- Фермент2

Содержание
- 2. Метаболический путь – это цепь взаимосвязанных реакций, в ходе которой исходный субстрат S превращается в конечный
- 3. Регуляция скорости реакций метаболического пути осуществляется путем регуляции действия определенных ферментов, которые называются регуляторными (как правило,
- 4. Очень часто ингибитором регуляторного фермента является конечный продукт метаболического пути. Фермент, катализирующий превращение субстрата А в
- 5. Регуляция активности ферментов Основные способы регуляции активности ферментов: - аллостерическая регуляция; - регуляция с помощью белок-белковых
- 6. Аллостерическая регуляция Аллостерические ферменты: 1)Олигомерные белки, состоящие из 2-х или более субъединиц – регуляторной (R) и
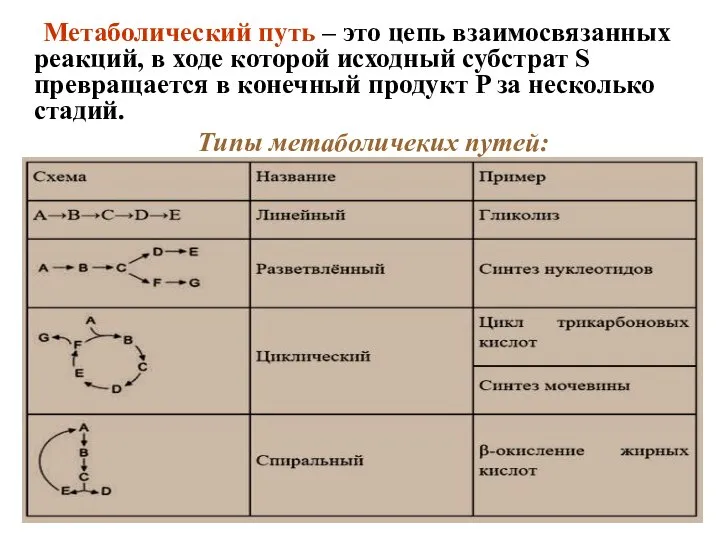
- 7. Аллостерический центр Активный центр Эффекторы делятся на: активаторы (A) и ингибиторы (I). Они влияют на сродство
- 8. Схема аллостерической регуляции
- 9. Частичный протеолиз - активация фермента через отщепление от него одного или нескольких пептидных фрагментов. Происходит изменение
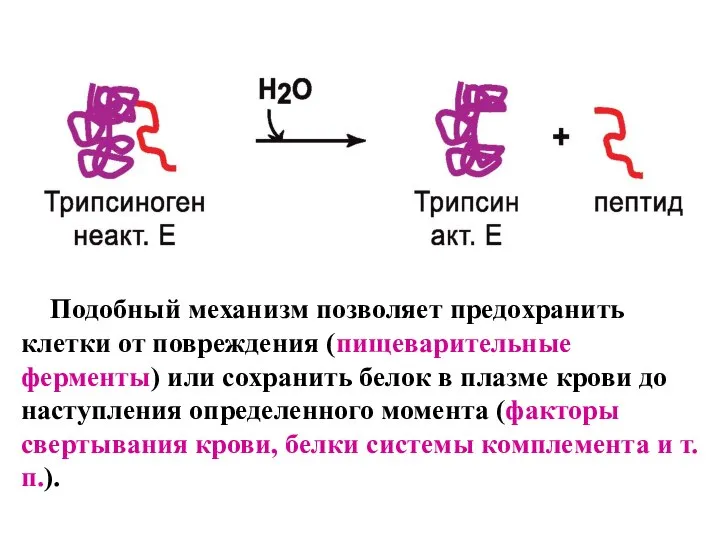
- 10. Подобный механизм позволяет предохранить клетки от повреждения (пищеварительные ферменты) или сохранить белок в плазме крови до
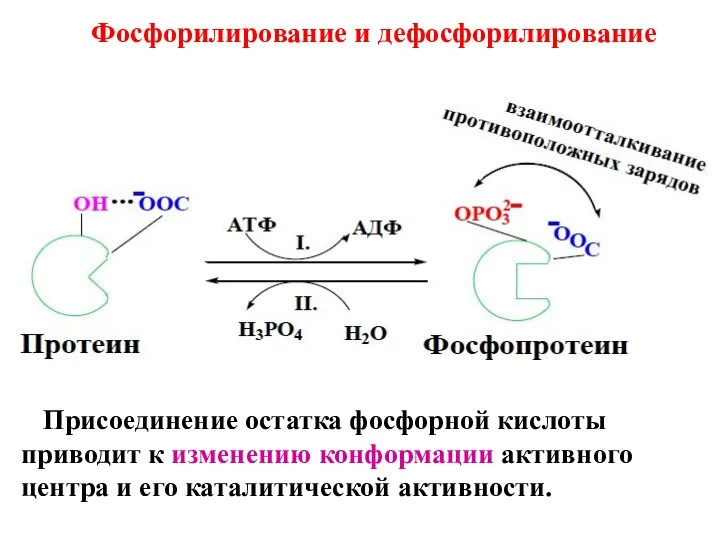
- 11. Фосфорилирование и дефосфорилирование Присоединение остатка фосфорной кислоты приводит к изменению конформации активного центра и его каталитической
- 12. I. Фосфорилирование – катализируют ферменты: протеинкиназы (A, C, G и др.); II. Дефосфорилирование – катализируют ферменты:
- 13. Белок-белковые взаимодействия Термин обозначает ситуацию, в качестве регулятора выступают не метаболиты биохимических процессов, а специфичные белки.
- 14. 1.Мембранный фермент аденилатциклаза является чувствительным к воздействию мембранного G-белка, который сам активируется при действии на клетку
- 15. 2. Регуляция активности протеинкиназы А через механизм ассоциации-диссоциации: Протеинкиназа А является тетрамерным ферментом (2С и 2R).
- 16. Инактивация протеинкиназы A Фермент фосфодиэстераза катализирует гидролиз цАМФ до АМФ, что приводит к сборке тетрамерного комплекса
- 17. Ингибиторы фосфодиэстеразы используются: а) при острой сердечной недостаточности как кардиотонические средства; б) для лечения бронхиальной астмы.
