- Энзимология III. Регуляция активности

Содержание
- 2. План лекции Общие принципы и способы регуляции активности ферментов Стадии ферментативного катализа Механизм действия ферментов Понятие
- 3. Роль РЕГУЛЯЦИИ ферментативных реакций В живой клетке множество разнообразных соединений, но реакции между ними не беспорядочны,
- 4. Основные принципы и механизмы регуляции ферментативных реакций регулирование скорости ферментативной реакции путем изменения количества фермента регулирование
- 5. СТАДИИ ФЕРМЕНТАТИВНОГО КАТАЛИЗА E +S - узнавание ферментом (Е) субстрата (S) ES – образование комплекса и
- 7. Механизмы действия ферментов Первоначальным событием при действии фермента является его специфическое связывание с лигандом - субстратом
- 8. ТЕОРИЯ ЖЕСТКОЙ МАТРИЦЫ Теория Фишера (модель "жесткой матрицы", "ключ-замок") – активный центр фермента строго соответствует конфигурации
- 9. ТЕОРИЯ ИНДУЦИРОВАННОГО ВЗАИМОДЕЙСТВИЯ В 1958 г. Дениел Кошланд предложил модель индуцированного взаимодействия. Ферменты, в основном, —
- 10. Кинетические свойства ферментов Кинетика ферментативных реакций – наука о скоростях ферментативных реакций, их зависимости от различных
- 11. Зависимость скорости ферментативной реакции от условий
- 12. Зависимость скорости ферментативной реакции от температуры
- 13. Оптимум рН соответствует максимальной степени ионизации функциональных групп фермента ( для большинства ферментов около 7.0, но
- 14. Регуляция ферментативной активности
- 15. ВИДЫ ИНГИБИРОВАНИЯ Ингибиторы способны взаимодействовать с ферментами с разной степенью прочности. На основании этого различают обратимое
- 16. Ингибиторы ферментов Действие ферментов можно полностью или частично подавить (ингибировать) определенными химическими веществами (ингибиторами). По характеру
- 17. Диизопропилфторфосфат ингибирует фермент, ацетилхолинэстеразу, имеющие серин в активном центре. Реакция проведения нервного импульса (происходит каждый раз,
- 18. Диизопропилфторфосфат - отравляющее вещество нервно-паралитического действия ( пример НЕОБРАТИМОГО ИНГИБИРОВАНИЯ). Приводит к утрате способности нейронов проводить
- 19. Механизм действия ацетилсалициловой кислоты (необратимое ингибирование) Терапевтическое действие аспирина как жаропонижающего и противовоспалительного средства объясняется тем,
- 20. Механизм действия ацетилсалициловой кислоты
- 21. Обратимые ингибиторы Существует два типа обратимых ингибиторов - конкурентные и неконкурентные. Конкурентный ингибитор конкурирует с субстратом
- 22. В отличие от субстрата связанный с ферментом конкурентный ингибитор не подвергается ферментативному превращению. Более того, образование
- 23. Конкурентными ингибиторами являются многие химиотерапевтические средства. Пример: сульфамидные препараты, используемые для лечения инфекционных болезней. . Сульфаниламиды
- 24. Механизм действия сульфаниламидных препаратов
- 25. Неконкурентное обратимое ингибирование - ингибиторы присоединяются к ферменту не в активном центре, а в другом месте
- 26. В живой клетке множество разнообразных соединений, но реакции между ними не беспорядочны, а образуют строго определенные
- 27. В живой клетке скорость ферментативных реакций находится под строгим контролем Метаболическая цепь: А, В, С, D
- 28. В каждой метаболической цепи есть фермент, который задает скорость всей цепочке реакций. Он называется регуляторным ферментом.
- 29. Основные способы изменения активности ферментов . Аллостерическая регуляция. Регуляция активности ферментов путем фосфорилирования-дефосфорилирования (путем ковалентной модификации).
- 30. Аллостерическая регуляция активности фермента «Сообщение» о присоединении аллостерического активатора передается посредством конформационных изменений каталитической субъединице, которая
- 31. Регуляция активности ферментов путем фосфорилирования-дефосфорилирования Фермент изменяет активность в результате ковалентной модификации.
- 32. Регуляция путем ассоциации-диссоциации субъединиц в олигомерном ферменте Этот процесс иногда начинается с ковалентной или нековалентной модификации
- 33. Аденилатциклазная система Аденилатциклаза и протеинкиназа катализируют взаимосвязанные реакции, которые составляют единую регуляторную систему.
- 34. В живой клетке скорость ферментативных реакций зависит от количества ферментов Количество фермента в клетке определяется соотношением
- 35. Между количеством фермента и скоростью ферментативной реакции существует прямая зависимость, т.е. с увеличением количества фермента скорость
- 36. С увеличением количества фермента скорость реакции увеличивается прямопропорционально
- 37. Контроль количества фермента Путь регуляции ферментативных реакций через изменение количества фермента является путем длительной адаптации метаболических
- 38. Индуцибельные ферменты - ферменты адаптации К индуцибельным ферментам относят ферменты метаболизма чужеродных веществ (монооксигеназы), синтез которых
- 39. Схема индукции ферментов Субстрат А, присутствующий в высокой концентрации, стимулирует синтез ферментов Ф1, Ф2 и Ф3,
- 40. Схема репрессии синтеза ферментов Метаболическая цепь: А, В, С, D - метаболиты, Е1, Е2, Е3, Е4
- 41. Некоторые ферменты синтезируются первоначально неактивными и лишь после секреции из клетки переходят в активную форму. Неактивный
- 42. В физиологическом диапазоне концентраций существует прямая зависимость между количеством субстрата и скоростью ферментативной реакции, т.е. при
- 43. Содержание субстрата в клетке зависит от следующих факторов: 1.от состояния проницаемости мембран для субстратов, поступающих извне
- 44. Компартментализация Регуляция химических реакций осуществляется также за счет пространственного отделения внутриклеточными мембранами одних процессов от других.
- 45. Изоферменты Изоферменты - это множественные формы ферментов, которые катализируют одну и ту же реакцию, но отличаются
- 46. Изменения спектра ЛДГ в онтогенезе Соотношение разных форм ЛДГ в тканях не является постоянным и изменяется
- 48. Скачать презентацию
Слайд 2План лекции
Общие принципы и способы регуляции активности ферментов
Стадии ферментативного катализа
Механизм действия ферментов
Понятие
План лекции
Общие принципы и способы регуляции активности ферментов
Стадии ферментативного катализа
Механизм действия ферментов
Понятие

