- Ферменты. Классификация фермента (шифр)

Содержание
- 2. ФЕРМЕНТЫ – это биологические катализаторы белковой природы, ускоряющие все химические реакции, протекающие в живом организме. В
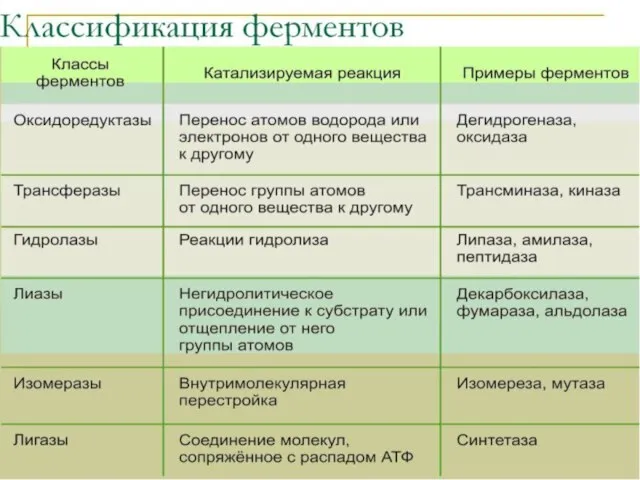
- 6. Классификация фермента (шифр)
- 9. Оксидоредуктазы Ферменты этого класса катализируют окислительно-восстановительные реакции, лежащие в основе биологического окисления. Класс насчитывает 22 подкласса.
- 10. Пример
- 11. Трансферазы Коферментами являются пиридоксальфосфат, коэнзим А, тетрагидрофолиевая кислота, метилкобаламин. Класс подразделяется на 9 подклассов в зависимости
- 13. Гидролазы Гидролазы – ферменты, осуществляющие разрыв внутримолекулярных связей в субстрате путем присоединения элементов Н2О, подразделяются на
- 15. Лиазы Лиазы являются сложными ферментами. Коферментами служат пиридоксальфосфат, тиаминдифосфат, участвует магний, кобальт. Ферменты делятся на подклассы
- 18. Изомеразы ферменты, катализирующие изомерные превращения в пределах одной молекулы. Изомеразы – сложные ферменты. К их коферментам
- 19. Лигазы ферменты, катализирующие присоединение друг к другу двух молекул с использованием энергии высокоэнергетических связей АТФ (или
- 20. Глутаминсинтетаза
- 27. Механизм действия ферментов состоит в образовании комплекса между ферментом и субстратом за счет образования нековалентных связей
- 30. Активный центр фермента Активный центр – это часть молекулы фермента, на которой происходит связывание и превращение
- 39. Свойства ферментов Ферменты обладают всеми свойствами белков. Также обладают уникальными свойствами. СПЕЦИФИЧНОСТЬ ТЕРМОЛАБИЛЬНОСТЬ рН-ЧУВСТВИТЕЛЬНОСТЬ
- 40. Специфичность А. Субстратная Б. Специфичность действия
- 42. Термолабильность – это неустойчивость ферментов к действию высоких температур. Это свойство объясняется денатурацией фермента при температурах
- 43. Линейное увеличение скорости ферментативной реакции от температуры наблюдается в области 5-25 о С. При дальнейшем повышении
- 45. Ферменты проявляют наибольшую активность при разном рН среды, например пепсин – при рН 1,5, липаза –
- 46. рН, при котором наблюдается максимальная активность фермента, называется рН-оптимумом. Он зависит от степени ионизации функциональных полярных
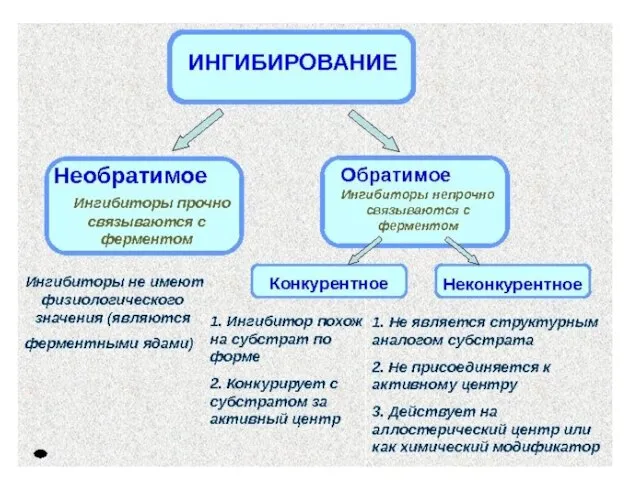
- 47. АКТИВАЦИЯ И ИНГИБИРОВАНИЕ АКТИВНОСТИ ФЕРМЕНТОВ
- 48. Активаторы ферментов Вещества, повышающие активность ферментов. Активаторами могут быть: ионы металлов: натрия, магния, калия, кальция, цинка
- 54. Кинетика ферментативной реакции Активность фермента – количество фермента, катализирующее превращение 1 моля субстрата за 1 сек.
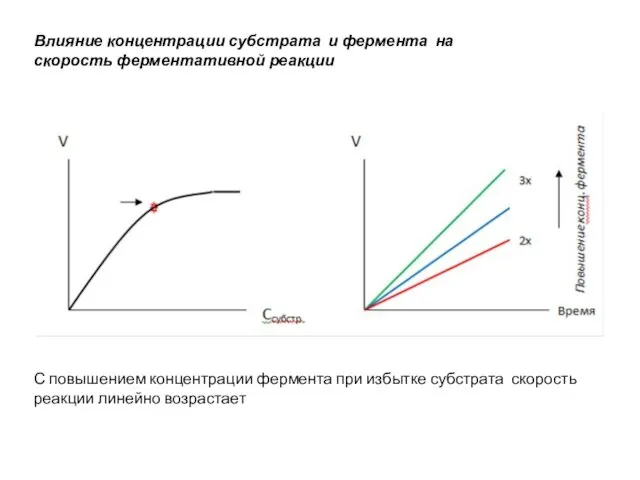
- 55. ВЛИЯНИЕ КОНЦЕНТРАЦИИ ФЕРМЕНТА И СУБСТРАТА НА СКОРОСТЬ ФЕРМЕНТАТИВНОЙ РЕАКЦИИ При постоянной концентрации фермента скорость реакции при
- 56. С повышением концентрации фермента при избытке субстрата скорость реакции линейно возрастает Влияние концентрации субстрата и фермента
- 57. Количественное соотношение между концентрацией субстрата и скоростью ферментативной реакции выражается уравнением Михаэлиса-Ментен
- 60. Скачать презентацию
Слайд 2ФЕРМЕНТЫ – это биологические катализаторы белковой природы, ускоряющие все химические реакции, протекающие
ФЕРМЕНТЫ – это биологические катализаторы белковой природы, ускоряющие все химические реакции, протекающие