- 18. Ингибиторы ферментов Ингибиторы – это вещества полностью или частично подавляющие активность ферментов. Ингибиторы способны взаимодействовать с
- 19. Обратимые ингибиторы присоединяются к ферменту слабыми связями и могут легко отделяться от фермента. Обратимые конкурентные ингибиторы:
- 20. Конкурентный тип ингибирования
- 22. Кинетические зависимости При обратимом конкурентном ингибировании возрастает Kм, но, как правило, не меняется максимальная скорость реакции.
- 23. Лекарственные препараты как конкурентные ингибиторы Прозерин, эндрофоний, дистигмин, галантамин – ингибируют ацетилхолинэстеразу, фермент, разрушающий ацетилхолин, что
- 24. Обратимые неконкурентные ингибиторы: не являются структурными аналогами субстрата; не присоединяются к активному центру фермента; действуют на
- 25. Степень торможения активности фермента неконкурентными ингибиторами зависит от продолжительности их действия и их концентрации. При снижении
- 26. Неконкурентный тип ингибирования
- 27. Кинетические зависимости При обратимом неконкурентном ингибировании уменьшается Vmax, но, как правило, не меняется Км. Vmax уменьшается,
- 28. Необратимые ингибиторы характеризуются тем, что связываются с молекулой фермента прочными ковалентными связями (чаще всего модификации подвергается
- 29. Необратимые специфические ингибиторы – присоединяются к строго определённой группе в области активного центра E, образуя с
- 30. ДФФ ингибирует АХЭ, которая участвует в передаче нервных импульсов от нейрона к нейрону. Ингибирование этого фермента
- 31. Неспецифические ингибиторы образуют прочную ковалентную связь с группами, расположенными в любом участке фермента (довольно часто: вне
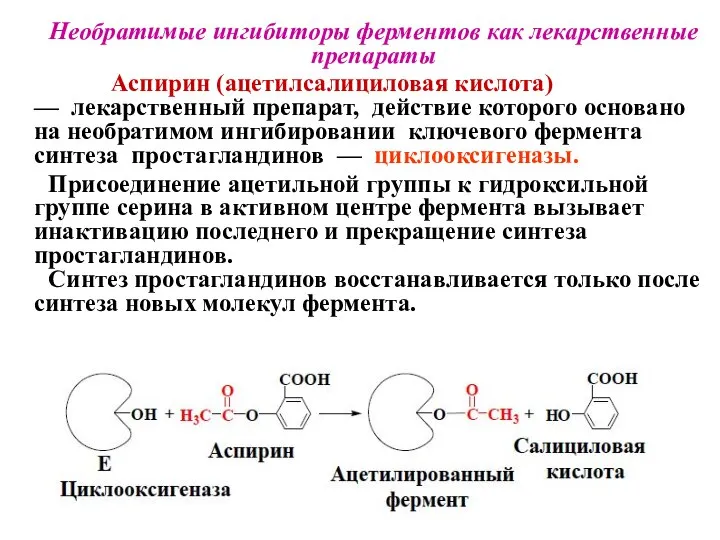
- 32. Необратимые ингибиторы ферментов как лекарственные препараты Аспирин (ацетилсалициловая кислота) — лекарственный препарат, действие которого основано на
- 33. Изоферменты – это молекулярные формы одного и того же фермента, возникшие в результате небольших генетических различий
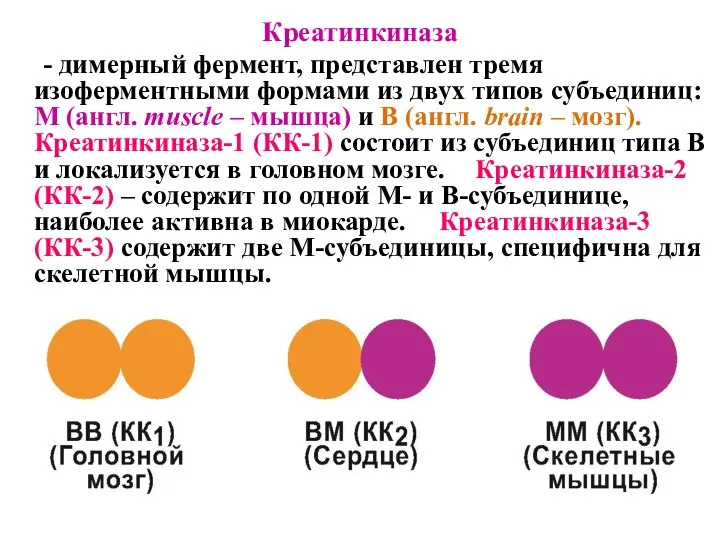
- 34. Креатинкиназа - димерный фермент, представлен тремя изоферментными формами из двух типов субъединиц: M (англ. muscle –
- 36. Скачать презентацию
Слайд 3 Регуляция скорости реакций метаболического пути осуществляется путем регуляции действия определенных ферментов,
Регуляция скорости реакций метаболического пути осуществляется путем регуляции действия определенных ферментов,

