаться при растягивании
Пластичность − способность сохранять приданную растяжением
Свойства мышцы
Возбудимость
Проводимость
Сократимость
Эластичность – способность сокращаться при растягивании
Пластичность − способность сохранять приданную растяжением

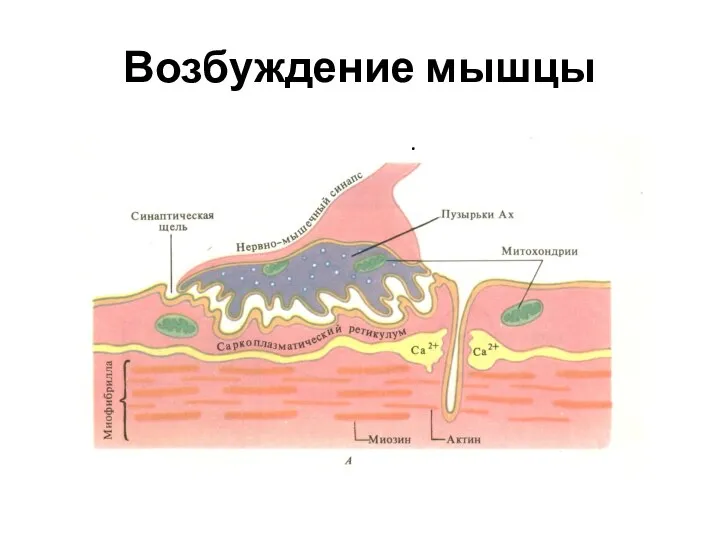
Слайд 10Возбуждение мышцы
.
Возбуждение мышцы
.
Слайд 11Особенности
нервно-мышечного синапса
(концевой пластинки)
Большая поверхность пресинаптической мембраны
Большая складчатость постсинаптической мембраны
Нет суммации
Особенности
нервно-мышечного синапса
(концевой пластинки)
Большая поверхность пресинаптической мембраны
Большая складчатость постсинаптической мембраны
Нет суммации

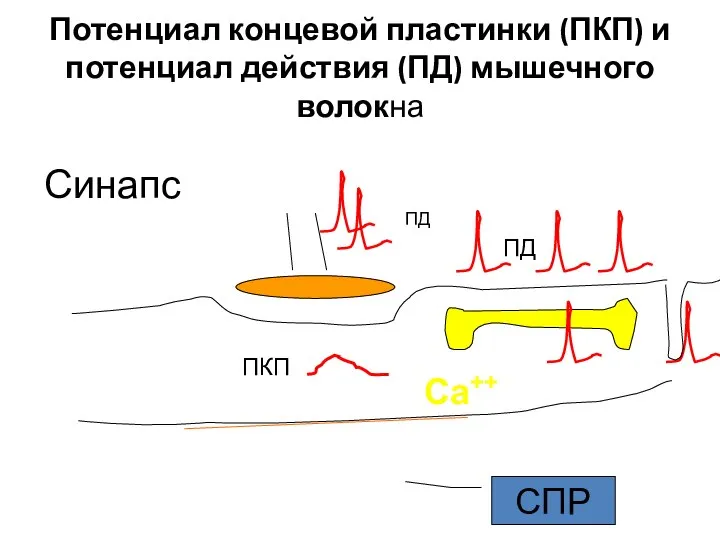
Слайд 12Потенциал концевой пластинки (ПКП) и потенциал действия (ПД) мышечного волокна
Синапс
ПКП
ПД
ПД
Са++
СПР
Потенциал концевой пластинки (ПКП) и потенциал действия (ПД) мышечного волокна
Синапс
ПКП
ПД
ПД
Са++
СПР
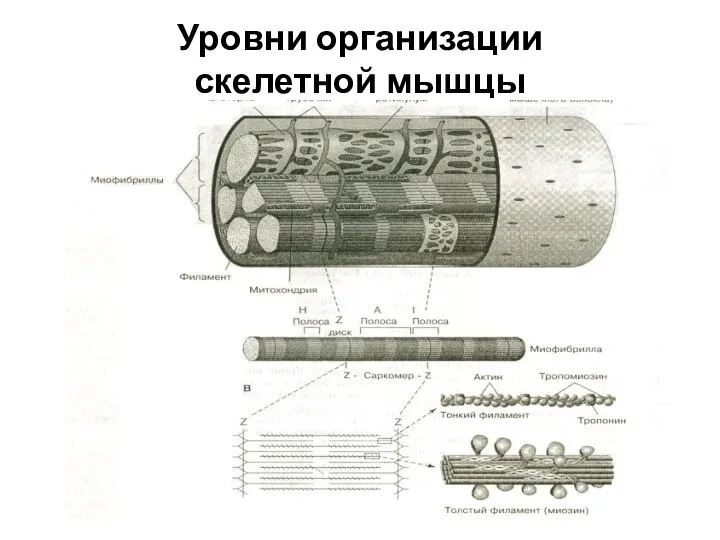
Слайд 13Уровни организации
скелетной мышцы
Уровни организации
скелетной мышцы
Слайд 14Микрофотография мышечного волокна
Микрофотография мышечного волокна

