- Физиология слуха и равновесия

Содержание
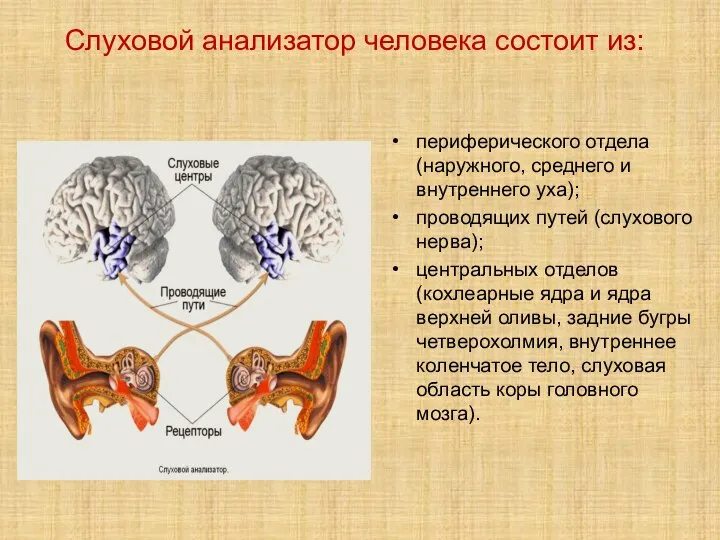
- 2. Слуховой анализатор человека состоит из: периферического отдела (наружного, среднего и внутреннего уха); проводящих путей (слухового нерва);
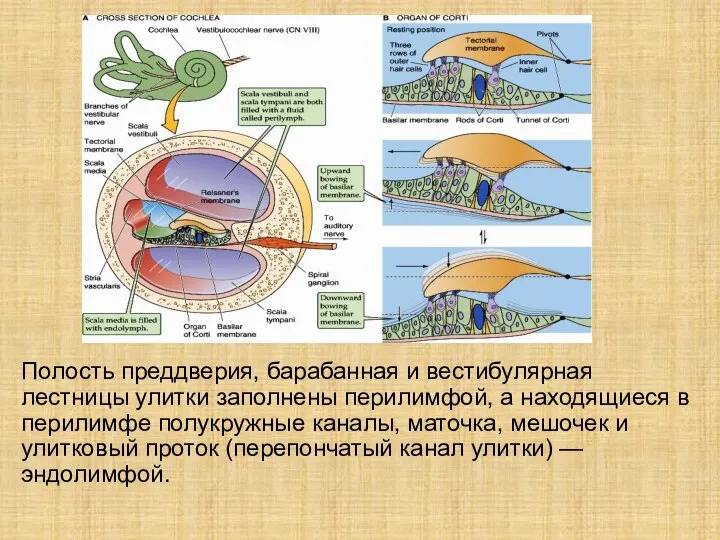
- 6. Полость преддверия, барабанная и вестибулярная лестницы улитки заполнены перилимфой, а находящиеся в перилимфе полукружные каналы, маточка,
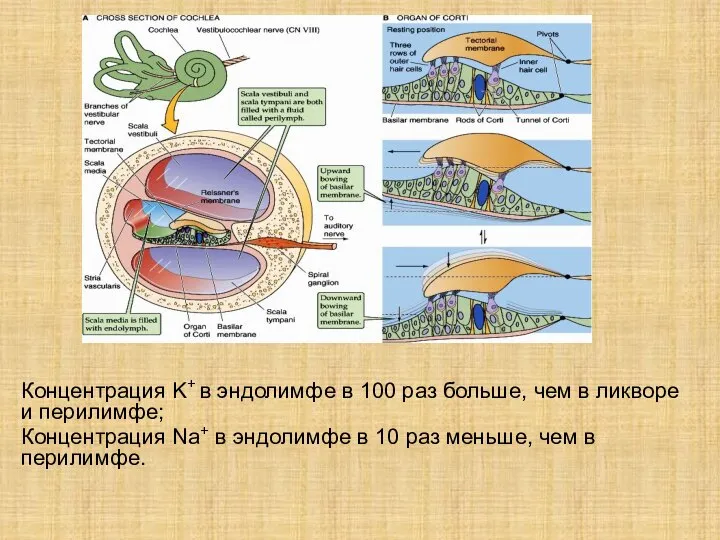
- 7. Концентрация K+ в эндолимфе в 100 раз больше, чем в ликворе и перилимфе; Концентрация Na+ в
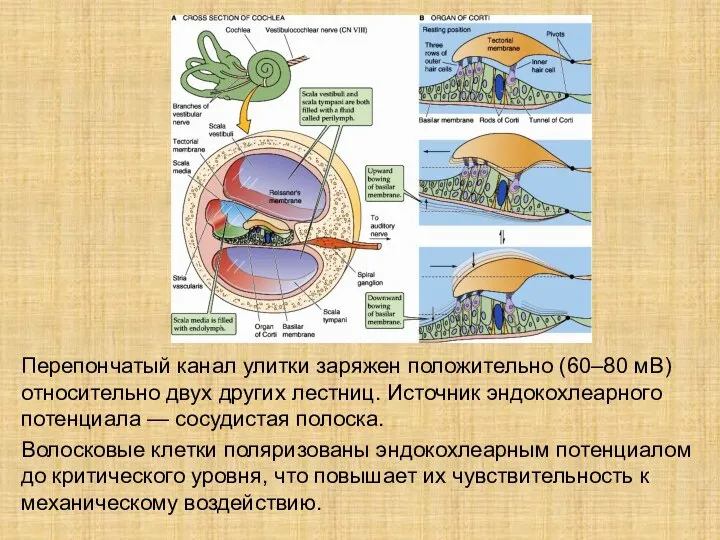
- 8. Перепончатый канал улитки заряжен положительно (60–80 мВ) относительно двух других лестниц. Источник эндокохлеарного потенциала — сосудистая
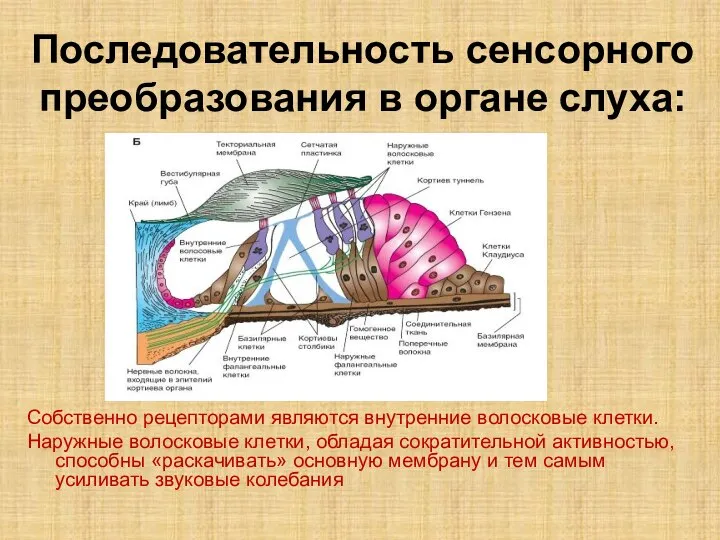
- 9. Последовательность сенсорного преобразования в органе слуха: Собственно рецепторами являются внутренние волосковые клетки. Наружные волосковые клетки, обладая
- 11. Сканирующая электронная микрофотография волосковых клеток кортиева органа Покровная мембрана удалена, а микроскоп направлен на базиллярную мембрану.
- 12. Волосковые клетки
- 16. Высота звука - это субъективное восприятие частоты звуковых колебаний. Человеческое ухо воспринимает частоты в диапазоне от
- 17. 2. Временным способом (20 – 200 Гц) Информация кодируется разной частотой импульсации в одних и тех
- 18. Острота слуха Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек в 50% случаев
- 19. Сила звука Диапазон силы звука, воспринимаемый человеческим ухом, огромен (болевой порог в 1013 раз выше порога
- 20. Слуховые пути и центры кохлеарное ядро (нейроны 2-го порядка), комплекс ядер верхней оливы и трапецевидного тела
- 21. Направление источника звука определяется 2 путями: При звуке высокой частоты (звук отражается от головы) источник оценивается
- 22. Слуховая кора Первичная слуховая кора непосредственно получает сигналы от медиального коленчатого тела, в то время как
- 23. Чувство равновесия
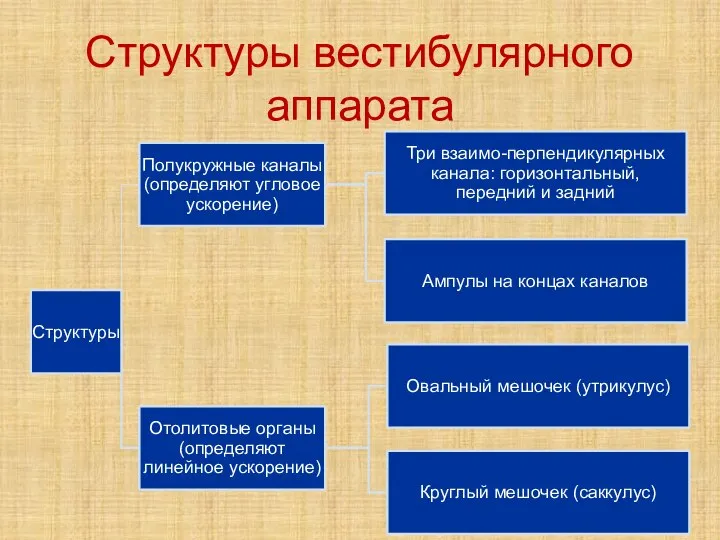
- 25. Структуры вестибулярного аппарата
- 26. Полукружные каналы работают в парах (билатерально) При вращение головы вправо: Волосковые клетки с правой стороны гиперполяризуются
- 27. Определение углового ускорения Полукружные каналы Примерно ортогональны друг другу Оба конца заканчиваются в овальном мешочке (место
- 28. Определение линейного ускорения Отолитовые органы: Активный участок – макула – представлена волосковыми клетками, отолитовой мембраной и
- 30. Волосковые клетки При возбуждении в волосковых клетках генерируется рецепторный потенциал и происходит выброс ацетилхолина, который и
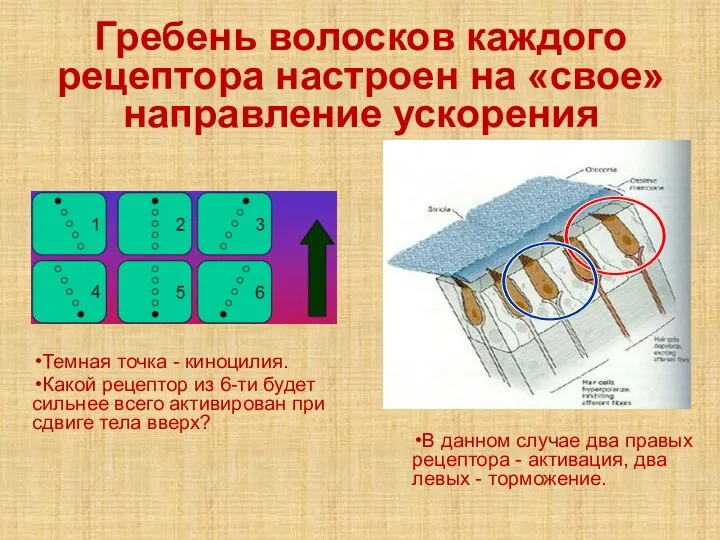
- 31. Гребень волосков каждого рецептора настроен на «свое» направление ускорения Темная точка - киноцилия. Какой рецептор из
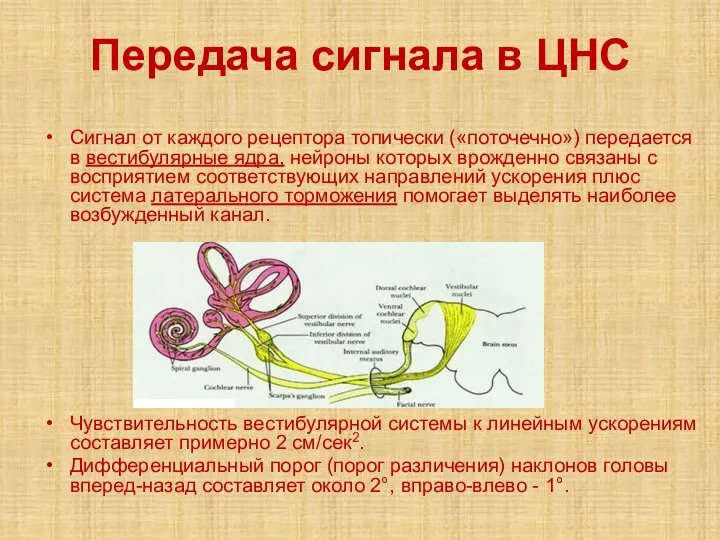
- 32. Передача сигнала в ЦНС Сигнал от каждого рецептора топически («поточечно») передается в вестибулярные ядра, нейроны которых
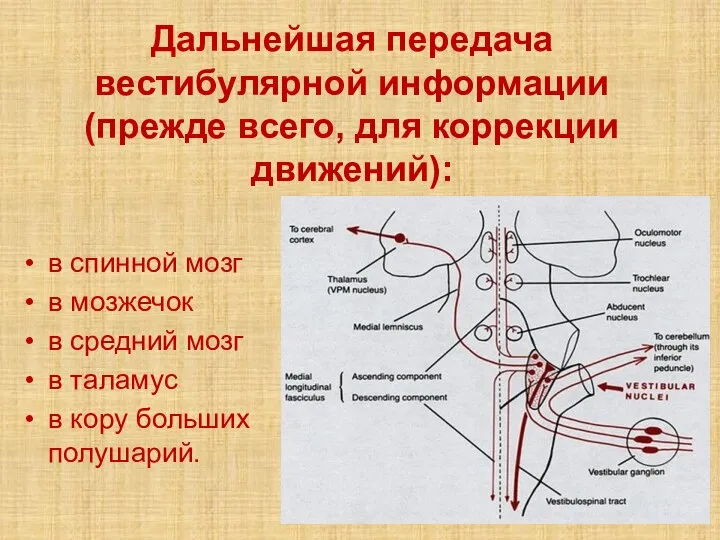
- 33. Дальнейшая передача вестибулярной информации (прежде всего, для коррекции движений): в спинной мозг в мозжечок в средний
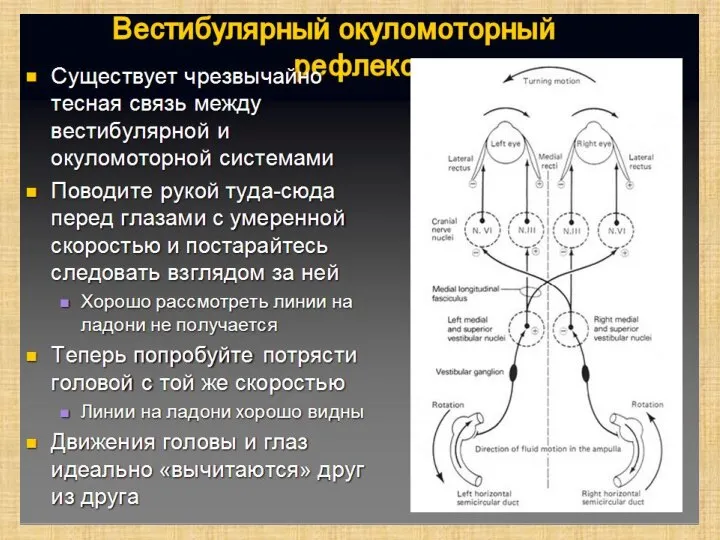
- 34. Вестибулярная информация необходима: Спинному мозгу (от ядра Дейтерса и медиального ядер по латеральному и медиальному вестибулоспинальным
- 36. Укачивание Укачивание - иррадиация возбуждения по центрам ромбовидной ямки. Субъективные ощущения: головокружение, тошнота и другие реакции,