Слайд 4 Очень часто ингибитором регуляторного фермента является конечный продукт метаболического пути.
Фермент, катализирующий
Очень часто ингибитором регуляторного фермента является конечный продукт метаболического пути.
Фермент, катализирующий

Слайд 5 Регуляция активности ферментов
Основные способы регуляции активности ферментов:
- аллостерическая регуляция;
- регуляция с помощью
Регуляция активности ферментов
Основные способы регуляции активности ферментов:
- аллостерическая регуляция;
- регуляция с помощью

Слайд 6 Аллостерическая регуляция
Аллостерические ферменты:
1)Олигомерные белки, состоящие из 2-х или более субъединиц –
Аллостерическая регуляция
Аллостерические ферменты:
1)Олигомерные белки, состоящие из 2-х или более субъединиц –

Слайд 7 Аллостерический центр Активный центр
Эффекторы делятся на:
активаторы (A) и ингибиторы
Аллостерический центр Активный центр
Эффекторы делятся на:
активаторы (A) и ингибиторы
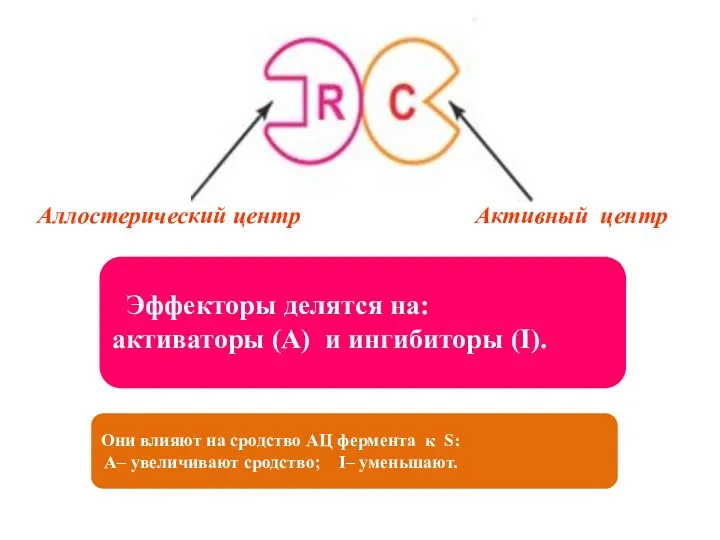
Слайд 8Схема аллостерической регуляции
Схема аллостерической регуляции
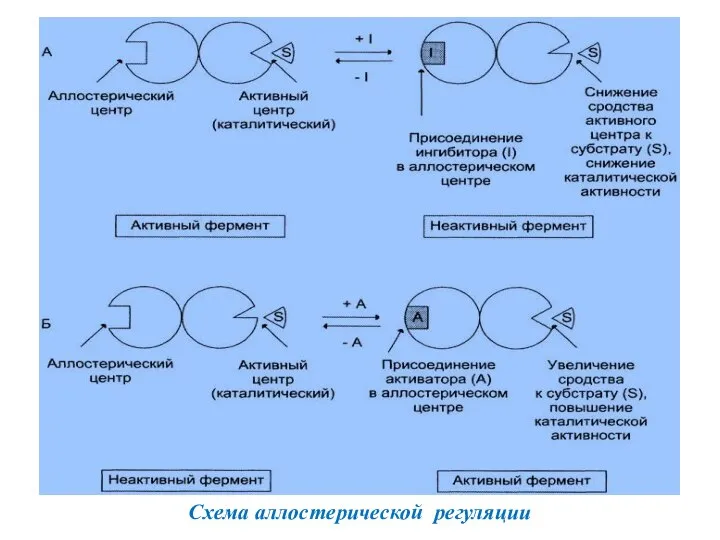
Слайд 9Частичный протеолиз
- активация фермента через отщепление от него одного или нескольких пептидных
Частичный протеолиз
- активация фермента через отщепление от него одного или нескольких пептидных
Слайд 10 Подобный механизм позволяет предохранить клетки от повреждения (пищеварительные ферменты) или сохранить
Подобный механизм позволяет предохранить клетки от повреждения (пищеварительные ферменты) или сохранить
Слайд 11Фосфорилирование и дефосфорилирование
Присоединение остатка фосфорной кислоты приводит к изменению конформации активного
Фосфорилирование и дефосфорилирование
Присоединение остатка фосфорной кислоты приводит к изменению конформации активного
Слайд 12I. Фосфорилирование – катализируют ферменты: протеинкиназы (A, C, G и др.);
II.
I. Фосфорилирование – катализируют ферменты: протеинкиназы (A, C, G и др.);
II.

Слайд 13 Белок-белковые взаимодействия
Термин обозначает ситуацию, в качестве регулятора выступают не метаболиты
Белок-белковые взаимодействия
Термин обозначает ситуацию, в качестве регулятора выступают не метаболиты