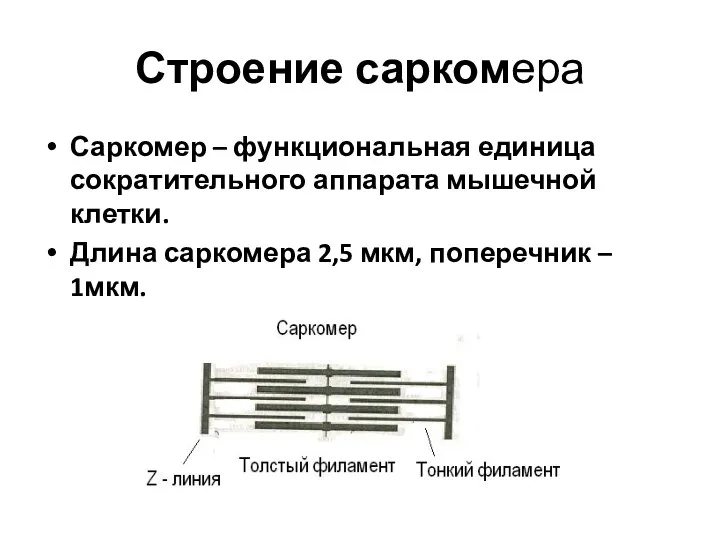
Слайд 15Строение саркомера
Саркомер – функциональная единица сократительного аппарата мышечной клетки.
Длина саркомера 2,5 мкм,
Строение саркомера
Саркомер – функциональная единица сократительного аппарата мышечной клетки.
Длина саркомера 2,5 мкм,
Слайд 16Толстые миофиламенты
Толстые миофиламенты
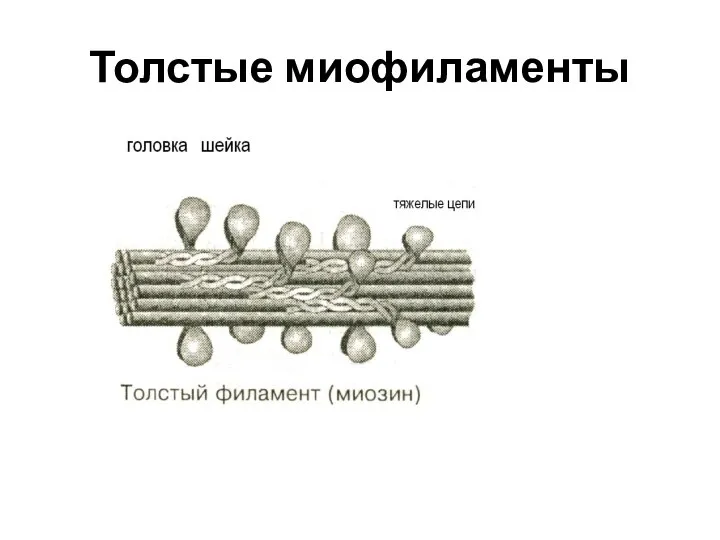
Слайд 17Свойства миозина
Толстые миофиламенты – образованы молекулами миозина, которые имеют четыре легкие цепи
Свойства миозина
Толстые миофиламенты – образованы молекулами миозина, которые имеют четыре легкие цепи