- 37. Вестибулярная система как источник эмоций (новизна ощущений!)
- 38. Обонятельная сенсорная система
- 39. Классификация запахов по Эймуэру Первичные или основные: - камфорный - (камфора, 1,8-цинеол) - острый или едкий
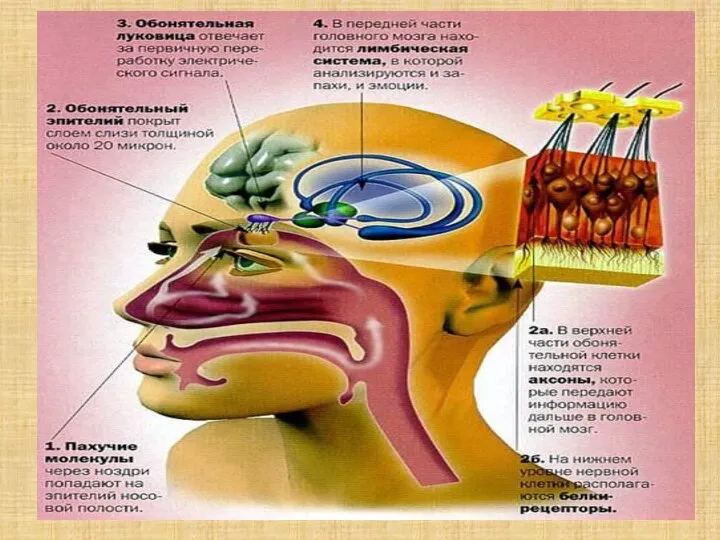
- 40. Воспринимающие обонятельные структуры образуют в слизистой оболочке носа специализированную область — парную обонятельную выстилку (обонятельное поле),
- 41. Втягивание воздуха Область, содержащая обонятельные рецепторы, плохо вентилируется. Улучшение их вентиляции обеспечивается принюхиванием. Втягивание воздуха —
- 42. Рецепторный аппарат Обонятельный эпителий содержит опорные эпителиальные клетки и расположенные между ними примерно 100 млн рецепторных
- 43. От булавы отходит от 8 до 40 тонких обонятельных волосков — ресничек. В волоски встроены обонятельные
- 44. ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА В обонятельной луковице аксоны рецепторных клеток образуют синапсы с дендритами митральных и пучковых клеток,
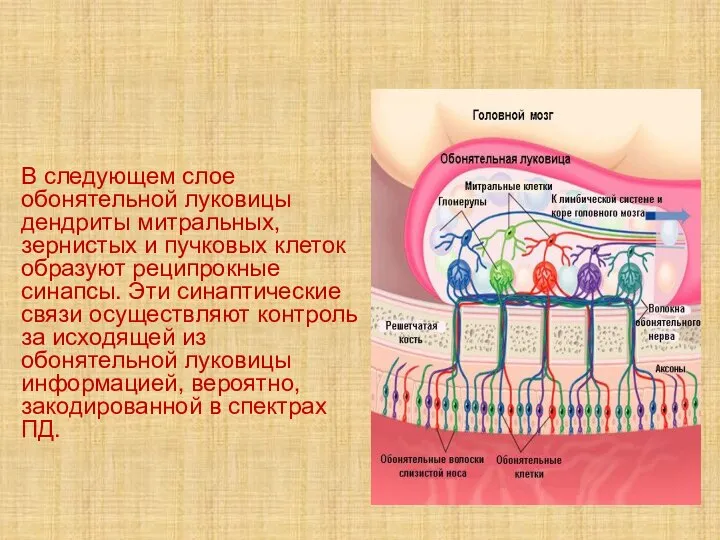
- 45. В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых клеток образуют реципрокные синапсы. Эти синаптические
- 46. Регистрация обонятельного сигнала взаимодействие одоранта с рецептором в плазмолемме обонятельных волосков активация G‑белка повышение активности аденилатциклазы
- 47. Через цАМФ–зависимые воротные ионные каналы внутрь клетки проходят не только одновалентные катионы, но и Ca2+, связывающийся
- 48. Концепция комбинаторного кодирования Каждый из миллионов одорантов имеет уникальный код; кодирование происходит на уровне органа обоняния,
- 49. Пути передачи обонятельной информации Аксоны митральных клеток в составе обонятельного тракта поднимаются в первичные обонятельные центры
- 50. Пути передачи обонятельной информации Аксоны нервных клеток латеральной обонятельной области, расположенных в грушевидной коре и миндалевидном
- 52. Вкусовая сенсорная система
- 53. Основные вкусы Человек различает четыре первичных вкуса: сладкий, кислый, горький солёный, а также «умами» (от японского
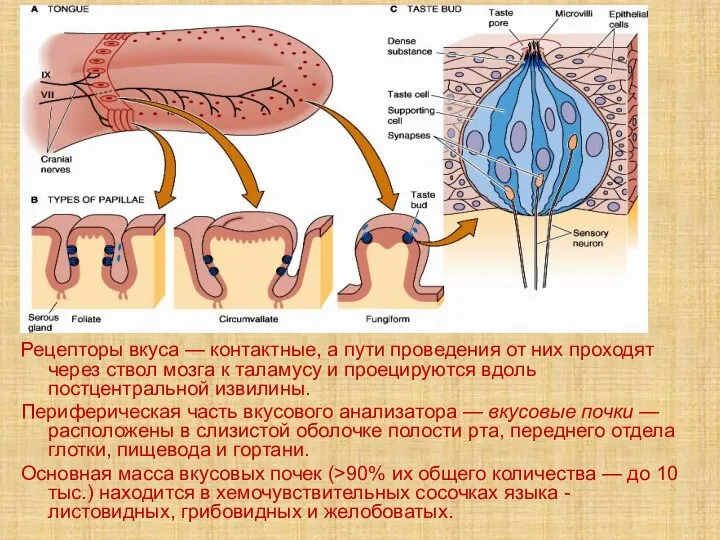
- 54. Рецепторы вкуса — контактные, а пути проведения от них проходят через ствол мозга к таламусу и
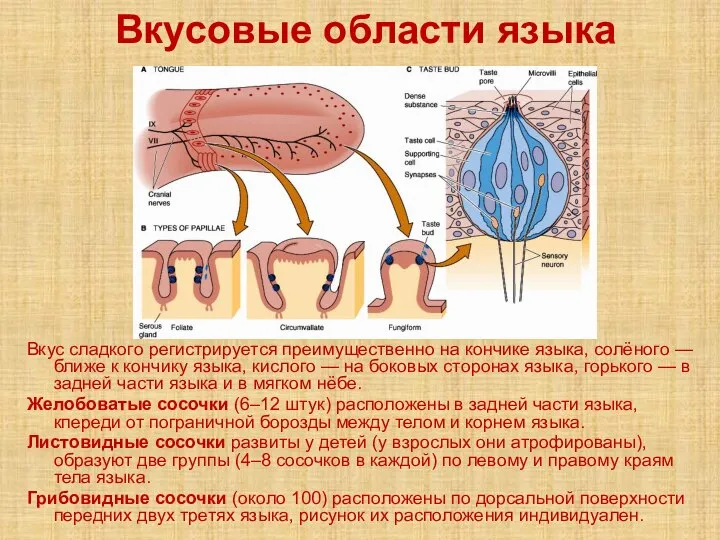
- 55. Вкусовые области языка Вкус сладкого регистрируется преимущественно на кончике языка, солёного — ближе к кончику языка,
- 56. Вкусовая почка состоит из удлинённых светлых клеток различных типов. - На апикальной части микроворсинки, занимающие вкусовой
- 57. Различие вкусовых клеток При низких концентрациях деполяризуются и формируют рецепторный потенциал лишь отдельные вкусовые клетки. Некоторые
- 58. Восприятие соленого и кислого Соленый стимул взаимодействует с эпителиальными натриевыми каналами (ENaC), открывая их для натрия.
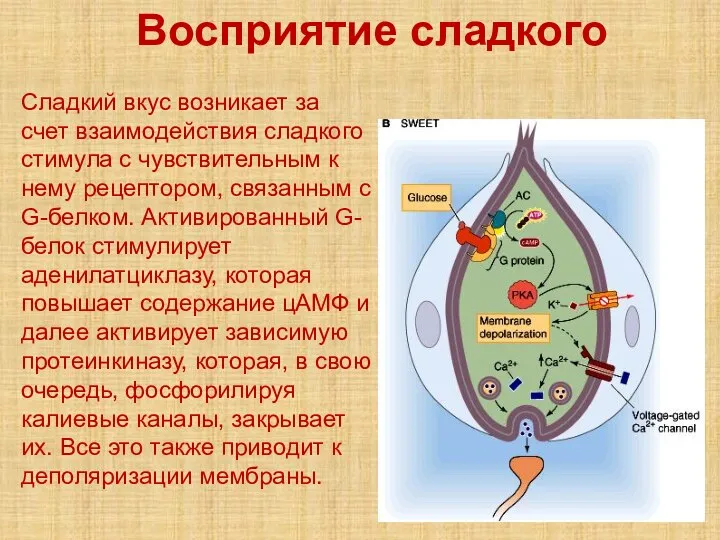
- 59. Восприятие сладкого Сладкий вкус возникает за счет взаимодействия сладкого стимула с чувствительным к нему рецептором, связанным
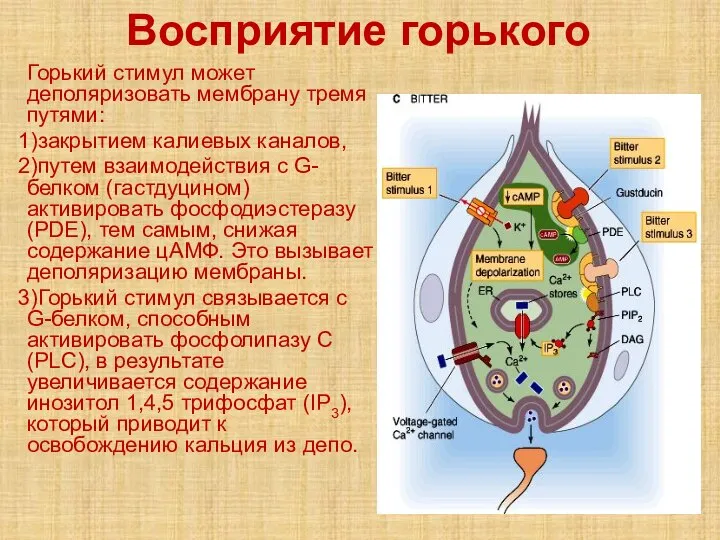
- 60. Восприятие горького Горький стимул может деполяризовать мембрану тремя путями: закрытием калиевых каналов, путем взаимодействия с G-белком
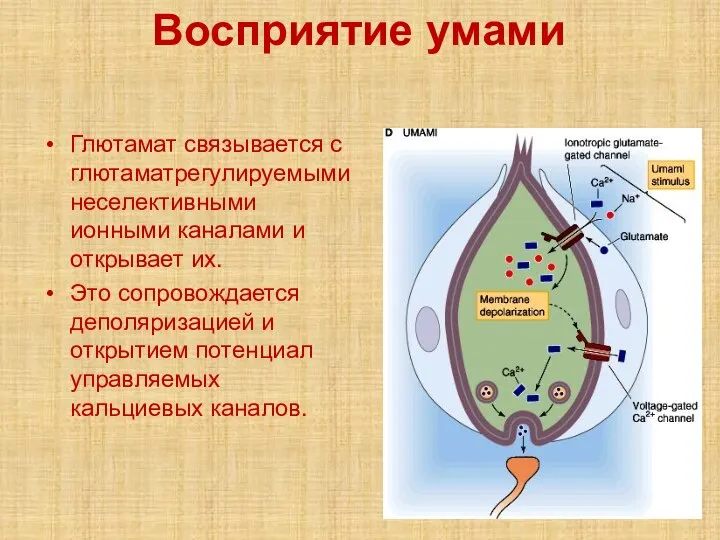
- 61. Восприятие умами Глютамат связывается с глютаматрегулируемыми неселективными ионными каналами и открывает их. Это сопровождается деполяризацией и
- 62. ВКУСОВОЙ ПОРОГ Наиболее значительные различия наблюдают между веществами, создающими ощущение горького и всех остальных первичных вкусов.