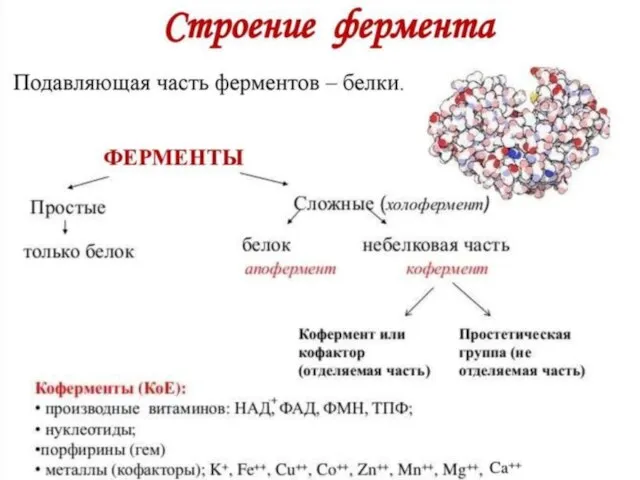


Слайд 6Классификация фермента (шифр)
Классификация фермента (шифр)


Слайд 9Оксидоредуктазы
Ферменты этого класса катализируют окислительно-восстановительные реакции, лежащие в основе биологического окисления. Класс насчитывает 22 подкласса.
Оксидоредуктазы
Ферменты этого класса катализируют окислительно-восстановительные реакции, лежащие в основе биологического окисления. Класс насчитывает 22 подкласса.

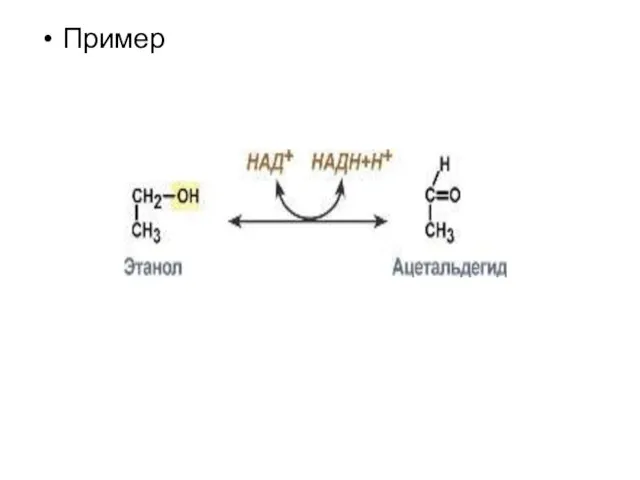
Слайд 10Пример
Пример
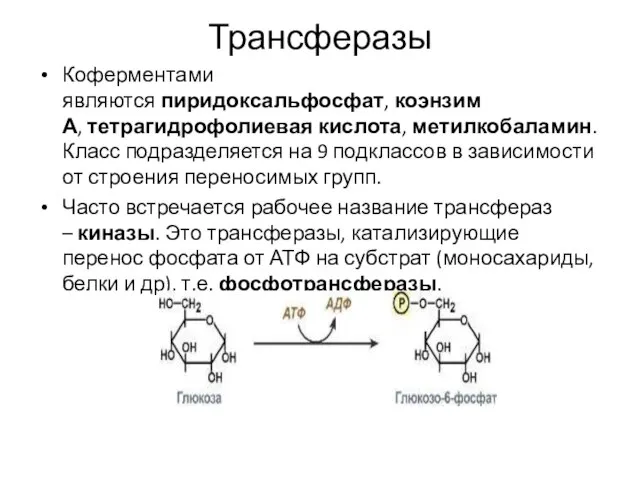
Слайд 11Трансферазы
Коферментами являются пиридоксальфосфат, коэнзим А, тетрагидрофолиевая кислота, метилкобаламин. Класс подразделяется на 9 подклассов в зависимости от
Трансферазы
Коферментами являются пиридоксальфосфат, коэнзим А, тетрагидрофолиевая кислота, метилкобаламин. Класс подразделяется на 9 подклассов в зависимости от