Слайд 18Тонкие миофиламенты
Тонкие миофиламенты
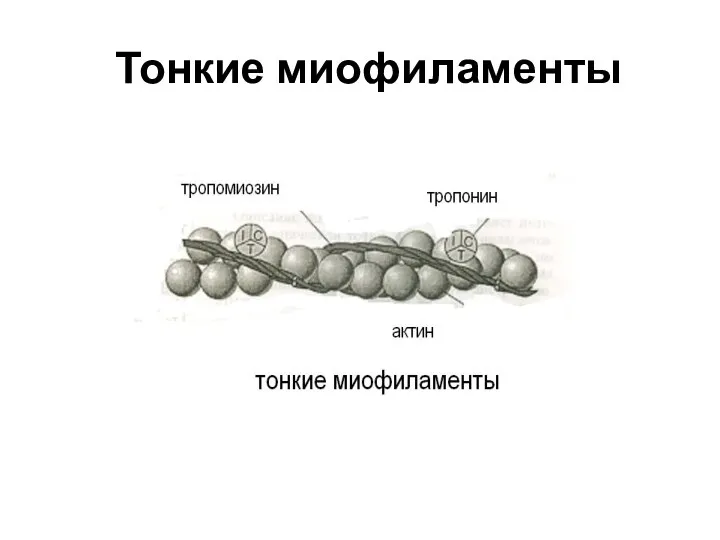
Слайд 19Свойства актина
Тонкие миофиламенты построены из нитей белка актина.
Мономеры актина взаимодействуют между
Свойства актина
Тонкие миофиламенты построены из нитей белка актина.
Мономеры актина взаимодействуют между

Слайд 20Регуляторные белки
В продольных бороздках актиновой спирали располагаются нитевидные молекулы белка тропомиозина.
Тропомиозин
Регуляторные белки
В продольных бороздках актиновой спирали располагаются нитевидные молекулы белка тропомиозина.
Тропомиозин

Слайд 21Механизм скольжения
Механизм скольжения

Слайд 22Укорочение саркомера
Укорочение саркомера

Слайд 23Последовательность событий при сокращении
I этап.ПД пресинаптического окончания приводит к выделению медиатора
Последовательность событий при сокращении
I этап.ПД пресинаптического окончания приводит к выделению медиатора

Слайд 24Временная характеристика процесса сокращения
Временная характеристика процесса сокращения