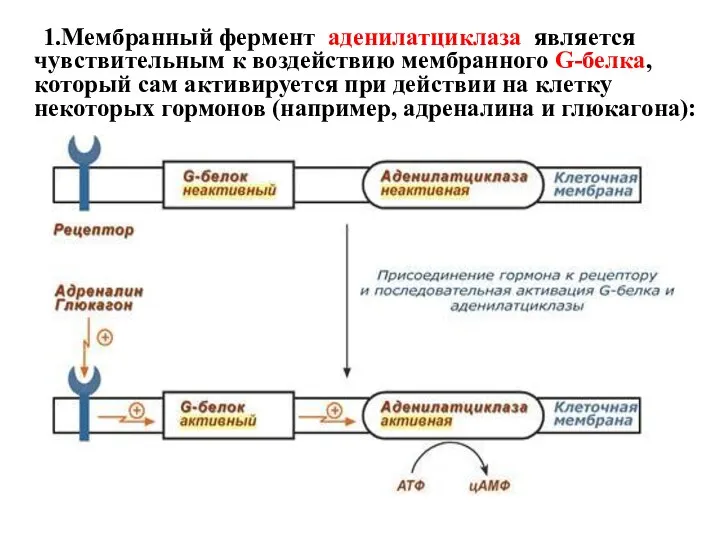
Слайд 14 1.Мембранный фермент аденилатциклаза является чувствительным к воздействию мембранного G-белка, который сам активируется при
1.Мембранный фермент аденилатциклаза является чувствительным к воздействию мембранного G-белка, который сам активируется при
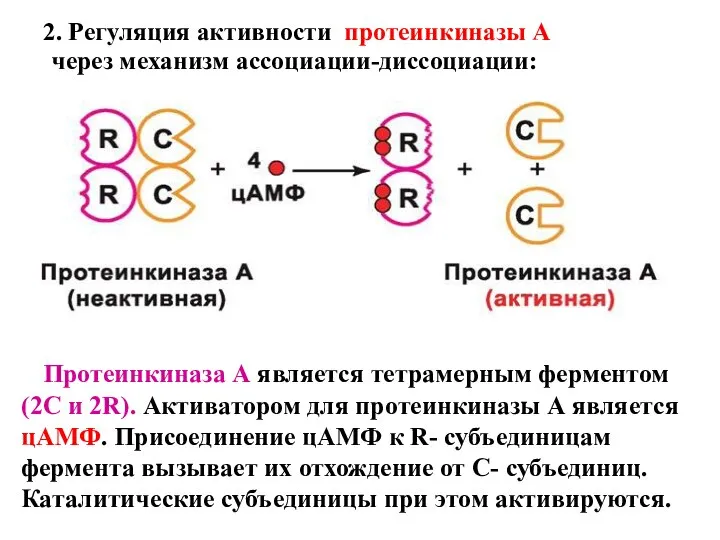
Слайд 15 2. Регуляция активности протеинкиназы А
через механизм ассоциации-диссоциации:
Протеинкиназа А является тетрамерным ферментом
2. Регуляция активности протеинкиназы А
через механизм ассоциации-диссоциации:
Протеинкиназа А является тетрамерным ферментом
Слайд 16Инактивация протеинкиназы A
Фермент фосфодиэстераза катализирует гидролиз цАМФ до АМФ, что приводит
Инактивация протеинкиназы A