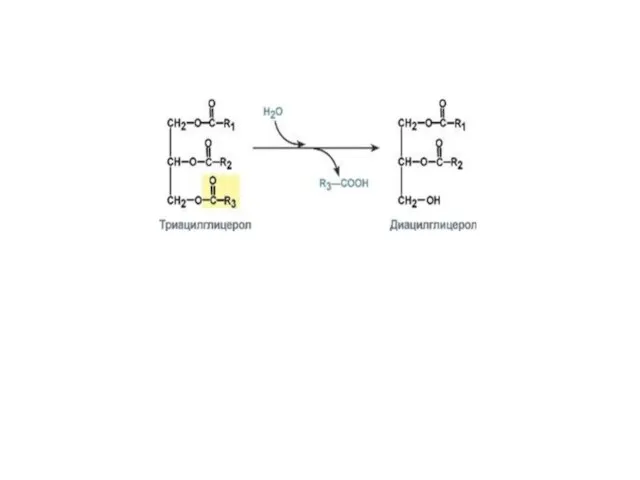
Слайд 13Гидролазы
Гидролазы – ферменты, осуществляющие разрыв внутримолекулярных связей в субстрате путем присоединения элементов
Гидролазы
Гидролазы – ферменты, осуществляющие разрыв внутримолекулярных связей в субстрате путем присоединения элементов