Слайд 3 Роль РЕГУЛЯЦИИ ферментативных
реакций
В живой клетке множество разнообразных соединений, но реакции
Роль РЕГУЛЯЦИИ ферментативных
реакций
В живой клетке множество разнообразных соединений, но реакции

Слайд 4Основные принципы и механизмы регуляции ферментативных реакций
регулирование скорости ферментативной реакции путем изменения
Основные принципы и механизмы регуляции ферментативных реакций
регулирование скорости ферментативной реакции путем изменения

Слайд 5СТАДИИ ФЕРМЕНТАТИВНОГО КАТАЛИЗА
E +S - узнавание ферментом (Е) субстрата (S)
ES –
СТАДИИ ФЕРМЕНТАТИВНОГО КАТАЛИЗА
E +S - узнавание ферментом (Е) субстрата (S)
ES –

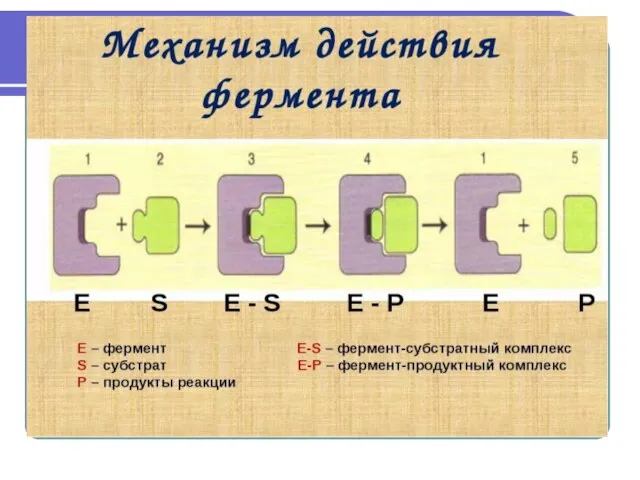
Слайд 7Механизмы действия ферментов
Первоначальным событием при действии фермента является его специфическое связывание с
Механизмы действия ферментов
Первоначальным событием при действии фермента является его специфическое связывание с

Слайд 8ТЕОРИЯ ЖЕСТКОЙ МАТРИЦЫ
Теория Фишера (модель "жесткой матрицы", "ключ-замок") – активный центр фермента строго
ТЕОРИЯ ЖЕСТКОЙ МАТРИЦЫ
Теория Фишера (модель "жесткой матрицы", "ключ-замок") – активный центр фермента строго

Слайд 9ТЕОРИЯ ИНДУЦИРОВАННОГО ВЗАИМОДЕЙСТВИЯ
В 1958 г. Дениел Кошланд предложил модель индуцированного взаимодействия.
Ферменты, в основном, —
ТЕОРИЯ ИНДУЦИРОВАННОГО ВЗАИМОДЕЙСТВИЯ
В 1958 г. Дениел Кошланд предложил модель индуцированного взаимодействия.
Ферменты, в основном, —

Слайд 10Кинетические свойства ферментов
Кинетика ферментативных реакций – наука о скоростях ферментативных реакций, их
Кинетические свойства ферментов
Кинетика ферментативных реакций – наука о скоростях ферментативных реакций, их