Фермент фосфодиэстераза катализирует гидролиз цАМФ до АМФ, что приводит
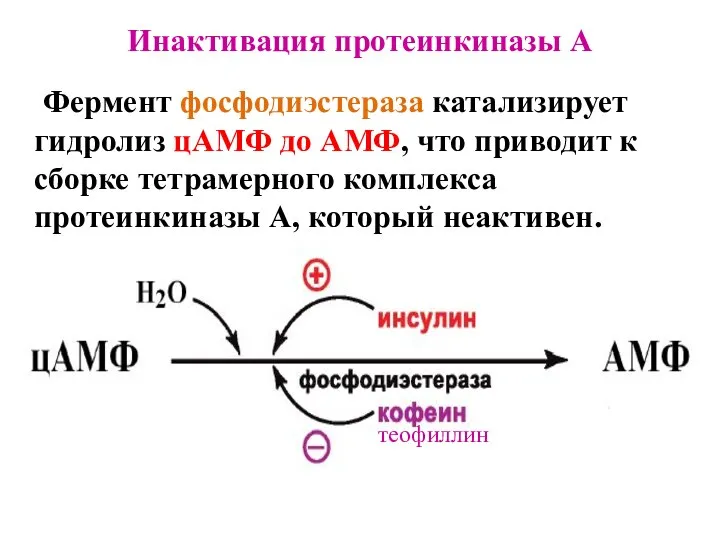
Слайд 17 Ингибиторы фосфодиэстеразы используются:
а) при острой сердечной недостаточности как кардиотонические средства;
б) для
Ингибиторы фосфодиэстеразы используются:
а) при острой сердечной недостаточности как кардиотонические средства;
б) для

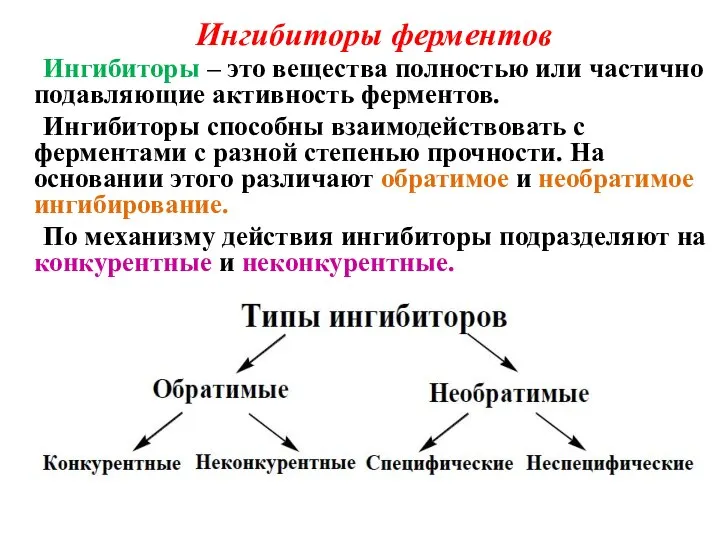
Слайд 18Ингибиторы ферментов
Ингибиторы – это вещества полностью или частично подавляющие активность ферментов.
Ингибиторы
Ингибиторы ферментов
Ингибиторы – это вещества полностью или частично подавляющие активность ферментов.
Ингибиторы
Слайд 19 Обратимые ингибиторы
присоединяются к ферменту слабыми связями и могут легко
Обратимые ингибиторы
присоединяются к ферменту слабыми связями и могут легко