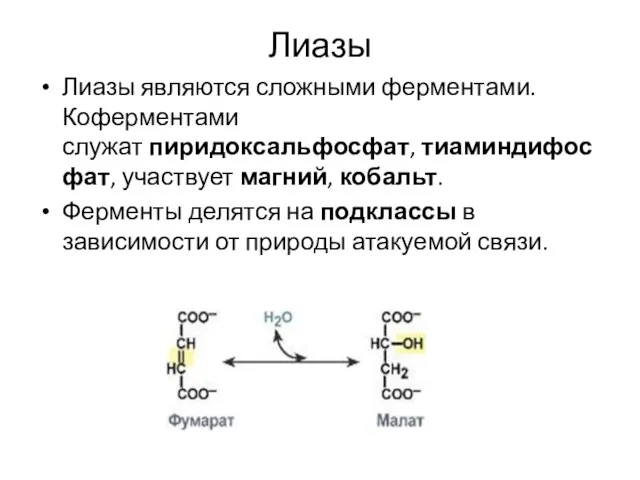
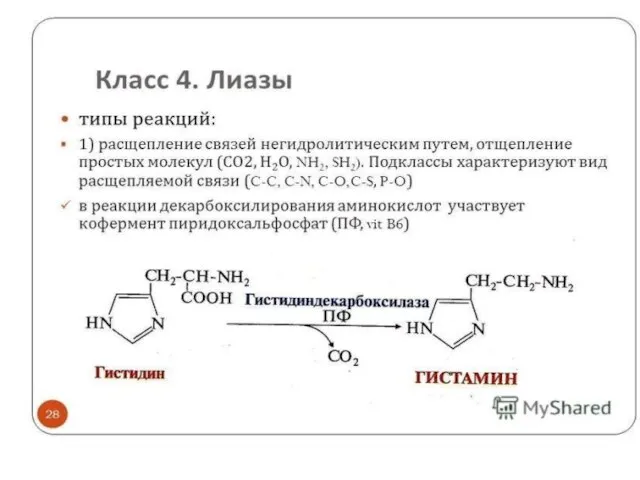
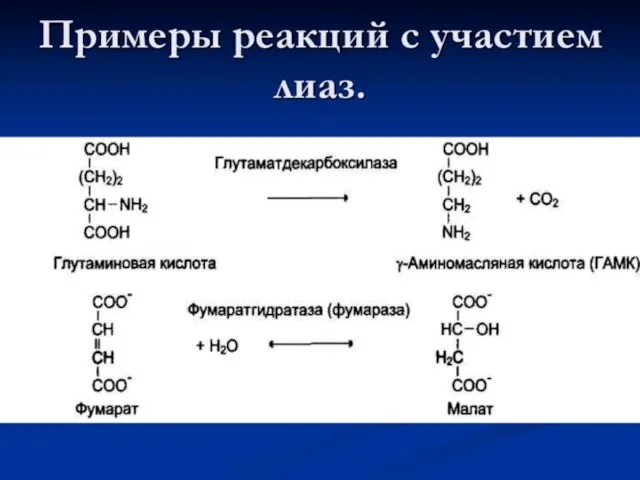
Слайд 15Лиазы
Лиазы являются сложными ферментами. Коферментами служат пиридоксальфосфат, тиаминдифосфат, участвует магний, кобальт.
Ферменты делятся на подклассы в зависимости от природы
Лиазы
Лиазы являются сложными ферментами. Коферментами служат пиридоксальфосфат, тиаминдифосфат, участвует магний, кобальт.
Ферменты делятся на подклассы в зависимости от природы
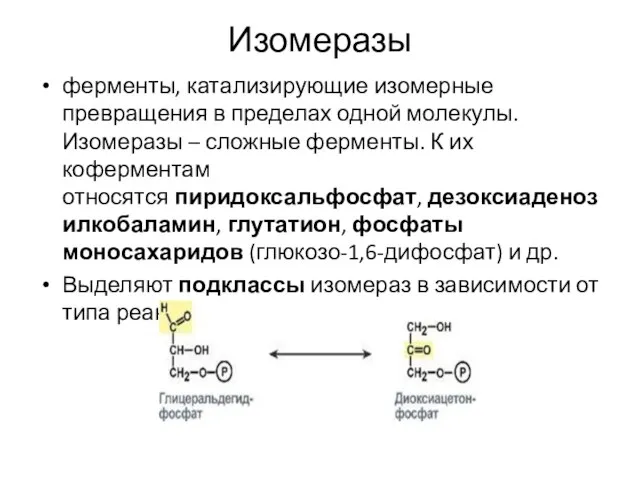
Слайд 18Изомеразы
ферменты, катализирующие изомерные превращения в пределах одной молекулы. Изомеразы – сложные ферменты.
Изомеразы
ферменты, катализирующие изомерные превращения в пределах одной молекулы. Изомеразы – сложные ферменты.
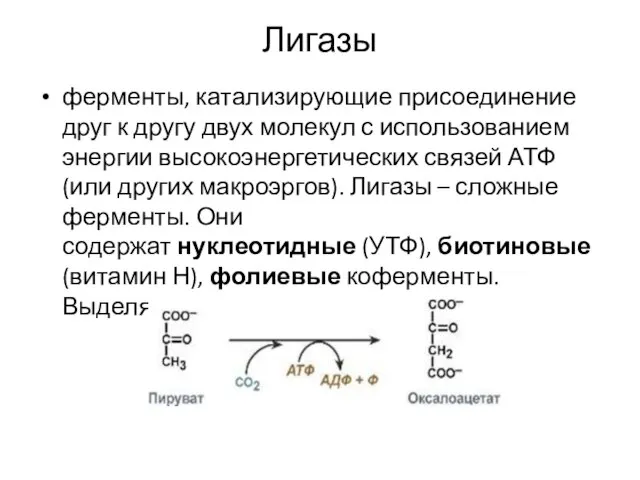
Слайд 19Лигазы
ферменты, катализирующие присоединение друг к другу двух молекул с использованием энергии высокоэнергетических
Лигазы
ферменты, катализирующие присоединение друг к другу двух молекул с использованием энергии высокоэнергетических
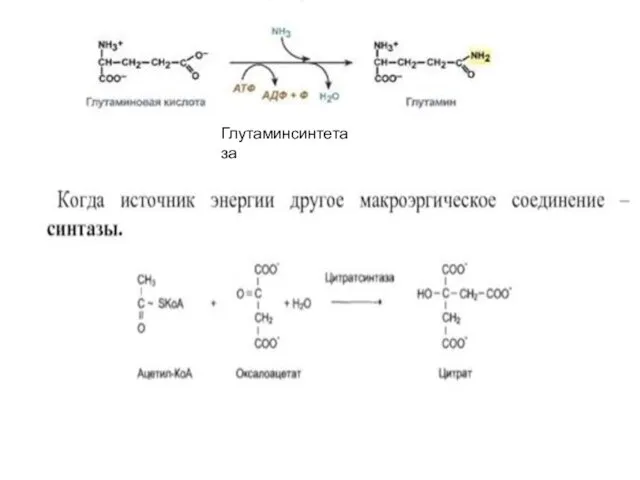
Слайд 20Глутаминсинтетаза
Глутаминсинтетаза