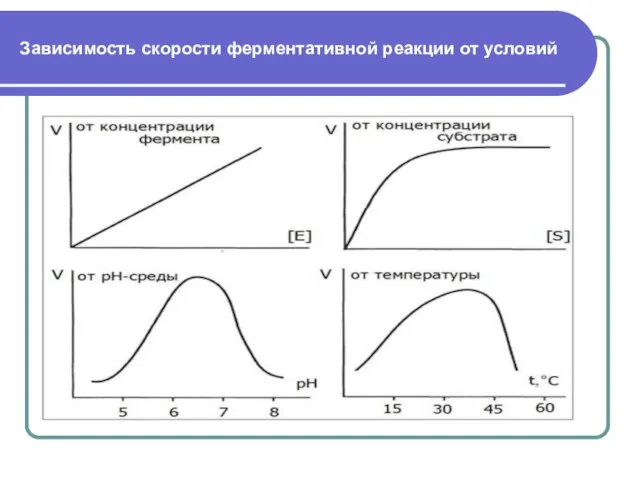
Слайд 11Зависимость скорости ферментативной реакции от условий
Зависимость скорости ферментативной реакции от условий
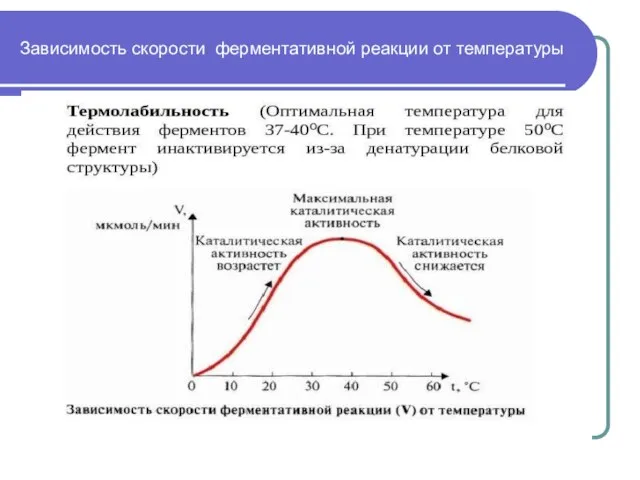
Слайд 12Зависимость скорости ферментативной реакции от температуры
Зависимость скорости ферментативной реакции от температуры
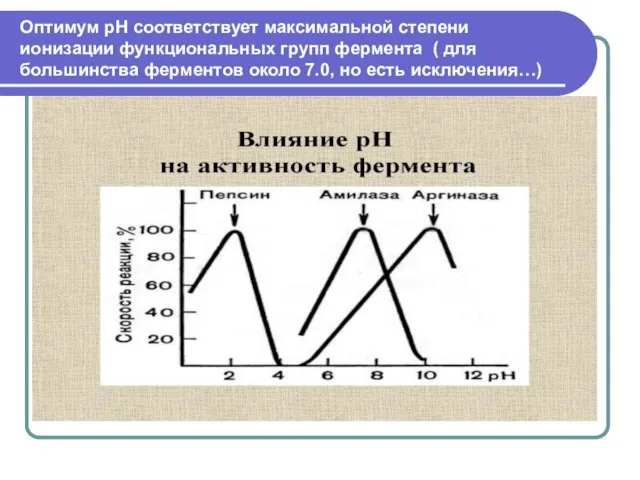
Слайд 13Оптимум рН соответствует максимальной степени ионизации функциональных групп фермента ( для большинства
Оптимум рН соответствует максимальной степени ионизации функциональных групп фермента ( для большинства
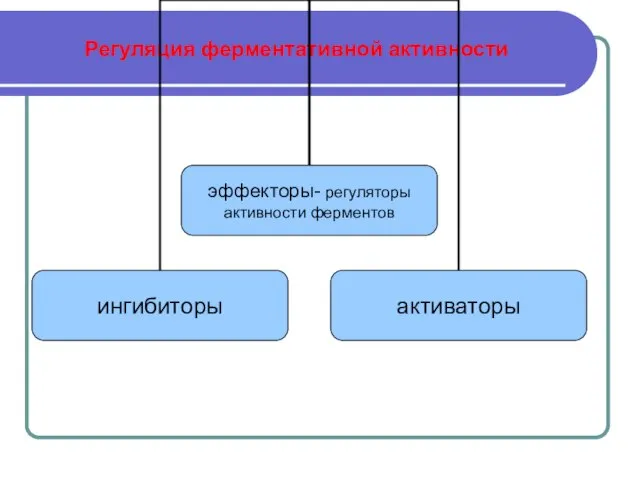
Слайд 14 Регуляция ферментативной активности
Регуляция ферментативной активности
Слайд 15ВИДЫ ИНГИБИРОВАНИЯ
Ингибиторы способны взаимодействовать с ферментами с разной степенью прочности. На основании
ВИДЫ ИНГИБИРОВАНИЯ
Ингибиторы способны взаимодействовать с ферментами с разной степенью прочности. На основании

Слайд 16Ингибиторы ферментов
Действие ферментов можно полностью или частично подавить (ингибировать) определенными химическими веществами
Ингибиторы ферментов
Действие ферментов можно полностью или частично подавить (ингибировать) определенными химическими веществами