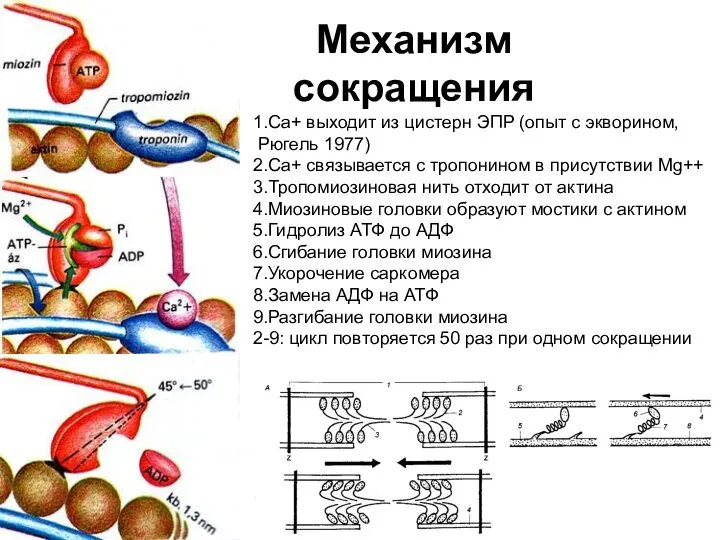
Слайд 25Механизм
сокращения
1.Са+ выходит из цистерн ЭПР (опыт с экворином,
Рюгель 1977)
2.Са+ связывается
Механизм
сокращения
1.Са+ выходит из цистерн ЭПР (опыт с экворином,
Рюгель 1977)
2.Са+ связывается
Слайд 26Кинематика сокращения
Кинематика сокращения
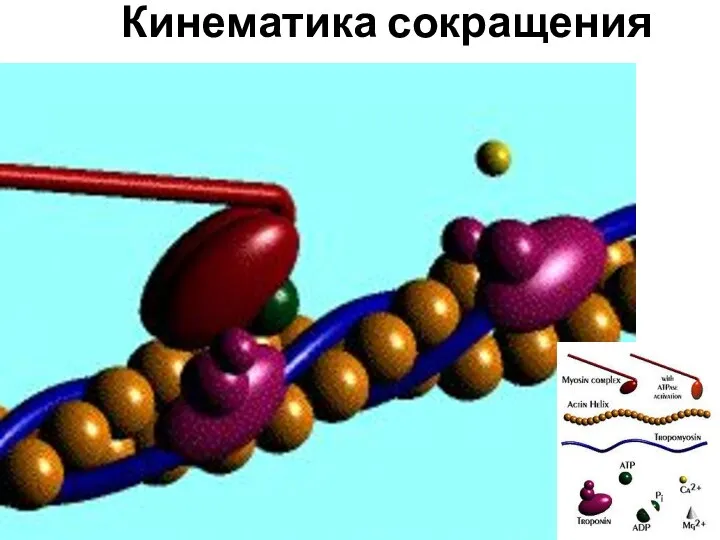
Слайд 27Виды и режимы мышечного сокращения
Виды
Изометрическое
Изотоническое
Ауксотоническое
Режимы
Одиночное сокращение
Тетанус
Зубчатый
Гладкий
Оптимум
Пессимум
Виды и режимы мышечного сокращения
Виды
Изометрическое
Изотоническое
Ауксотоническое
Режимы
Одиночное сокращение
Тетанус
Зубчатый
Гладкий
Оптимум
Пессимум