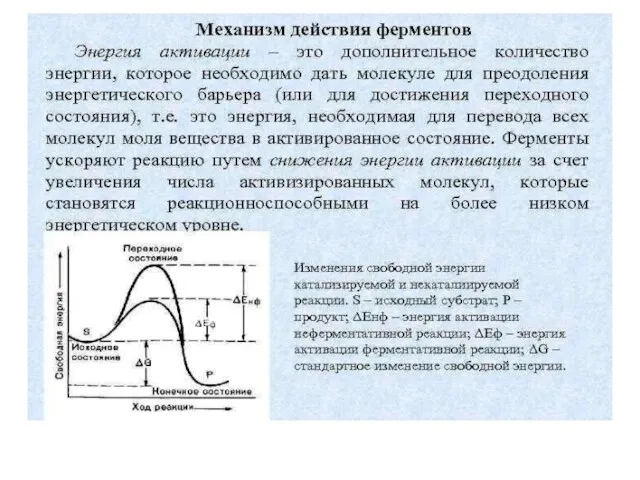
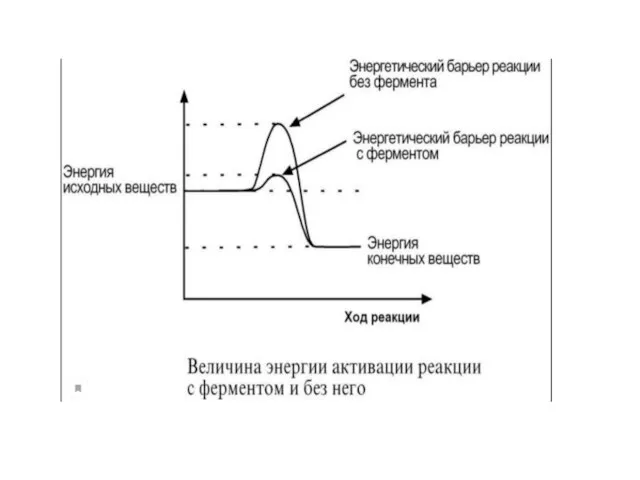

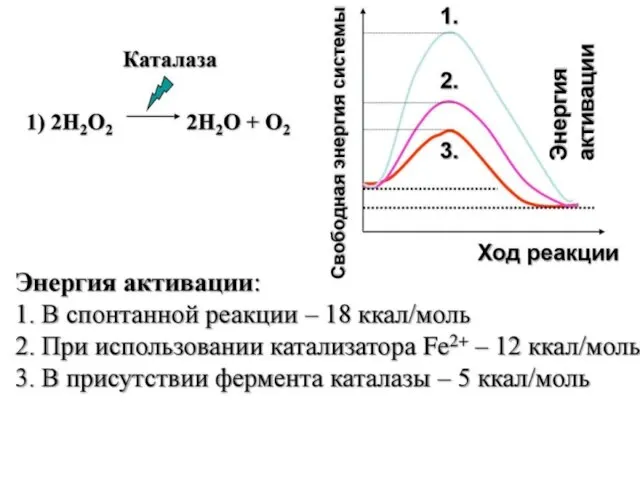

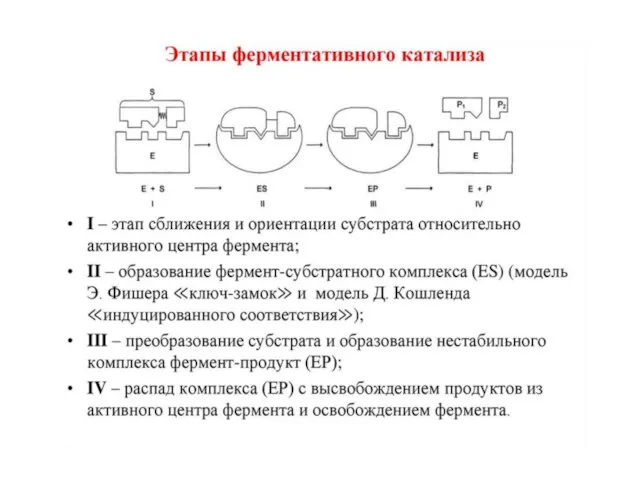
Слайд 27Механизм действия ферментов состоит в образовании комплекса между ферментом и субстратом за
Механизм действия ферментов состоит в образовании комплекса между ферментом и субстратом за