Слайд 17
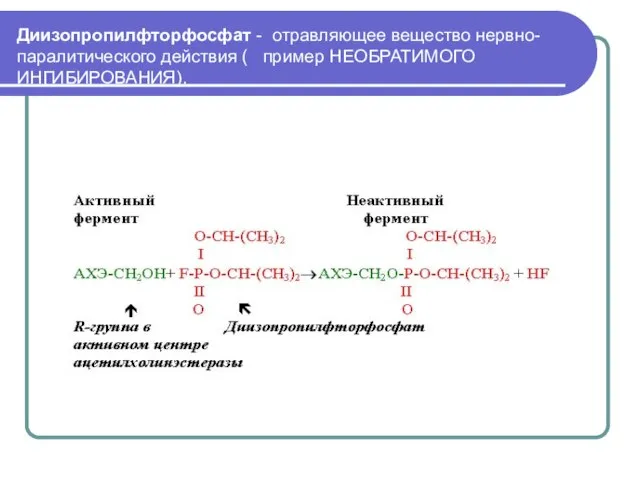
Диизопропилфторфосфат ингибирует фермент, ацетилхолинэстеразу, имеющие серин в активном центре.
Реакция проведения нервного
Диизопропилфторфосфат ингибирует фермент, ацетилхолинэстеразу, имеющие серин в активном центре.
Реакция проведения нервного

Слайд 18Диизопропилфторфосфат - отравляющее вещество нервно-паралитического действия ( пример НЕОБРАТИМОГО ИНГИБИРОВАНИЯ).
Приводит к
Диизопропилфторфосфат - отравляющее вещество нервно-паралитического действия ( пример НЕОБРАТИМОГО ИНГИБИРОВАНИЯ).
Приводит к
Слайд 19Механизм действия ацетилсалициловой кислоты (необратимое ингибирование)
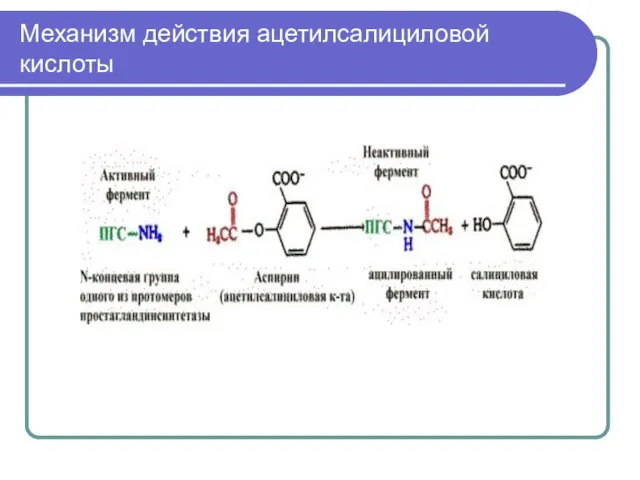
Терапевтическое действие аспирина как жаропонижающего и противовоспалительного
Механизм действия ацетилсалициловой кислоты (необратимое ингибирование)
Терапевтическое действие аспирина как жаропонижающего и противовоспалительного

Слайд 20Механизм действия ацетилсалициловой кислоты
Механизм действия ацетилсалициловой кислоты
Слайд 21Обратимые ингибиторы
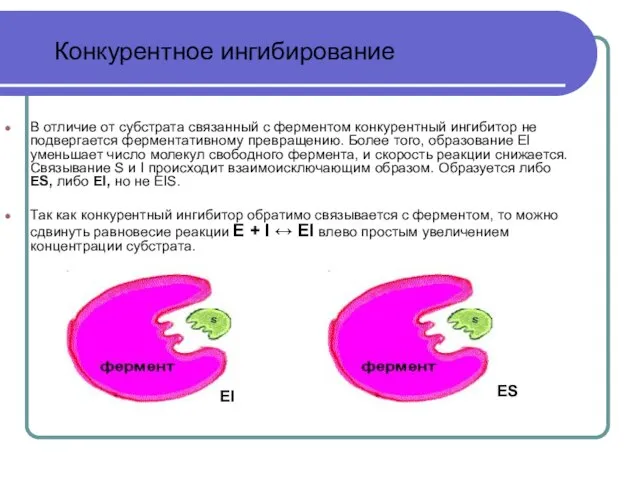
Существует два типа обратимых ингибиторов - конкурентные и неконкурентные.
Конкурентный ингибитор
Обратимые ингибиторы
Существует два типа обратимых ингибиторов - конкурентные и неконкурентные.
Конкурентный ингибитор

Слайд 22В отличие от субстрата связанный с ферментом конкурентный ингибитор не подвергается ферментативному
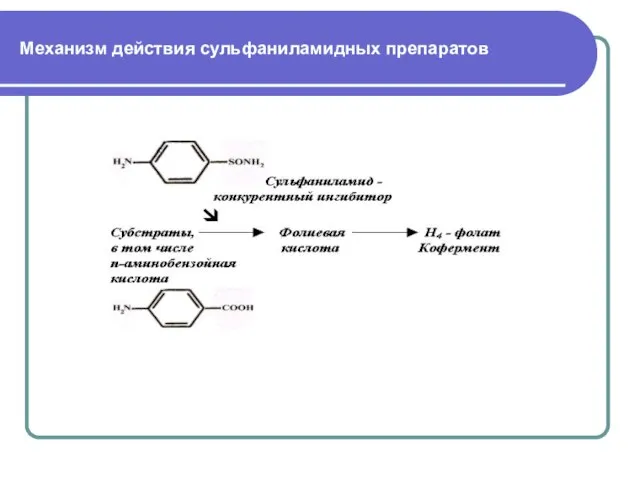
Слайд 23Конкурентными ингибиторами являются многие химиотерапевтические средства. Пример: сульфамидные препараты, используемые для лечения
Конкурентными ингибиторами являются многие химиотерапевтические средства. Пример: сульфамидные препараты, используемые для лечения