Слайд 28Сопоставление во времени сокращения мышцы и возбудимости
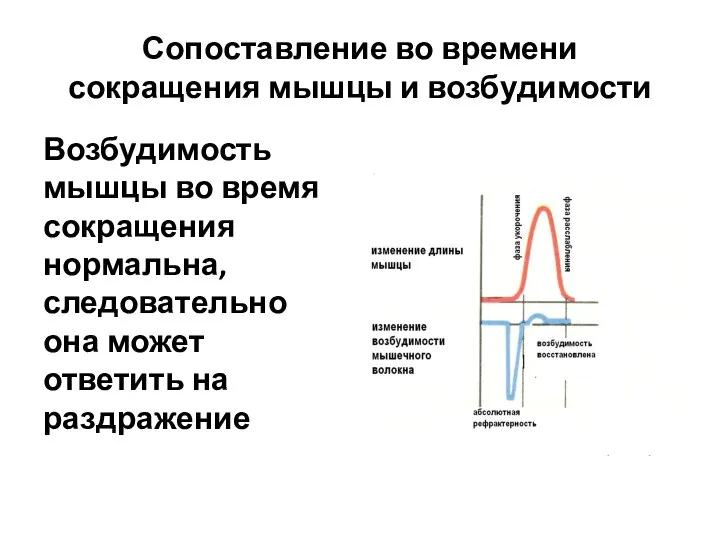
Возбудимость мышцы во время сокращения нормальна,
Сопоставление во времени сокращения мышцы и возбудимости
Возбудимость мышцы во время сокращения нормальна,
Слайд 29Расслабление мышцы
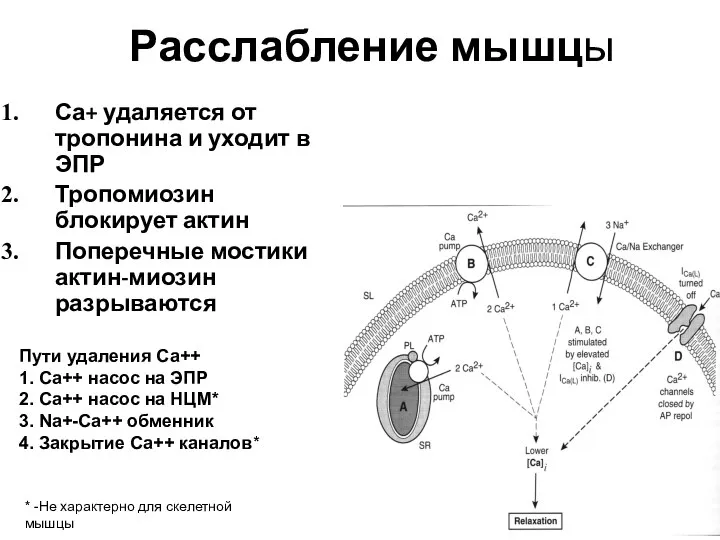
Са+ удаляется от тропонина и уходит в ЭПР
Тропомиозин блокирует актин
Поперечные мостики
Расслабление мышцы
Са+ удаляется от тропонина и уходит в ЭПР
Тропомиозин блокирует актин
Поперечные мостики
Слайд 30Запись мышечных сокращений
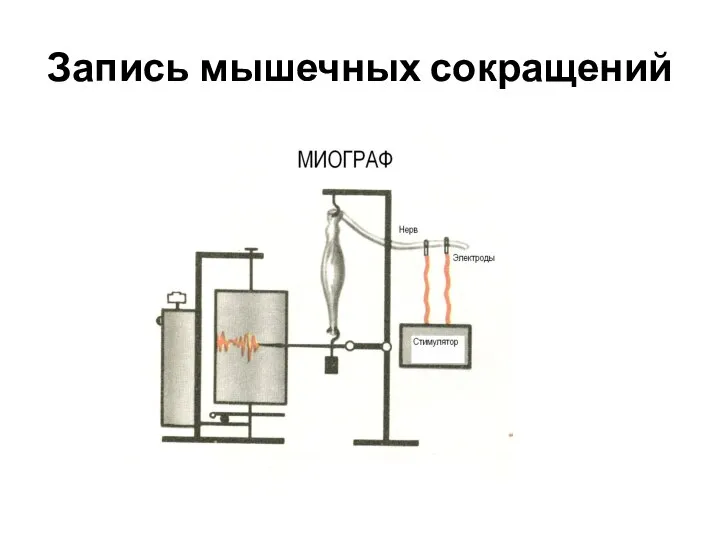
Запись мышечных сокращений
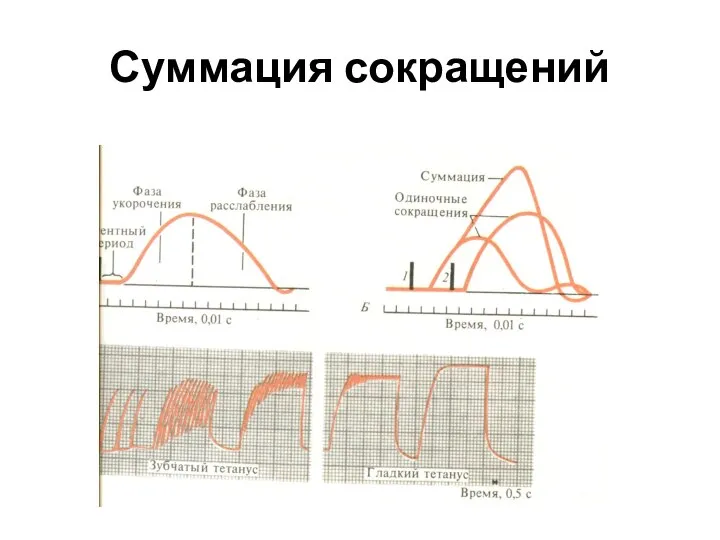
Слайд 31Суммация сокращений
Суммация сокращений
Слайд 32Режимы
мышечного сокращения
Одиночное сокращение
Тетанус – слитное сокращение без расслабления
А)Зубчатый – импульс в фазу
Режимы
мышечного сокращения
Одиночное сокращение
Тетанус – слитное сокращение без расслабления
А)Зубчатый – импульс в фазу