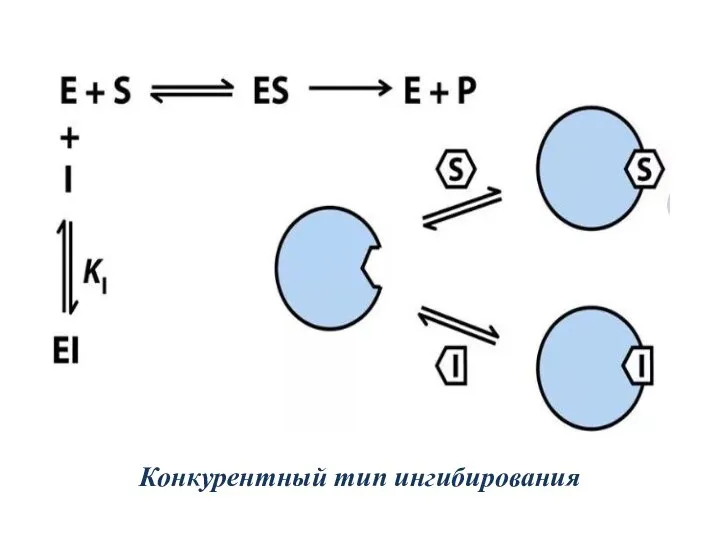
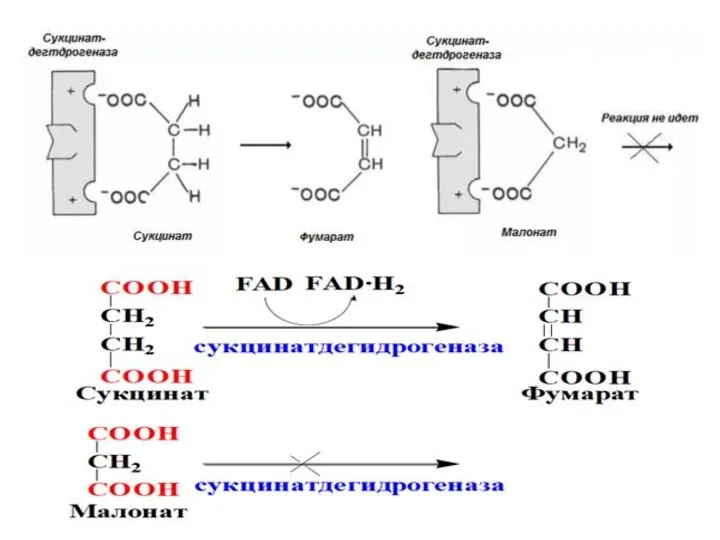
Слайд 20Конкурентный тип ингибирования
Конкурентный тип ингибирования
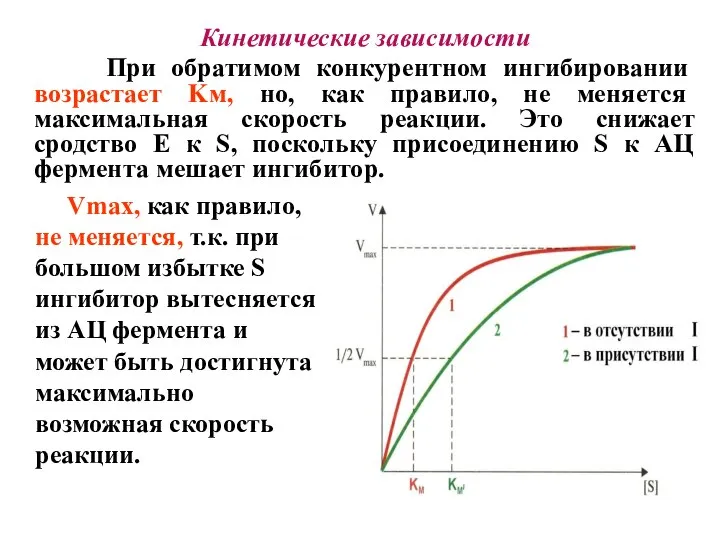
Слайд 22 Кинетические зависимости
При обратимом конкурентном ингибировании возрастает Kм, но, как правило, не
Кинетические зависимости
При обратимом конкурентном ингибировании возрастает Kм, но, как правило, не
Слайд 23Лекарственные препараты как конкурентные ингибиторы
Прозерин, эндрофоний, дистигмин, галантамин – ингибируют ацетилхолинэстеразу, фермент,
Лекарственные препараты как конкурентные ингибиторы
Прозерин, эндрофоний, дистигмин, галантамин – ингибируют ацетилхолинэстеразу, фермент,