- 63. АДАПТАЦИЯ Адаптация к воздействию вкусового вещества развивается медленно (минуты) и пропорциональна его концентрации. К сладкому и
- 64. Восходящие пути вкусовой чувствительности состоят из трёх нейронов: 1. первичного чувствительного нейрона - от передних двух
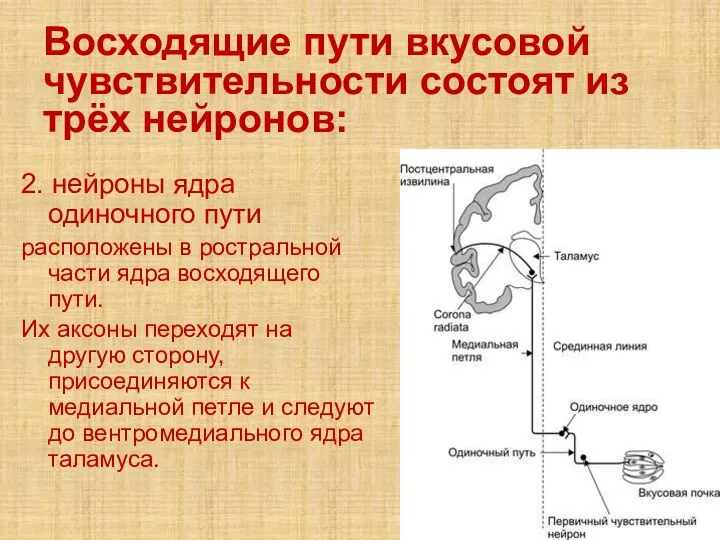
- 65. Восходящие пути вкусовой чувствительности состоят из трёх нейронов: 2. нейроны ядра одиночного пути расположены в ростральной
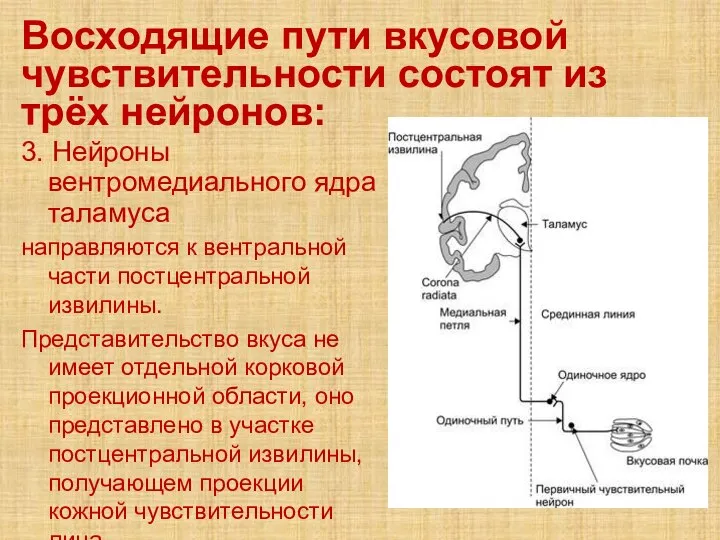
- 66. Восходящие пути вкусовой чувствительности состоят из трёх нейронов: 3. Нейроны вентромедиального ядра таламуса направляются к вентральной
- 67. Тактильная сенсорная система
- 68. Тактильные ощущения прикосновения, давления и вибрации относятся к раздельным видам ощущений, но воспринимаются одними и теми
- 69. Рецепторы кожи Свободные нервные окончания образуют механорецепторы (прикосновение и давление), терморецепторы и рецепторы болевой чувствительности. Тельца
- 70. Количество и плотность размещения рецепторов - Общее количество тактильных рецепторов оценивается в 10 миллионов штук, сгруппированных
- 71. Разрешающая способность и пороги ощущений: - Порог ощущения кожных рецепторов при надавливании: 10 мкм - Одновременный
- 72. Адаптация
- 73. Передача импульсации Передача тактильных ощущений тонкой дифференцировки происходит по миелинизированным нервным волокнам со скоростью 30-70 м/с.
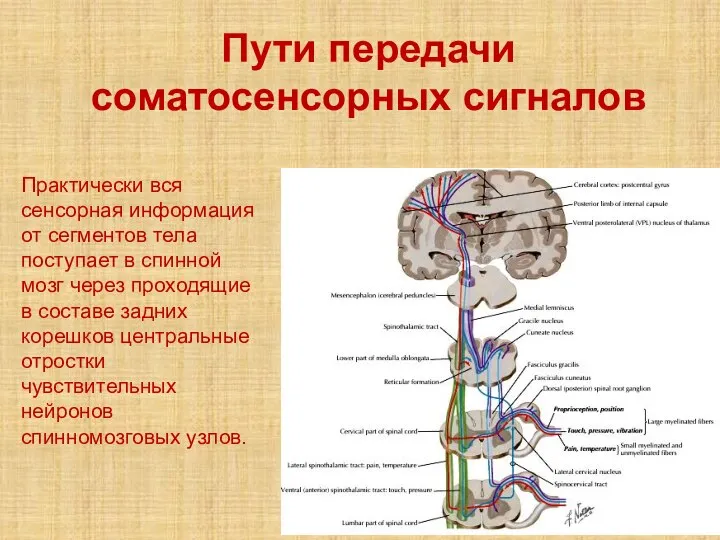
- 74. Пути передачи соматосенсорных сигналов Практически вся сенсорная информация от сегментов тела поступает в спинной мозг через
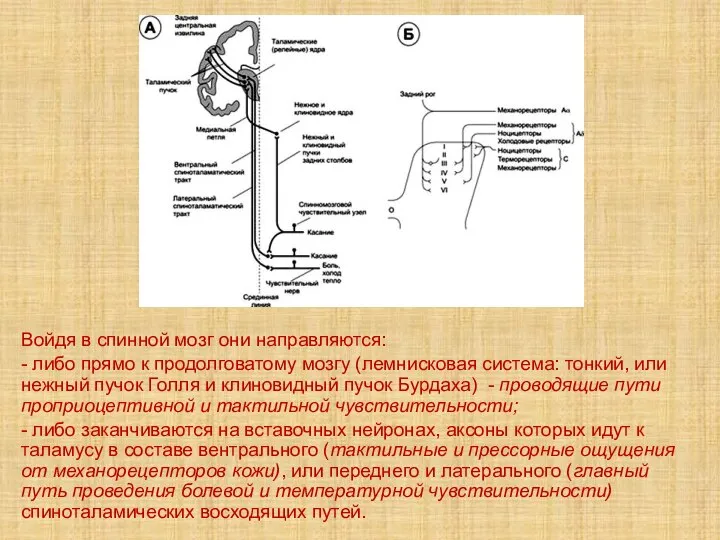
- 75. Войдя в спинной мозг они направляются: - либо прямо к продолговатому мозгу (лемнисковая система: тонкий, или
- 77. Скачать презентацию
Слайд 2Слуховой анализатор человека состоит из:
периферического отдела (наружного, среднего и внутреннего уха);
Слуховой анализатор человека состоит из:
периферического отдела (наружного, среднего и внутреннего уха);
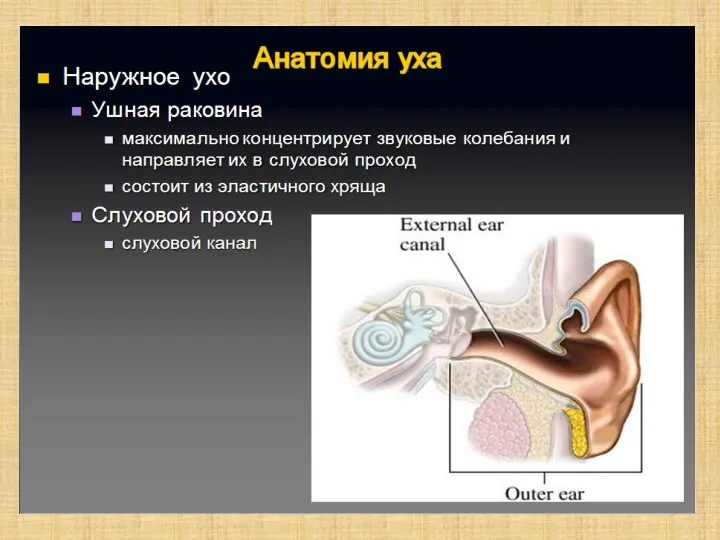
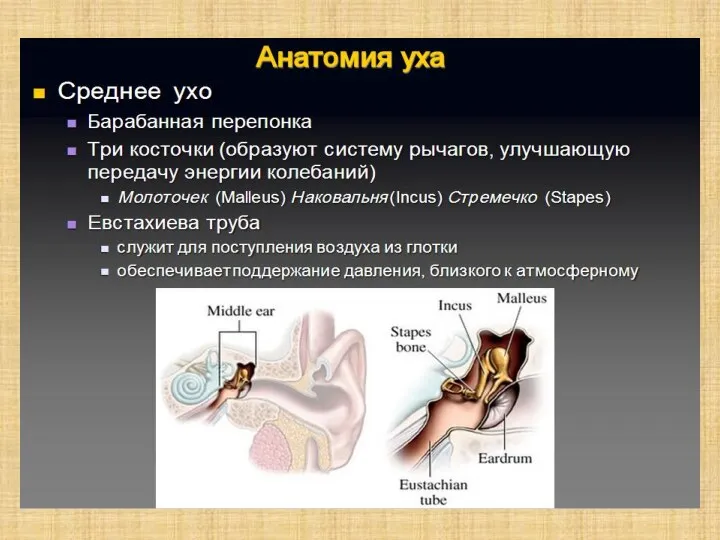
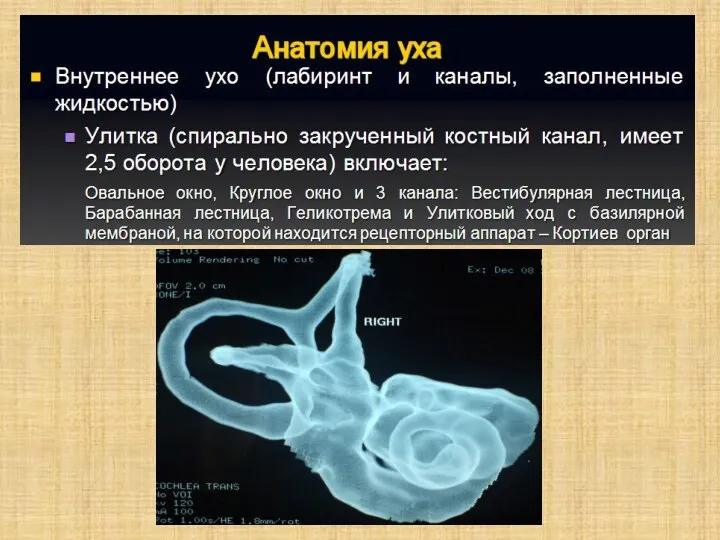
Слайд 6Полость преддверия, барабанная и вестибулярная лестницы улитки заполнены перилимфой, а находящиеся в
Полость преддверия, барабанная и вестибулярная лестницы улитки заполнены перилимфой, а находящиеся в
Слайд 7Концентрация K+ в эндолимфе в 100 раз больше, чем в ликворе и
Концентрация K+ в эндолимфе в 100 раз больше, чем в ликворе и
Слайд 8Перепончатый канал улитки заряжен положительно (60–80 мВ) относительно двух других лестниц. Источник
Перепончатый канал улитки заряжен положительно (60–80 мВ) относительно двух других лестниц. Источник
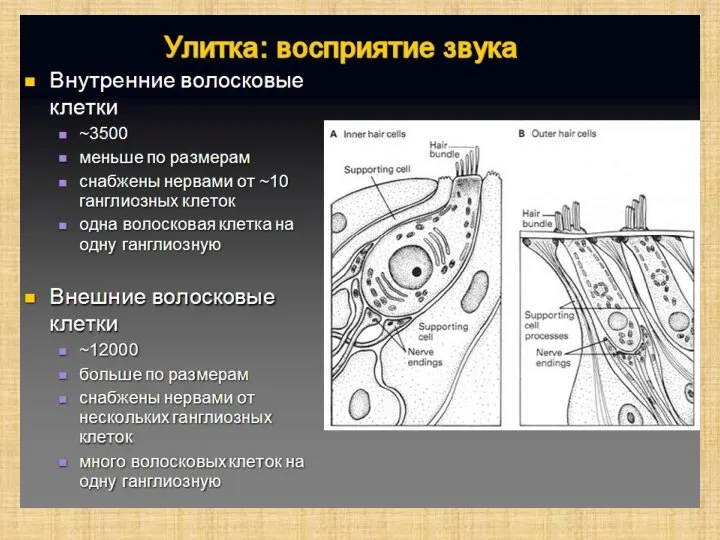
Слайд 9Последовательность сенсорного преобразования в органе слуха:
Собственно рецепторами являются внутренние волосковые клетки.
Наружные волосковые клетки,
Последовательность сенсорного преобразования в органе слуха:
Собственно рецепторами являются внутренние волосковые клетки.
Наружные волосковые клетки,
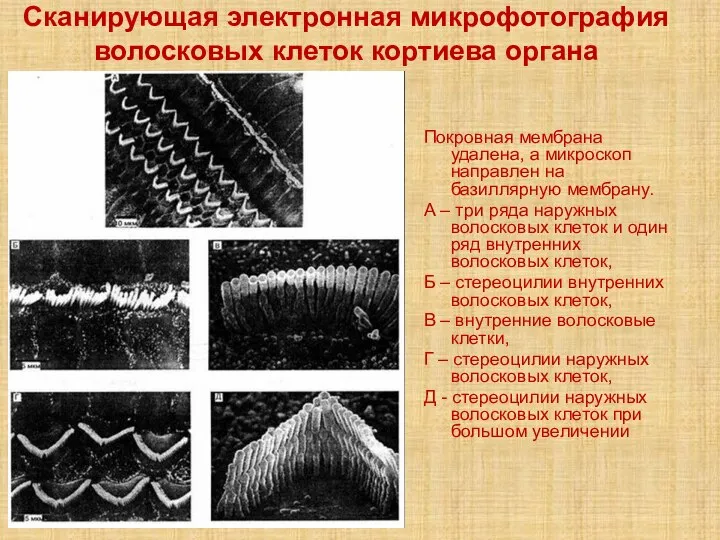
Слайд 11Сканирующая электронная микрофотография волосковых клеток кортиева органа
Покровная мембрана удалена, а микроскоп направлен
Сканирующая электронная микрофотография волосковых клеток кортиева органа
Покровная мембрана удалена, а микроскоп направлен
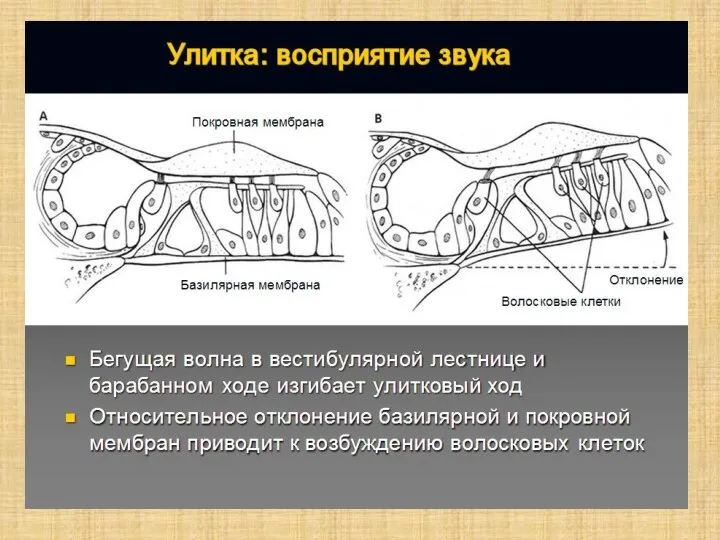
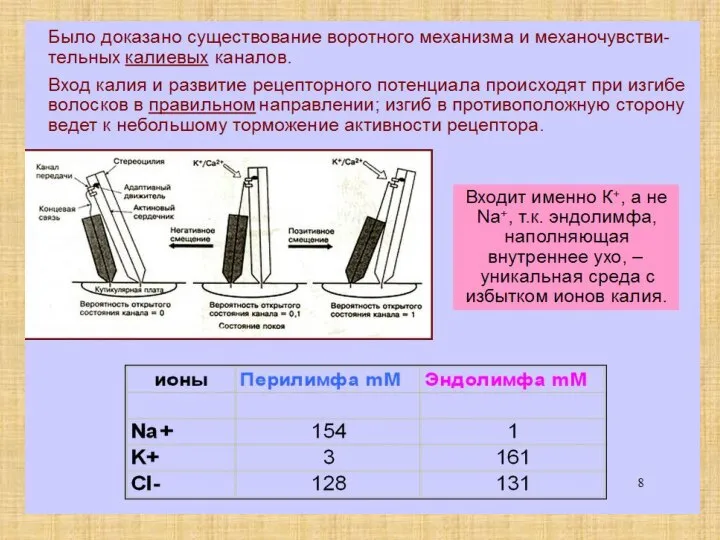
Слайд 12Волосковые клетки
Волосковые клетки