Слайд 33Регуляция силы сокращения мышцы
Сила сокращения зависит от
числа включенных нейромоторных единиц;
частоты импульсации
Регуляция силы сокращения мышцы
Сила сокращения зависит от
числа включенных нейромоторных единиц;
частоты импульсации

Слайд 34Законы мышечного сокращения
1. Мышечное волокно подчиняется закону «все или ничего»
2. Целая мышца
Законы мышечного сокращения
1. Мышечное волокно подчиняется закону «все или ничего»
2. Целая мышца

Слайд 35Гладкие мышцы
Мелкие (2-10 мкм диаметр, 50-500 мкм длина)
Одноядерные, способны к делению
Механо- и
Гладкие мышцы
Мелкие (2-10 мкм диаметр, 50-500 мкм длина)
Одноядерные, способны к делению
Механо- и

 Экология синиц
Экология синиц Строение клетки
Строение клетки Популяционная морфология и генетика. Актуальные направления исследований в популяционной генетике
Популяционная морфология и генетика. Актуальные направления исследований в популяционной генетике Белки, состав, структура, функции
Белки, состав, структура, функции Как размножаются живые организмы. 6 класс
Как размножаются живые организмы. 6 класс Нервная система
Нервная система Презентация на тему Увеличительные приборы и правила работы с ними
Презентация на тему Увеличительные приборы и правила работы с ними  Анатомо-физиологические особенности подростковом возрасте. 7 класс
Анатомо-физиологические особенности подростковом возрасте. 7 класс Комнатные растения
Комнатные растения Нервная ткань
Нервная ткань Анатомия передней черепной ямки
Анатомия передней черепной ямки Автор: учитель биологии Константинова Елена Анатольевна
Автор: учитель биологии Константинова Елена Анатольевна Dostovalov_Arseniy
Dostovalov_Arseniy Цветок одуванчик
Цветок одуванчик Законы Менделя
Законы Менделя Внутреннее строение насекомых
Внутреннее строение насекомых Биокатализ и витамины
Биокатализ и витамины Анатолийский карабаш
Анатолийский карабаш Какой тип газообмена у амебы и дождевого червя?
Какой тип газообмена у амебы и дождевого червя? Тірі жүйелердегі зат және энергия алмасу типтері
Тірі жүйелердегі зат және энергия алмасу типтері Терморегуляция организма. Закаливание
Терморегуляция организма. Закаливание BRAINWASH
BRAINWASH Мир собак. Как правильно выбрать питомца
Мир собак. Как правильно выбрать питомца Предмет, значение и краткая история развития микробиологии, роль отечественных ученых. Систематика микроорганизмов
Предмет, значение и краткая история развития микробиологии, роль отечественных ученых. Систематика микроорганизмов Аттестационная работа. Методическая разработка по сопровождению проектной работы 5 кл. Биологическая модель, процесс, явление
Аттестационная работа. Методическая разработка по сопровождению проектной работы 5 кл. Биологическая модель, процесс, явление Животный мир
Животный мир Митоз. Этапы митоза
Митоз. Этапы митоза Протеомика. Массспектрометрия
Протеомика. Массспектрометрия