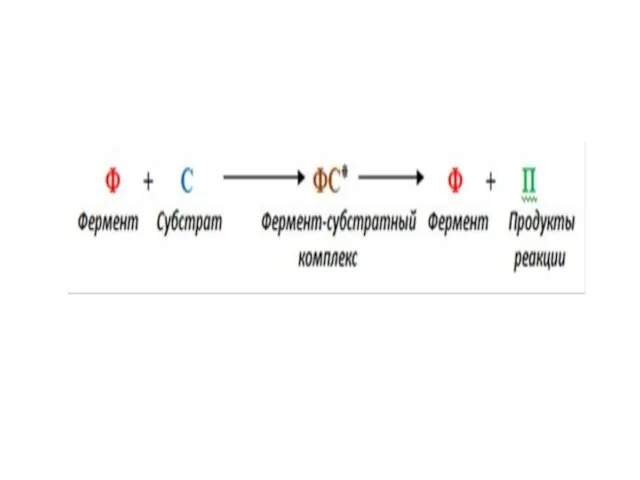
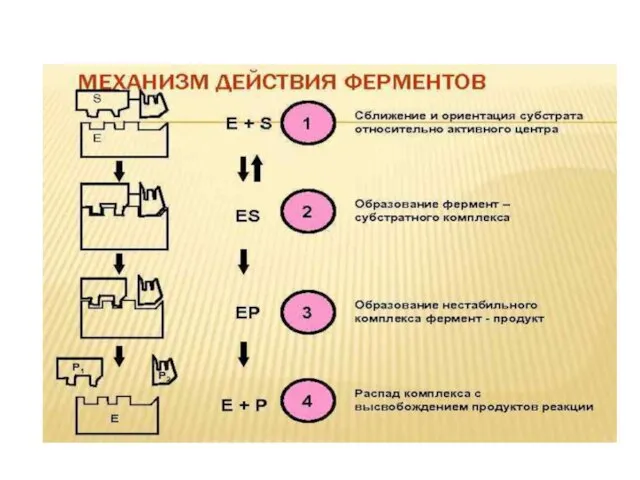
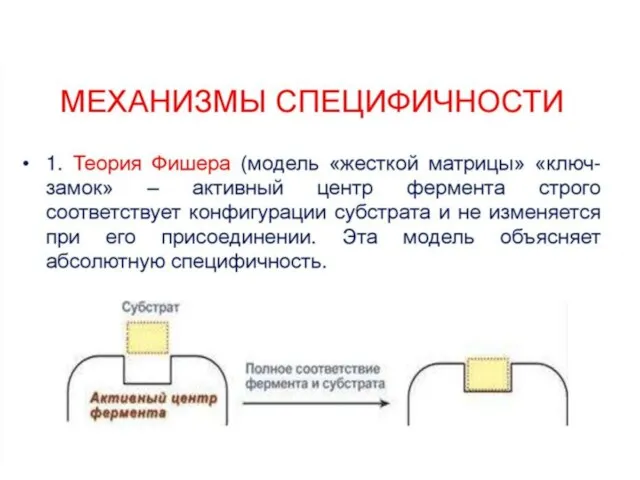
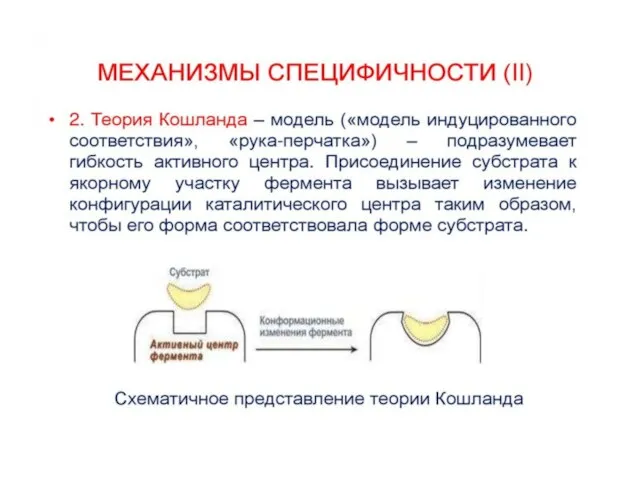
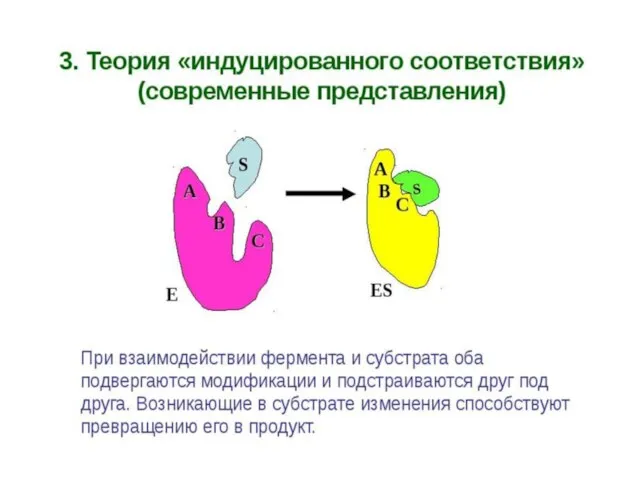
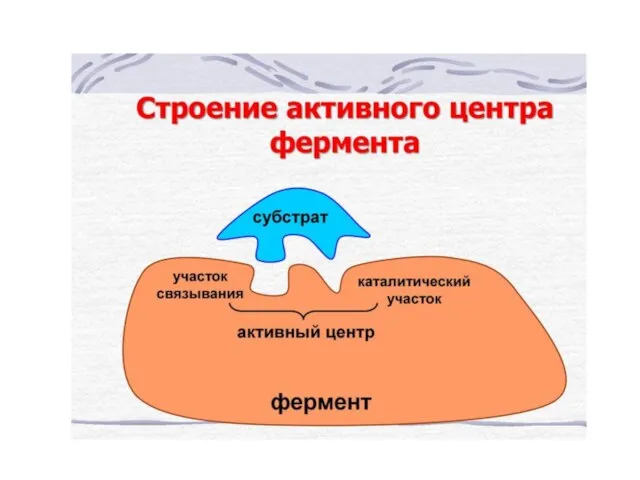
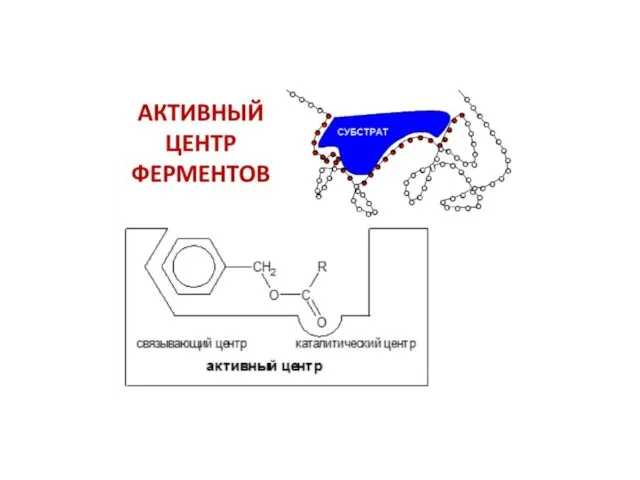
Слайд 30Активный центр фермента
Активный центр – это часть молекулы фермента, на которой происходит
Активный центр фермента
Активный центр – это часть молекулы фермента, на которой происходит