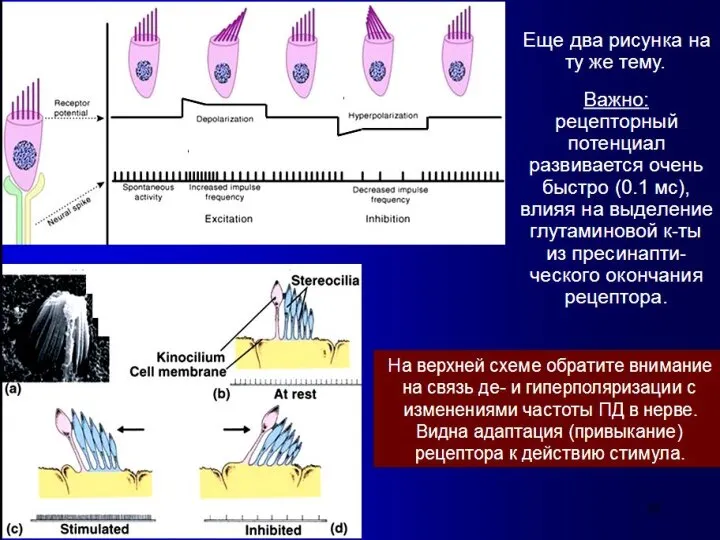
Слайд 16Высота звука
- это субъективное восприятие частоты звуковых колебаний.
Человеческое ухо воспринимает
Высота звука
- это субъективное восприятие частоты звуковых колебаний.
Человеческое ухо воспринимает