Слайд 24Механизм действия сульфаниламидных препаратов
Механизм действия сульфаниламидных препаратов
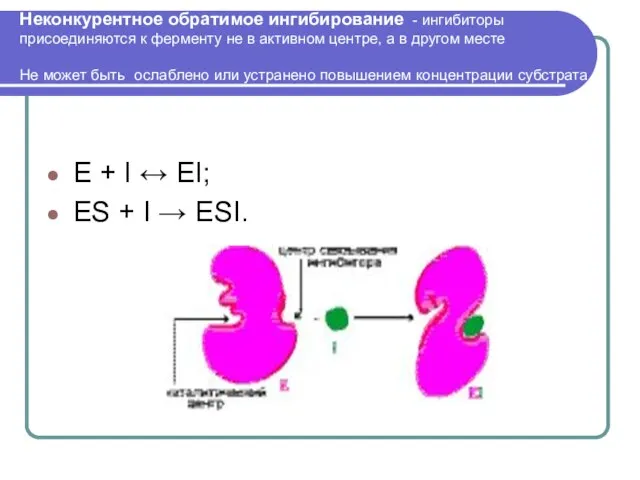
Слайд 25Неконкурентное обратимое ингибирование - ингибиторы присоединяются к ферменту не в активном центре,
Неконкурентное обратимое ингибирование - ингибиторы присоединяются к ферменту не в активном центре,
Слайд 26В живой клетке множество разнообразных соединений, но реакции между ними не беспорядочны,
В живой клетке множество разнообразных соединений, но реакции между ними не беспорядочны,

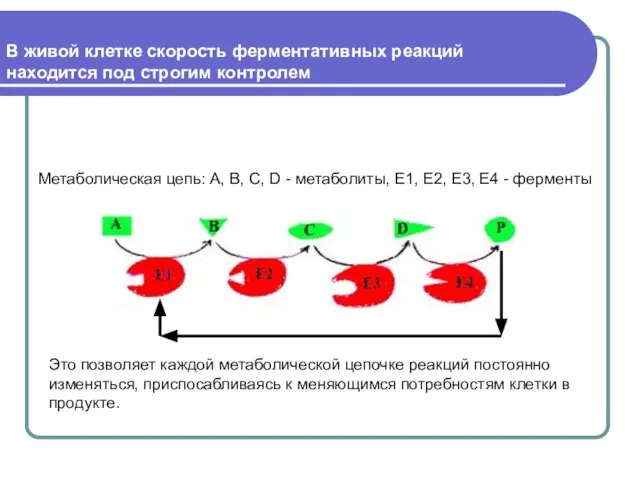
Слайд 27В живой клетке скорость ферментативных реакций находится под строгим контролем
Метаболическая цепь: А,
В живой клетке скорость ферментативных реакций находится под строгим контролем
Метаболическая цепь: А,
Слайд 28В каждой метаболической цепи есть фермент, который задает скорость всей цепочке реакций.
В каждой метаболической цепи есть фермент, который задает скорость всей цепочке реакций.

Слайд 29Основные способы изменения активности ферментов
.
Аллостерическая регуляция.
Регуляция активности ферментов путем фосфорилирования-дефосфорилирования (путем
Основные способы изменения активности ферментов
.
Аллостерическая регуляция.
Регуляция активности ферментов путем фосфорилирования-дефосфорилирования (путем