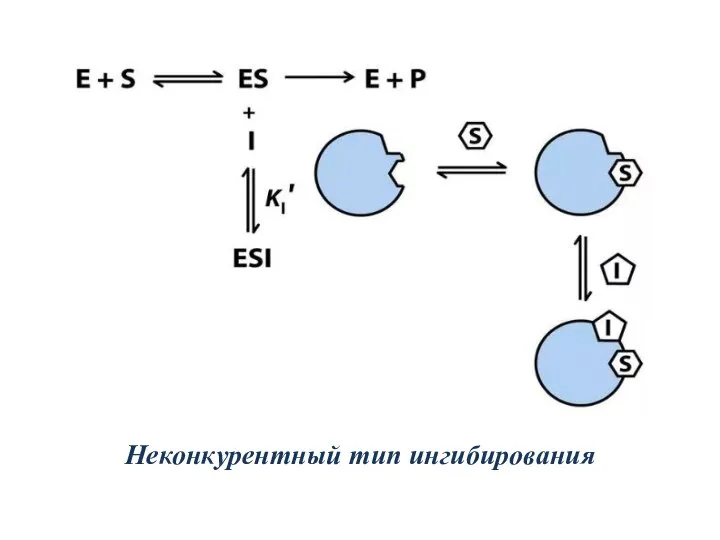
Слайд 24Обратимые неконкурентные ингибиторы:
не являются структурными аналогами субстрата;
не присоединяются к активному
не являются структурными аналогами субстрата;
не присоединяются к активному

Слайд 25 Степень торможения активности фермента неконкурентными ингибиторами зависит от продолжительности их действия и
Степень торможения активности фермента неконкурентными ингибиторами зависит от продолжительности их действия и

Слайд 26Неконкурентный тип ингибирования
Неконкурентный тип ингибирования
Слайд 27Кинетические зависимости
При обратимом неконкурентном ингибировании уменьшается Vmax, но, как правило, не
Кинетические зависимости
При обратимом неконкурентном ингибировании уменьшается Vmax, но, как правило, не

Слайд 28 Необратимые ингибиторы
характеризуются тем, что связываются с молекулой фермента прочными ковалентными
Необратимые ингибиторы
характеризуются тем, что связываются с молекулой фермента прочными ковалентными

Слайд 29 Необратимые специфические ингибиторы
– присоединяются к строго определённой группе в области активного центра
Необратимые специфические ингибиторы
– присоединяются к строго определённой группе в области активного центра