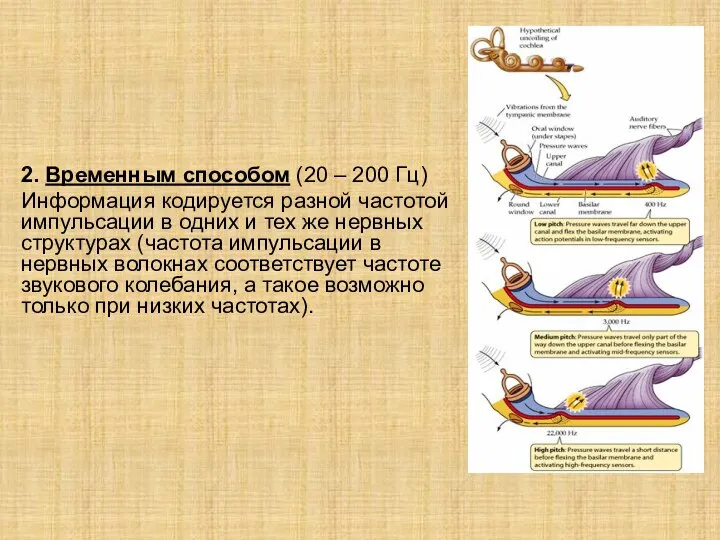
Слайд 172. Временным способом (20 – 200 Гц)
Информация кодируется разной частотой импульсации в одних
2. Временным способом (20 – 200 Гц)
Информация кодируется разной частотой импульсации в одних
Слайд 18Острота слуха
Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек в
Острота слуха
Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек в