Слайд 39Свойства ферментов
Ферменты обладают всеми свойствами белков.
Также обладают уникальными свойствами.
СПЕЦИФИЧНОСТЬ
ТЕРМОЛАБИЛЬНОСТЬ
рН-ЧУВСТВИТЕЛЬНОСТЬ
Свойства ферментов
Ферменты обладают всеми свойствами белков.
Также обладают уникальными свойствами.
СПЕЦИФИЧНОСТЬ
ТЕРМОЛАБИЛЬНОСТЬ
рН-ЧУВСТВИТЕЛЬНОСТЬ

Слайд 40Специфичность
А. Субстратная
Б. Специфичность действия
Специфичность
А. Субстратная
Б. Специфичность действия


Слайд 42Термолабильность – это неустойчивость ферментов к действию высоких температур. Это свойство объясняется
Термолабильность – это неустойчивость ферментов к действию высоких температур. Это свойство объясняется
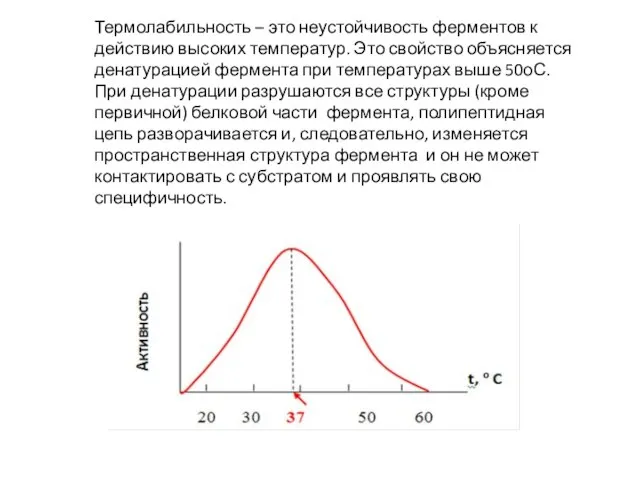
Слайд 43Линейное увеличение скорости ферментативной реакции от температуры наблюдается в области 5-25 о
Линейное увеличение скорости ферментативной реакции от температуры наблюдается в области 5-25 о


Слайд 45Ферменты проявляют наибольшую активность при разном рН среды, например пепсин – при
Ферменты проявляют наибольшую активность при разном рН среды, например пепсин – при
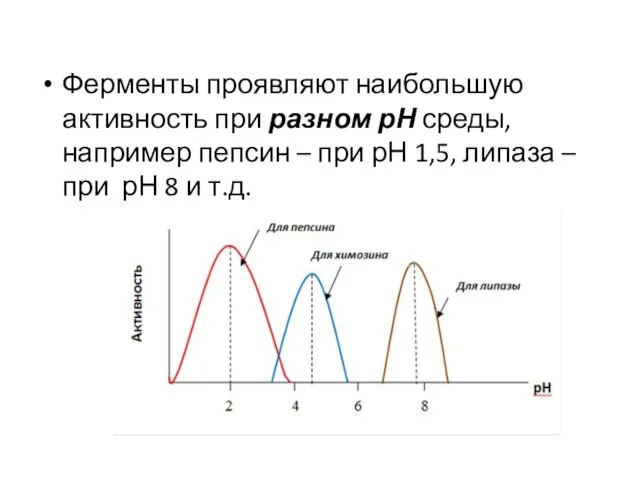
Слайд 46рН, при котором наблюдается максимальная активность фермента, называется рН-оптимумом. Он зависит от
рН, при котором наблюдается максимальная активность фермента, называется рН-оптимумом. Он зависит от