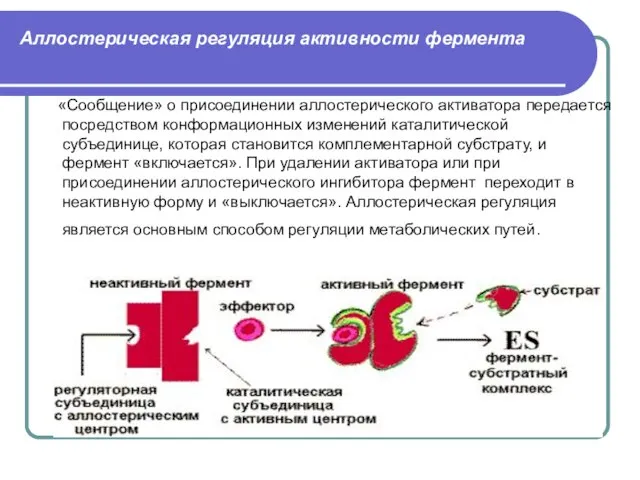
Слайд 30Аллостерическая регуляция активности фермента
«Сообщение» о присоединении аллостерического активатора передается посредством конформационных
Аллостерическая регуляция активности фермента
«Сообщение» о присоединении аллостерического активатора передается посредством конформационных
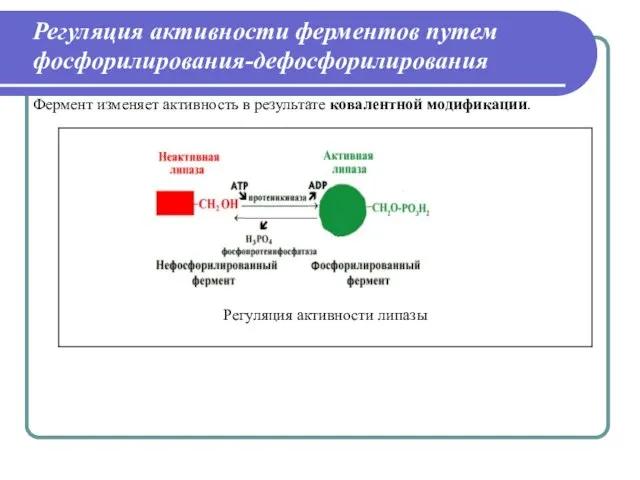
Слайд 31Регуляция активности ферментов путем фосфорилирования-дефосфорилирования
Фермент изменяет активность в результате ковалентной модификации.
Регуляция активности ферментов путем фосфорилирования-дефосфорилирования
Фермент изменяет активность в результате ковалентной модификации.
Слайд 32Регуляция путем ассоциации-диссоциации субъединиц в олигомерном ферменте
Этот процесс иногда начинается с
Регуляция путем ассоциации-диссоциации субъединиц в олигомерном ферменте
Этот процесс иногда начинается с

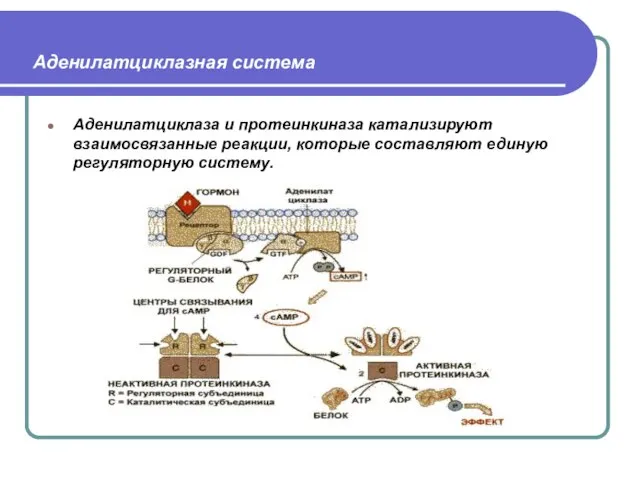
Слайд 33
Аденилатциклазная система
Аденилатциклаза и протеинкиназа катализируют взаимосвязанные реакции, которые составляют единую регуляторную систему.
Аденилатциклазная система
Аденилатциклаза и протеинкиназа катализируют взаимосвязанные реакции, которые составляют единую регуляторную систему.
Слайд 34В живой клетке скорость ферментативных реакций зависит от количества ферментов
Количество фермента в
В живой клетке скорость ферментативных реакций зависит от количества ферментов
Количество фермента в

Слайд 35Между количеством фермента и скоростью ферментативной реакции существует прямая зависимость, т.е. с
Между количеством фермента и скоростью ферментативной реакции существует прямая зависимость, т.е. с

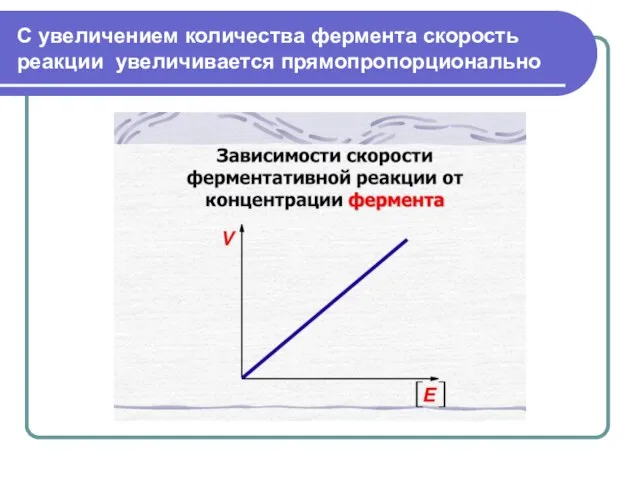
Слайд 36С увеличением количества фермента скорость реакции увеличивается прямопропорционально
С увеличением количества фермента скорость реакции увеличивается прямопропорционально
Слайд 37 Контроль количества фермента
Путь регуляции ферментативных реакций через изменение количества фермента является
Контроль количества фермента
Путь регуляции ферментативных реакций через изменение количества фермента является

Слайд 38Индуцибельные ферменты - ферменты адаптации
К индуцибельным ферментам относят ферменты метаболизма чужеродных веществ
Индуцибельные ферменты - ферменты адаптации
К индуцибельным ферментам относят ферменты метаболизма чужеродных веществ