Слайд 19Сила звука
Диапазон силы звука, воспринимаемый человеческим ухом, огромен (болевой порог в 1013 раз выше
Сила звука
Диапазон силы звука, воспринимаемый человеческим ухом, огромен (болевой порог в 1013 раз выше

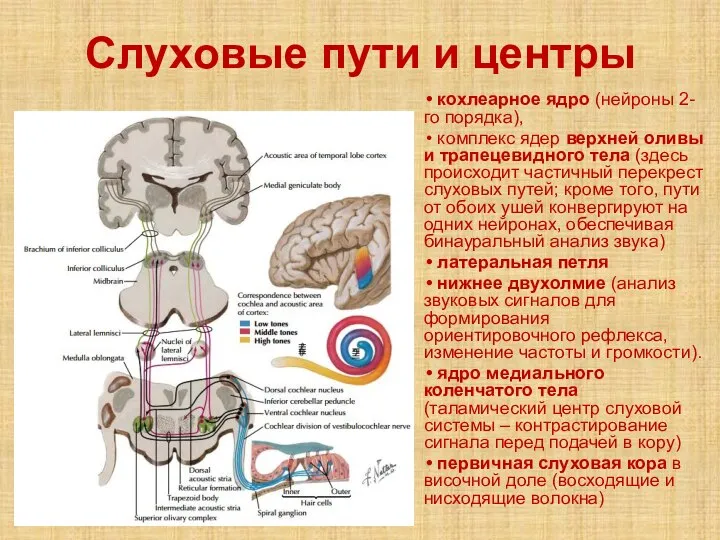
Слайд 20Слуховые пути и центры
кохлеарное ядро (нейроны 2-го порядка),
комплекс ядер
Слуховые пути и центры
кохлеарное ядро (нейроны 2-го порядка),
комплекс ядер
Слайд 21Направление источника звука определяется 2 путями:
При звуке высокой частоты (звук отражается от
Направление источника звука определяется 2 путями:
При звуке высокой частоты (звук отражается от

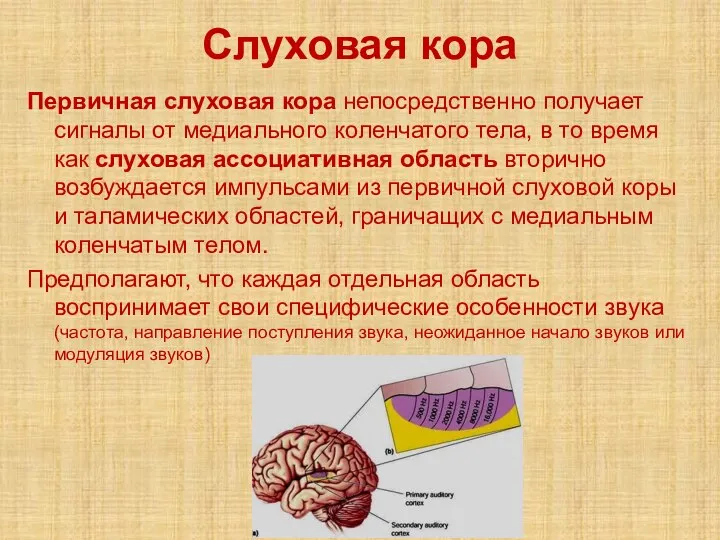
Слайд 22Слуховая кора
Первичная слуховая кора непосредственно получает сигналы от медиального коленчатого тела, в
Слуховая кора
Первичная слуховая кора непосредственно получает сигналы от медиального коленчатого тела, в
Слайд 23Чувство равновесия
Чувство равновесия


Слайд 25Структуры вестибулярного аппарата
Структуры вестибулярного аппарата
Слайд 26Полукружные каналы работают в парах (билатерально)
При вращение головы вправо:
Волосковые клетки с правой
Полукружные каналы работают в парах (билатерально)
При вращение головы вправо:
Волосковые клетки с правой

Слайд 27Определение углового ускорения
Полукружные каналы
Примерно ортогональны друг другу
Оба конца заканчиваются в овальном мешочке
Определение углового ускорения
Полукружные каналы
Примерно ортогональны друг другу
Оба конца заканчиваются в овальном мешочке

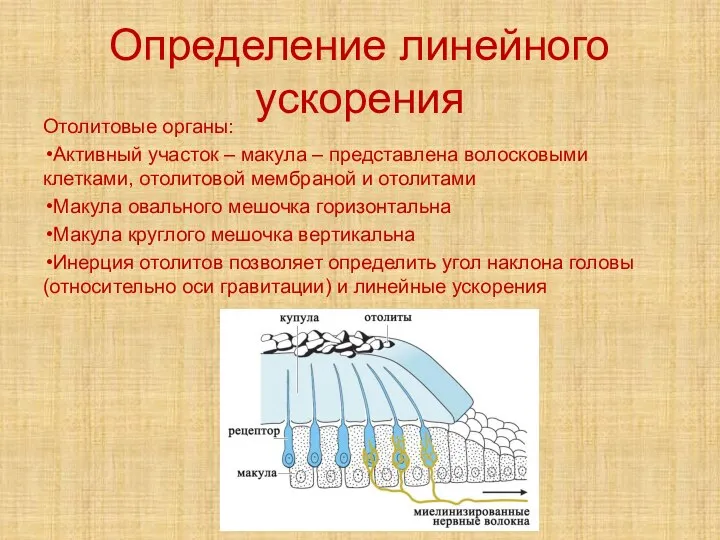
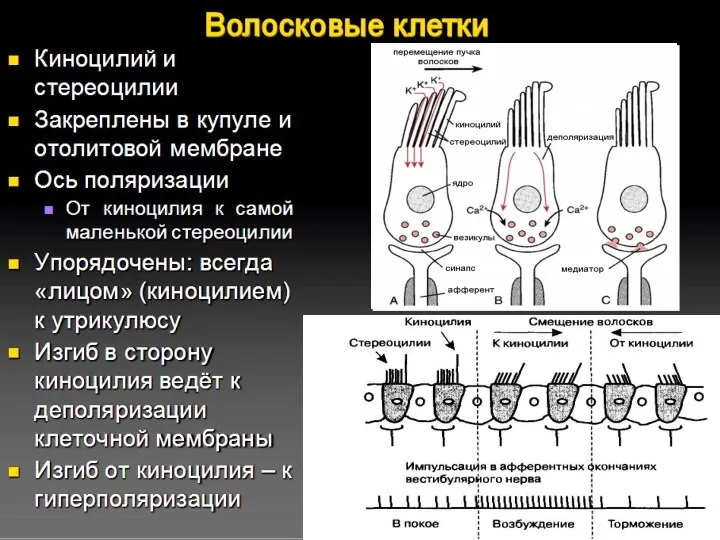
Слайд 28Определение линейного ускорения
Отолитовые органы:
Активный участок – макула – представлена волосковыми клетками, отолитовой
Определение линейного ускорения
Отолитовые органы:
Активный участок – макула – представлена волосковыми клетками, отолитовой
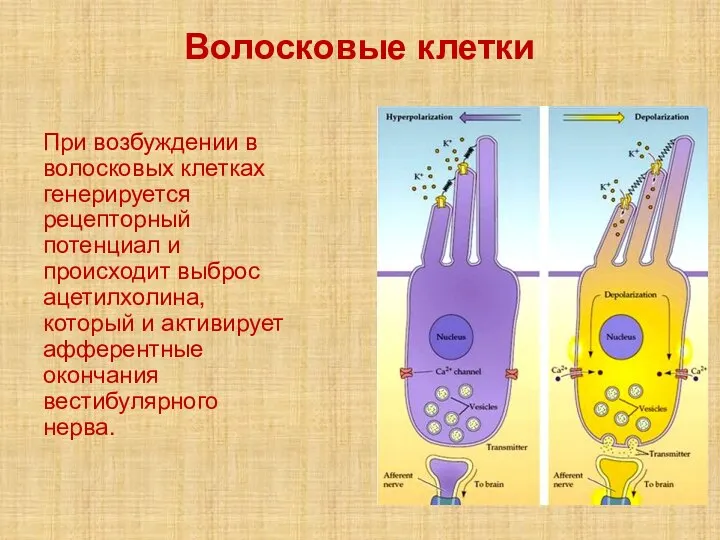
Слайд 30Волосковые клетки
При возбуждении в волосковых клетках генерируется рецепторный потенциал и происходит выброс
Волосковые клетки
При возбуждении в волосковых клетках генерируется рецепторный потенциал и происходит выброс
Слайд 31Гребень волосков каждого рецептора настроен на «свое» направление ускорения
Темная точка - киноцилия.
Гребень волосков каждого рецептора настроен на «свое» направление ускорения
Темная точка - киноцилия.
Слайд 32Передача сигнала в ЦНС
Сигнал от каждого рецептора топически («поточечно») передается в вестибулярные
Передача сигнала в ЦНС
Сигнал от каждого рецептора топически («поточечно») передается в вестибулярные
Слайд 33Дальнейшая передача вестибулярной информации (прежде всего, для коррекции движений):
в спинной мозг
в
Дальнейшая передача вестибулярной информации (прежде всего, для коррекции движений):
в спинной мозг
в
Слайд 34Вестибулярная информация необходима:
Спинному мозгу (от ядра Дейтерса и медиального ядер по латеральному
Вестибулярная информация необходима:
Спинному мозгу (от ядра Дейтерса и медиального ядер по латеральному