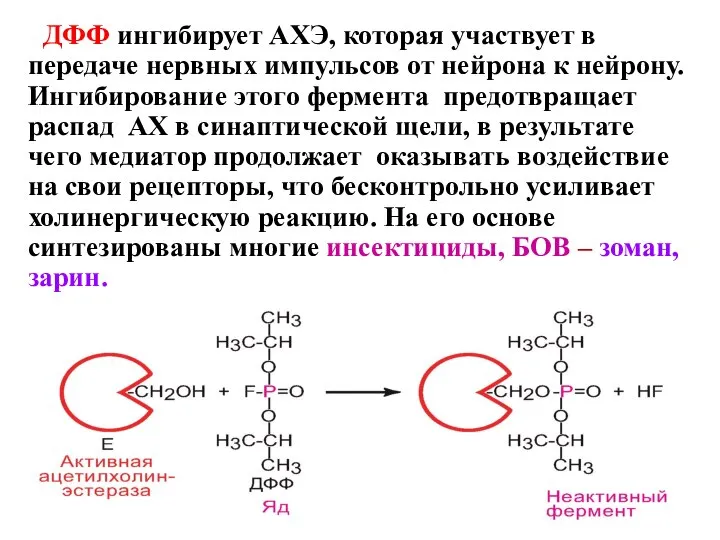
Слайд 30 ДФФ ингибирует АХЭ, которая участвует в передаче нервных импульсов от нейрона к
ДФФ ингибирует АХЭ, которая участвует в передаче нервных импульсов от нейрона к
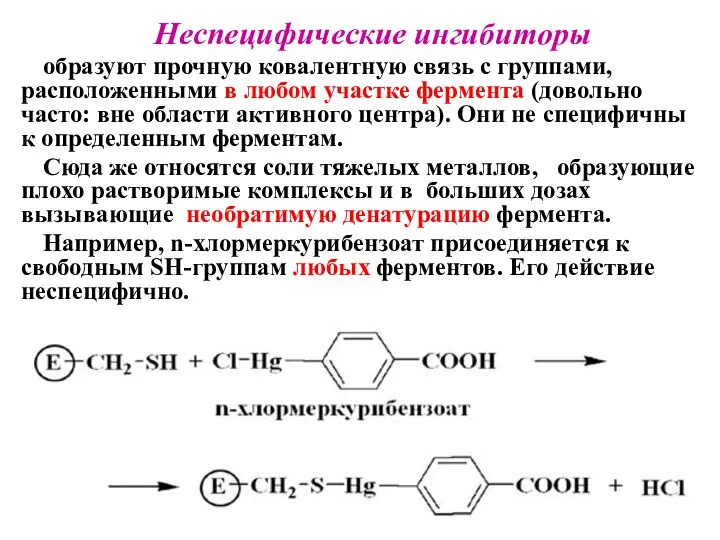
Слайд 31 Неспецифические ингибиторы
образуют прочную ковалентную связь с группами, расположенными в любом участке
Неспецифические ингибиторы
образуют прочную ковалентную связь с группами, расположенными в любом участке
Слайд 32 Необратимые ингибиторы ферментов как лекарственные препараты
Аспирин (ацетилсалициловая кислота) — лекарственный препарат,
Необратимые ингибиторы ферментов как лекарственные препараты
Аспирин (ацетилсалициловая кислота) — лекарственный препарат,
Слайд 33 Изоферменты – это молекулярные формы одного и того же фермента, возникшие
Изоферменты – это молекулярные формы одного и того же фермента, возникшие

Слайд 34Креатинкиназа
- димерный фермент, представлен тремя изоферментными формами из двух типов субъединиц:
Креатинкиназа
- димерный фермент, представлен тремя изоферментными формами из двух типов субъединиц:
 Происхождение жизни на Земле
Происхождение жизни на Земле Сравнение свойств организма человека и животных
Сравнение свойств организма человека и животных Кристаллин. Локализация. Функции
Кристаллин. Локализация. Функции Презентация на тему Физические явления в живых организмах
Презентация на тему Физические явления в живых организмах  Неполное доминирование у растений и животных
Неполное доминирование у растений и животных Взаимодействие генов
Взаимодействие генов Функции корней
Функции корней Презентация на тему Многообразие млекопитающих
Презентация на тему Многообразие млекопитающих  Презентация на тему Строение спинного мозга
Презентация на тему Строение спинного мозга  Путешествие вглубь листа
Путешествие вглубь листа Растения Самбурга
Растения Самбурга Весенний букет цветов
Весенний букет цветов Строение корня и побега растений
Строение корня и побега растений Охота в Рязанских лесах
Охота в Рязанских лесах Обмен серосодержащих аминокислот
Обмен серосодержащих аминокислот Грибы Саратовской области
Грибы Саратовской области Биология и история
Биология и история Понятия проницаемости и селективности. Типы мембранного транспорта
Понятия проницаемости и селективности. Типы мембранного транспорта Липиды. Углеводы_, 9-10 класс
Липиды. Углеводы_, 9-10 класс Мышцы головы животного
Мышцы головы животного Царство бактерии
Царство бактерии Среда обитания организмов
Среда обитания организмов Презентация на тему Яблочный спас
Презентация на тему Яблочный спас  Срок жизни растений
Срок жизни растений Сравнение свойств организма человека и животных
Сравнение свойств организма человека и животных Микроскопический метод исследования. Морфология микроорганизмов. Занятие №4
Микроскопический метод исследования. Морфология микроорганизмов. Занятие №4 Фикус в нашем доме
Фикус в нашем доме Классификация в психиатрии. Тревожные расстройства. Нейропсихиатрия и биология зависимости. Занятие 2
Классификация в психиатрии. Тревожные расстройства. Нейропсихиатрия и биология зависимости. Занятие 2