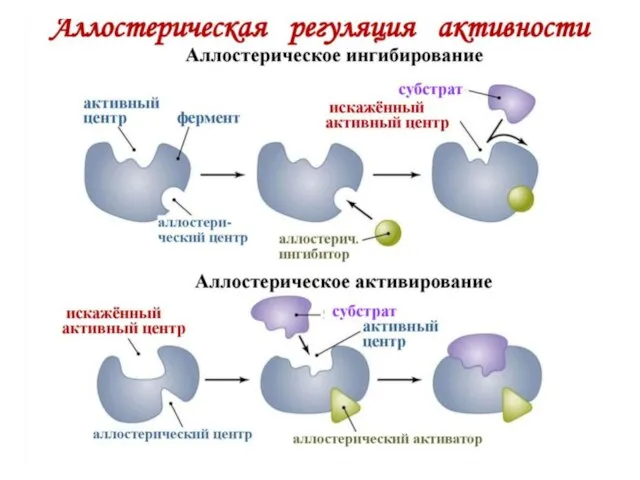
Слайд 47АКТИВАЦИЯ И ИНГИБИРОВАНИЕ АКТИВНОСТИ ФЕРМЕНТОВ
АКТИВАЦИЯ И ИНГИБИРОВАНИЕ АКТИВНОСТИ ФЕРМЕНТОВ

Слайд 48Активаторы ферментов
Вещества, повышающие активность ферментов.
Активаторами могут быть: ионы металлов: натрия, магния, калия,
Активаторы ферментов
Вещества, повышающие активность ферментов.
Активаторами могут быть: ионы металлов: натрия, магния, калия,





Слайд 54Кинетика ферментативной реакции
Активность фермента – количество фермента, катализирующее превращение 1 моля субстрата
Кинетика ферментативной реакции
Активность фермента – количество фермента, катализирующее превращение 1 моля субстрата

Слайд 55ВЛИЯНИЕ КОНЦЕНТРАЦИИ ФЕРМЕНТА И СУБСТРАТА НА СКОРОСТЬ ФЕРМЕНТАТИВНОЙ РЕАКЦИИ
При постоянной концентрации фермента
ВЛИЯНИЕ КОНЦЕНТРАЦИИ ФЕРМЕНТА И СУБСТРАТА НА СКОРОСТЬ ФЕРМЕНТАТИВНОЙ РЕАКЦИИ
При постоянной концентрации фермента

Слайд 56С повышением концентрации фермента при избытке субстрата скорость реакции линейно возрастает
Влияние
С повышением концентрации фермента при избытке субстрата скорость реакции линейно возрастает
Влияние
Слайд 57Количественное соотношение между концентрацией субстрата и скоростью ферментативной реакции выражается уравнением Михаэлиса-Ментен
Количественное соотношение между концентрацией субстрата и скоростью ферментативной реакции выражается уравнением Михаэлиса-Ментен


 Отряды насекомых
Отряды насекомых Океанариум (фотографии)
Океанариум (фотографии) Временная организация клетки. Жизненный цикл клетки, его периодизация
Временная организация клетки. Жизненный цикл клетки, его периодизация Анатомия кровеносной системы
Анатомия кровеносной системы Биохимия крови
Биохимия крови Англійський бульдог
Англійський бульдог Презентация на тему Биоэкология
Презентация на тему Биоэкология  Действующие на верхнюю конечность мышцы туловища
Действующие на верхнюю конечность мышцы туловища Хвойные деревья
Хвойные деревья Презентация на тему Строение и соединение костей
Презентация на тему Строение и соединение костей  Многообразие живых организмов. Одноклеточные и многоклеточные организмы. 5 класс
Многообразие живых организмов. Одноклеточные и многоклеточные организмы. 5 класс Информация для сравнительной таблицы
Информация для сравнительной таблицы Биоинженерия. Мутагенез
Биоинженерия. Мутагенез Самовосстанавливающиеся материалы
Самовосстанавливающиеся материалы Обмен веществ
Обмен веществ Презентация на тему Цветки и плоды
Презентация на тему Цветки и плоды  История генетики
История генетики Клеточное строение организма 8 класс Учитель биологии Кожевникова Маргарита Михайловна МКОУ ХМР СОШ д.Согом
Клеточное строение организма 8 класс Учитель биологии Кожевникова Маргарита Михайловна МКОУ ХМР СОШ д.Согом Строение и свойства ферментов
Строение и свойства ферментов Андский кондор – найбільший літаючий птах
Андский кондор – найбільший літаючий птах Исследования Конрада Лоренца. Импринтинг, его характеристика и виды. Импринтинг как форма научения
Исследования Конрада Лоренца. Импринтинг, его характеристика и виды. Импринтинг как форма научения Что изучает биология
Что изучает биология Самые умные животные
Самые умные животные Аксолотль
Аксолотль Кости человека
Кости человека Вирусы
Вирусы Путешествие в микромир
Путешествие в микромир Ирис
Ирис