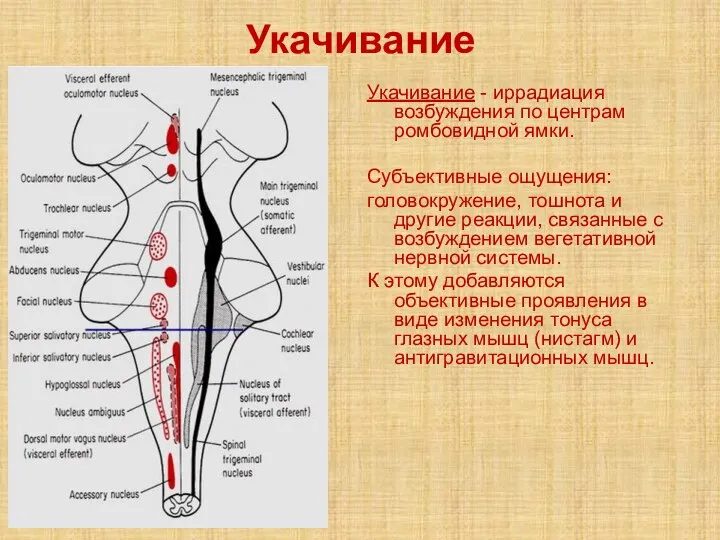
Слайд 36Укачивание
Укачивание - иррадиация возбуждения по центрам ромбовидной ямки.
Субъективные ощущения:
головокружение, тошнота и другие
Укачивание
Укачивание - иррадиация возбуждения по центрам ромбовидной ямки.
Субъективные ощущения:
головокружение, тошнота и другие
Слайд 37Вестибулярная система как источник эмоций
(новизна ощущений!)
Вестибулярная система как источник эмоций
(новизна ощущений!)

Слайд 38Обонятельная сенсорная система
Обонятельная сенсорная система

Слайд 39Классификация запахов по Эймуэру
Первичные или основные:
- камфорный - (камфора, 1,8-цинеол)
-
Классификация запахов по Эймуэру
Первичные или основные:
- камфорный - (камфора, 1,8-цинеол)
-

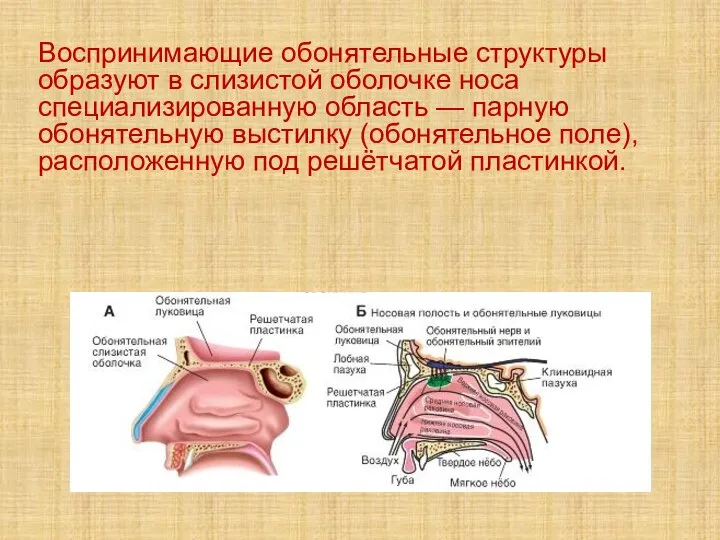
Слайд 40Воспринимающие обонятельные структуры образуют в слизистой оболочке носа специализированную область — парную обонятельную
Воспринимающие обонятельные структуры образуют в слизистой оболочке носа специализированную область — парную обонятельную
Слайд 41Втягивание воздуха
Область, содержащая обонятельные рецепторы, плохо вентилируется. Улучшение их вентиляции обеспечивается принюхиванием.
Втягивание
Втягивание воздуха
Область, содержащая обонятельные рецепторы, плохо вентилируется. Улучшение их вентиляции обеспечивается принюхиванием.
Втягивание

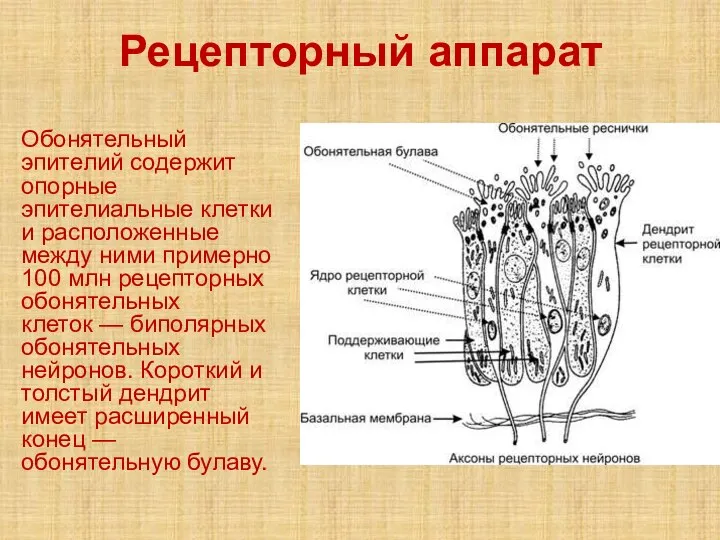
Слайд 42Рецепторный аппарат
Обонятельный эпителий содержит опорные эпителиальные клетки и расположенные между ними примерно
Рецепторный аппарат
Обонятельный эпителий содержит опорные эпителиальные клетки и расположенные между ними примерно
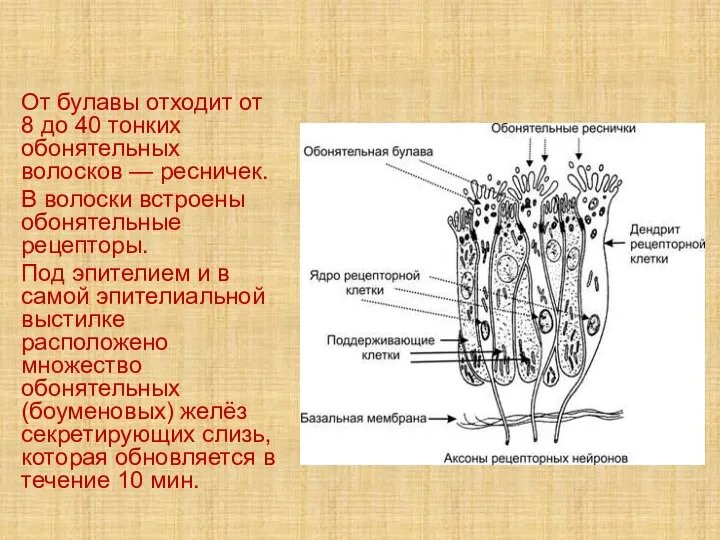
Слайд 43От булавы отходит от 8 до 40 тонких обонятельных волосков — ресничек.
В
От булавы отходит от 8 до 40 тонких обонятельных волосков — ресничек.
В
Слайд 44ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА
В обонятельной луковице аксоны рецепторных клеток образуют синапсы с дендритами митральных
ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА
В обонятельной луковице аксоны рецепторных клеток образуют синапсы с дендритами митральных
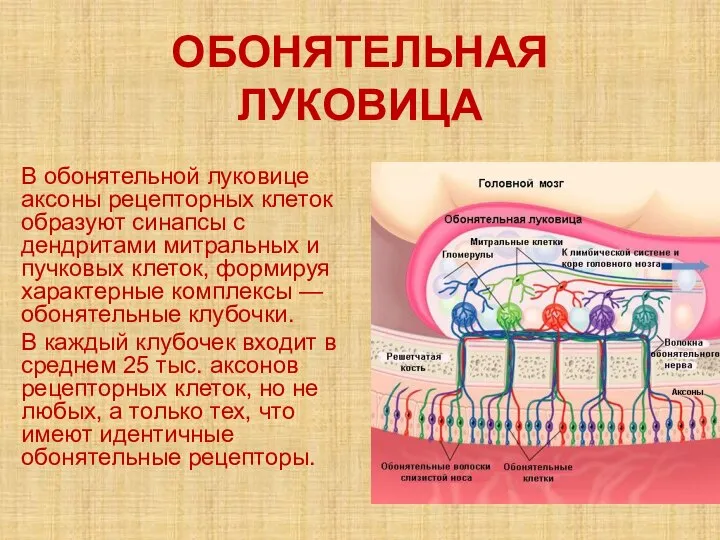
Слайд 45В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых клеток образуют
В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых клеток образуют
Слайд 46Регистрация обонятельного сигнала
взаимодействие одоранта с рецептором в плазмолемме обонятельных волосков
активация G‑белка
повышение
Регистрация обонятельного сигнала
взаимодействие одоранта с рецептором в плазмолемме обонятельных волосков
активация G‑белка
повышение