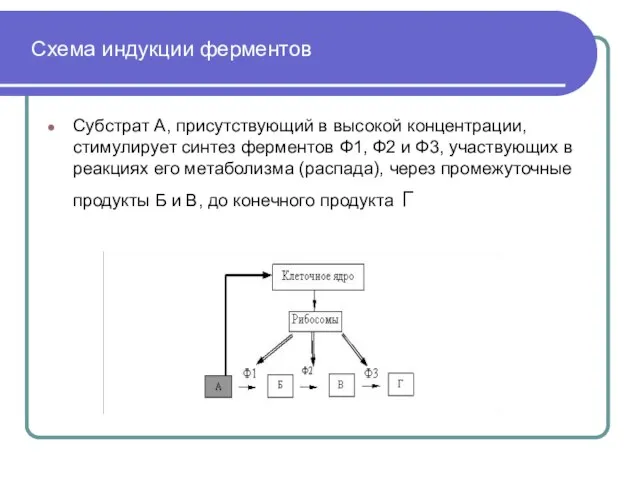
Слайд 39 Схема индукции ферментов
Субстрат А, присутствующий в высокой концентрации, стимулирует синтез ферментов
Схема индукции ферментов
Субстрат А, присутствующий в высокой концентрации, стимулирует синтез ферментов
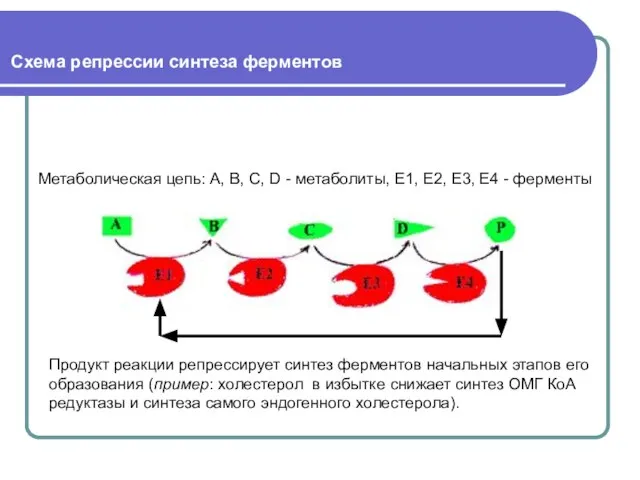
Слайд 40 Схема репрессии синтеза ферментов
Метаболическая цепь: А, В, С, D - метаболиты,
Схема репрессии синтеза ферментов
Метаболическая цепь: А, В, С, D - метаболиты,
Слайд 41
Некоторые ферменты синтезируются первоначально неактивными и лишь после секреции из клетки переходят
Некоторые ферменты синтезируются первоначально неактивными и лишь после секреции из клетки переходят

Слайд 42В физиологическом диапазоне концентраций существует прямая зависимость между количеством субстрата и скоростью
В физиологическом диапазоне концентраций существует прямая зависимость между количеством субстрата и скоростью

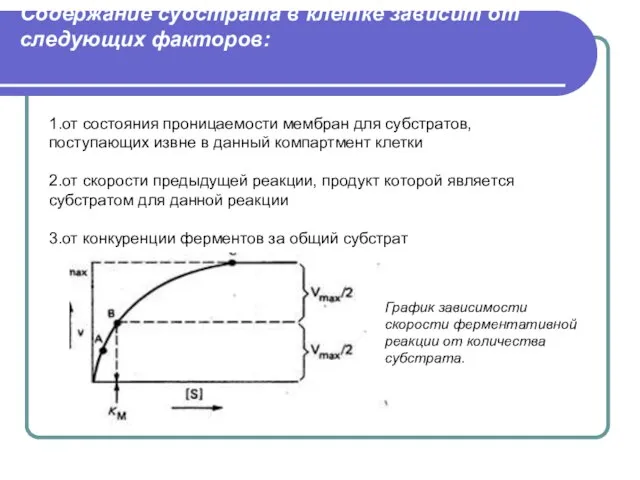
Слайд 43Содержание субстрата в клетке зависит от следующих факторов:
1.от состояния проницаемости мембран для
Содержание субстрата в клетке зависит от следующих факторов:
1.от состояния проницаемости мембран для
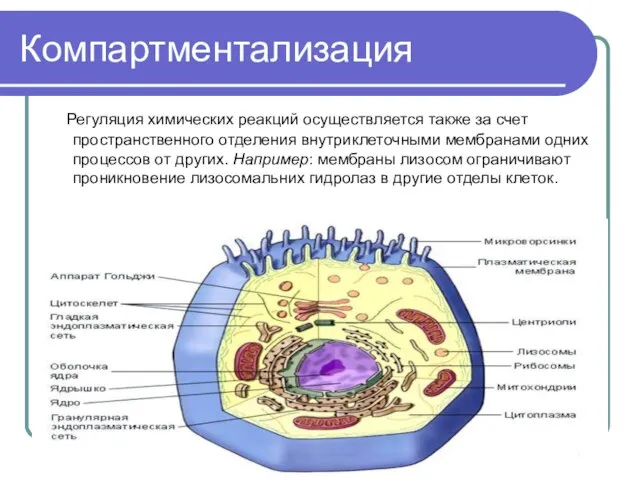
Слайд 44Компартментализация
Регуляция химических реакций осуществляется также за счет пространственного отделения внутриклеточными мембранами
Компартментализация
Регуляция химических реакций осуществляется также за счет пространственного отделения внутриклеточными мембранами
Слайд 45 Изоферменты
Изоферменты - это множественные формы ферментов, которые катализируют одну и ту
Изоферменты
Изоферменты - это множественные формы ферментов, которые катализируют одну и ту