Слайд 47Через цАМФ–зависимые воротные ионные каналы внутрь клетки проходят не только одновалентные катионы,
Через цАМФ–зависимые воротные ионные каналы внутрь клетки проходят не только одновалентные катионы,

Слайд 48Концепция комбинаторного кодирования
Каждый из миллионов одорантов имеет уникальный код; кодирование происходит на
Концепция комбинаторного кодирования
Каждый из миллионов одорантов имеет уникальный код; кодирование происходит на

Слайд 49Пути передачи обонятельной информации
Аксоны митральных клеток в составе обонятельного тракта поднимаются в
Пути передачи обонятельной информации
Аксоны митральных клеток в составе обонятельного тракта поднимаются в

Слайд 50Пути передачи обонятельной информации
Аксоны нервных клеток латеральной обонятельной области, расположенных в грушевидной
Пути передачи обонятельной информации
Аксоны нервных клеток латеральной обонятельной области, расположенных в грушевидной

Слайд 52Вкусовая сенсорная система
Вкусовая сенсорная система
Слайд 53Основные вкусы
Человек различает четыре первичных вкуса:
сладкий,
кислый,
горький
солёный,
а также «умами»
Основные вкусы
Человек различает четыре первичных вкуса:
сладкий,
кислый,
горький
солёный,
а также «умами»

Слайд 54Рецепторы вкуса — контактные, а пути проведения от них проходят через ствол мозга
Рецепторы вкуса — контактные, а пути проведения от них проходят через ствол мозга
Слайд 55Вкусовые области языка
Вкус сладкого регистрируется преимущественно на кончике языка, солёного — ближе к
Вкусовые области языка
Вкус сладкого регистрируется преимущественно на кончике языка, солёного — ближе к
Слайд 56Вкусовая почка
состоит из удлинённых светлых клеток различных типов.
- На апикальной части
Вкусовая почка
состоит из удлинённых светлых клеток различных типов.
- На апикальной части

Слайд 57Различие вкусовых клеток
При низких концентрациях деполяризуются и формируют рецепторный потенциал лишь отдельные
Различие вкусовых клеток
При низких концентрациях деполяризуются и формируют рецепторный потенциал лишь отдельные

Слайд 58Восприятие соленого и кислого
Соленый стимул взаимодействует с эпителиальными натриевыми каналами (ENaC), открывая
Восприятие соленого и кислого
Соленый стимул взаимодействует с эпителиальными натриевыми каналами (ENaC), открывая

Слайд 59Восприятие сладкого
Сладкий вкус возникает за счет взаимодействия сладкого стимула с чувствительным к
Восприятие сладкого
Сладкий вкус возникает за счет взаимодействия сладкого стимула с чувствительным к
Слайд 60Восприятие горького
Горький стимул может деполяризовать мембрану тремя путями:
закрытием калиевых каналов,
путем
Восприятие горького
Горький стимул может деполяризовать мембрану тремя путями:
закрытием калиевых каналов,
путем
Слайд 61Восприятие умами
Глютамат связывается с глютаматрегулируемыми неселективными ионными каналами и открывает их.
Это
Восприятие умами
Глютамат связывается с глютаматрегулируемыми неселективными ионными каналами и открывает их.
Это
Слайд 62ВКУСОВОЙ ПОРОГ
Наиболее значительные различия наблюдают между веществами, создающими ощущение горького и всех
ВКУСОВОЙ ПОРОГ
Наиболее значительные различия наблюдают между веществами, создающими ощущение горького и всех

Слайд 63АДАПТАЦИЯ
Адаптация к воздействию вкусового вещества развивается медленно (минуты) и пропорциональна его концентрации.
АДАПТАЦИЯ
Адаптация к воздействию вкусового вещества развивается медленно (минуты) и пропорциональна его концентрации.

Слайд 64Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
1. первичного чувствительного нейрона
- от
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
1. первичного чувствительного нейрона
- от

Слайд 65Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
2. нейроны ядра одиночного
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
2. нейроны ядра одиночного
Слайд 66Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
3. Нейроны вентромедиального ядра таламуса
направляются
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
3. Нейроны вентромедиального ядра таламуса
направляются
Слайд 67Тактильная сенсорная система
Тактильная сенсорная система

Слайд 68Тактильные ощущения прикосновения, давления и вибрации относятся к раздельным видам ощущений, но
Тактильные ощущения прикосновения, давления и вибрации относятся к раздельным видам ощущений, но

Слайд 69Рецепторы кожи
Свободные нервные окончания образуют механорецепторы (прикосновение и давление), терморецепторы и рецепторы болевой
Рецепторы кожи
Свободные нервные окончания образуют механорецепторы (прикосновение и давление), терморецепторы и рецепторы болевой

Слайд 70Количество и плотность размещения рецепторов
- Общее количество тактильных рецепторов оценивается в 10
Количество и плотность размещения рецепторов
- Общее количество тактильных рецепторов оценивается в 10

Слайд 71Разрешающая способность и пороги ощущений:
- Порог ощущения кожных рецепторов при надавливании: 10
Разрешающая способность и пороги ощущений:
- Порог ощущения кожных рецепторов при надавливании: 10

Слайд 72Адаптация
Адаптация

Слайд 73Передача импульсации
Передача тактильных ощущений тонкой дифференцировки происходит по миелинизированным нервным волокнам со
Передача импульсации
Передача тактильных ощущений тонкой дифференцировки происходит по миелинизированным нервным волокнам со

Слайд 74Пути передачи соматосенсорных сигналов
Практически вся сенсорная информация от сегментов тела поступает в
Пути передачи соматосенсорных сигналов
Практически вся сенсорная информация от сегментов тела поступает в
Слайд 75Войдя в спинной мозг они направляются:
- либо прямо к продолговатому мозгу (лемнисковая
Войдя в спинной мозг они направляются:
- либо прямо к продолговатому мозгу (лемнисковая
 Размножение ползучими побегами
Размножение ползучими побегами Цветоводство : виды и значение
Цветоводство : виды и значение Грибы. Вешенка обыкновенная
Грибы. Вешенка обыкновенная Ракообразные (жабродышащие)
Ракообразные (жабродышащие) Органы женского малого таза
Органы женского малого таза Китова акула
Китова акула Анатомия центральной нервной системы
Анатомия центральной нервной системы Учёт зимующих птиц в бухтах Севастополя
Учёт зимующих птиц в бухтах Севастополя Пластиды
Пластиды Химический состав клетки. 9 класс
Химический состав клетки. 9 класс Транспорт веществ в организме человека. Диффузия. Осмос
Транспорт веществ в организме человека. Диффузия. Осмос Деление клетки. Митоз
Деление клетки. Митоз Қияр өсіруді бақылау
Қияр өсіруді бақылау Рептилии Саратовской области
Рептилии Саратовской области Игры без правил или плохие новости из белковой упаковки
Игры без правил или плохие новости из белковой упаковки Периферическая нервная система. Черепные нервы человека
Периферическая нервная система. Черепные нервы человека Современное представление о механизмах памяти
Современное представление о механизмах памяти Животные спрятались
Животные спрятались Растительные ткани
Растительные ткани Нервная ткань
Нервная ткань Анатомия и физиология системы кровообращения. Лекция 7
Анатомия и физиология системы кровообращения. Лекция 7 Презентация по биологии на тему _Водоросли_ (7 класс)
Презентация по биологии на тему _Водоросли_ (7 класс) Презентация на тему Эволюция млекопитающих
Презентация на тему Эволюция млекопитающих  Йод, Кальций, Витамин А и Витамин К в жизни человека
Йод, Кальций, Витамин А и Витамин К в жизни человека Систематика типа Хордовые
Систематика типа Хордовые Тканевый инжиниринг
Тканевый инжиниринг Способности
Способности Как растет живое
Как растет живое