Слайд 46Изменения спектра ЛДГ в онтогенезе
Соотношение разных форм ЛДГ в тканях не является
Изменения спектра ЛДГ в онтогенезе
Соотношение разных форм ЛДГ в тканях не является

 Реакции окисления и восстановления биоорганических соединений. Структура и функции биолекул
Реакции окисления и восстановления биоорганических соединений. Структура и функции биолекул Морфологический критерий вида. Сходство внешнего и внутреннего строения особей
Морфологический критерий вида. Сходство внешнего и внутреннего строения особей Болетус, белый гриб Boletus edulis
Болетус, белый гриб Boletus edulis Бактерии
Бактерии Функция и строение нервной системы
Функция и строение нервной системы Белки. Первичная структура белка
Белки. Первичная структура белка Семейство бобовые
Семейство бобовые Тварини - багатство Землі
Тварини - багатство Землі Мамедова С.А. Мамедова С.А. Учитель биологии МБОУ «Средняя школа №6»
Мамедова С.А. Мамедова С.А. Учитель биологии МБОУ «Средняя школа №6» Презентация на тему Многообразие паукообразных и их значение
Презентация на тему Многообразие паукообразных и их значение  Биосемантика: интерпретационные процессы в живых организмах
Биосемантика: интерпретационные процессы в живых организмах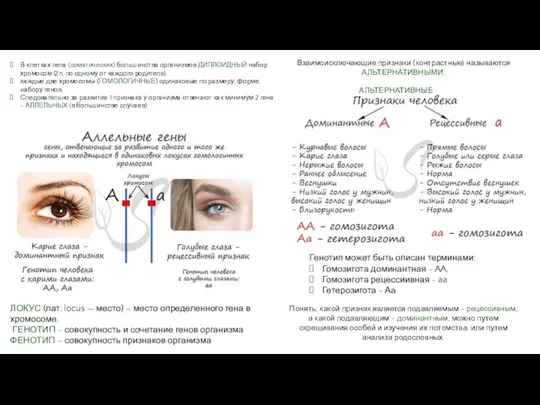 Zakony_Mendelya_analiz_skresch_EGE2
Zakony_Mendelya_analiz_skresch_EGE2 Зимующие птицы зимой
Зимующие птицы зимой Класс птицы. Основные ароморфозы
Класс птицы. Основные ароморфозы З9йн9б Биишева. 1908-1996
З9йн9б Биишева. 1908-1996 Плейотропия
Плейотропия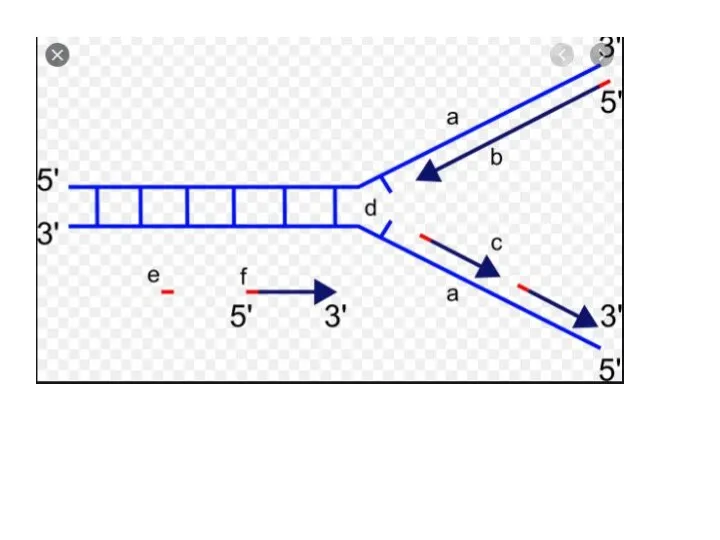 Репликация ДНК
Репликация ДНК Дыхательные движения
Дыхательные движения Селекция пшеницы, ржи и кукурузы
Селекция пшеницы, ржи и кукурузы Критерии и структура вида
Критерии и структура вида e8b9f2d5-8f92-41de-a0fd-b144b6a3fbfd
e8b9f2d5-8f92-41de-a0fd-b144b6a3fbfd Синдром верхнего мотонейрона. Спастичность: терминология, этиология, патогенез
Синдром верхнего мотонейрона. Спастичность: терминология, этиология, патогенез Технология генетического нокаута
Технология генетического нокаута Одежда в животном мире
Одежда в животном мире Технология создания настольного мини-садика в плошке. Ассортимент растений и виды ёмкостей
Технология создания настольного мини-садика в плошке. Ассортимент растений и виды ёмкостей Ткани (1)
Ткани (1) Гормональная регуляция углеводов
Гормональная регуляция углеводов Влияние факторов среды на строение листа. Видоизменения листьев
Влияние факторов среды на строение листа. Видоизменения листьев