- Физиология слухового и вестибулярного анализаторов

Содержание
- 2. С помощью слухового анализатора человек и животные способны ориентироваться во внешней среде, на основе фонорецепции возможны
- 3. Таким образом, первой характеристикой звука, которая оценивается слуховым анализатором, является частота. Частота 1-3 кГц является оптимальной
- 4. Во внутреннем ухе человека расположена улитка, имеющая 2,5 оборота. Именно в среднем канале (или лестнице) на
- 5. Анализ звуков осуществляется как частотой, так и местом локализации фонорецепторов на базиллярной мембране, а также количеством
- 6. Вестибулярный анализатор. Вестибулярный анализатор обеспечивает акселерационное чувство, чувство равновесия, участвует в поддержании позы и вовлечен в
- 7. +По современным представлениям, киноцилия и стереоцилии имеют связи в виде поперечных белковых мостиков, выполняющих роль пружинного
- 9. Скачать презентацию
Слайд 2С помощью слухового анализатора человек и животные способны ориентироваться во внешней среде,
С помощью слухового анализатора человек и животные способны ориентироваться во внешней среде,

Слайд 3Таким образом, первой характеристикой звука, которая оценивается слуховым анализатором, является частота. Частота
Таким образом, первой характеристикой звука, которая оценивается слуховым анализатором, является частота. Частота

Слайд 4Во внутреннем ухе человека расположена улитка, имеющая 2,5 оборота. Именно в среднем
Во внутреннем ухе человека расположена улитка, имеющая 2,5 оборота. Именно в среднем

Слайд 5Анализ звуков осуществляется как частотой, так и местом локализации фонорецепторов на базиллярной
Анализ звуков осуществляется как частотой, так и местом локализации фонорецепторов на базиллярной

Слайд 6Вестибулярный анализатор.
Вестибулярный анализатор обеспечивает акселерационное чувство, чувство равновесия, участвует в поддержании позы
Вестибулярный анализатор.
Вестибулярный анализатор обеспечивает акселерационное чувство, чувство равновесия, участвует в поддержании позы

Слайд 7+По современным представлениям, киноцилия и стереоцилии имеют связи в виде поперечных белковых
+По современным представлениям, киноцилия и стереоцилии имеют связи в виде поперечных белковых

 Высшие споровые растения
Высшие споровые растения Митоз и мейоз
Митоз и мейоз Бычий цепень. Микробиология
Бычий цепень. Микробиология Сердце, сосуды. Фазы работы сердца
Сердце, сосуды. Фазы работы сердца Гуморальная регуляции организма
Гуморальная регуляции организма Роль животных в жизни человека
Роль животных в жизни человека Презентация на тему Какая у нас память
Презентация на тему Какая у нас память  Протисты, эукариотические организмы, не входящие в состав животных, грибов и растений
Протисты, эукариотические организмы, не входящие в состав животных, грибов и растений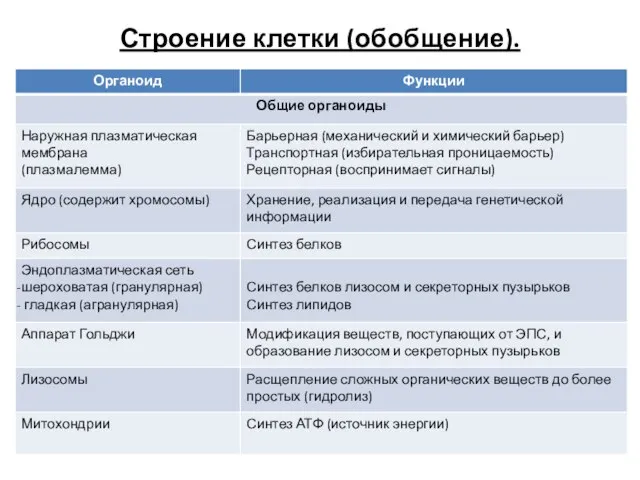 Обмен веществ
Обмен веществ Растения. Царство растения
Растения. Царство растения Головной мозг человека. (Лекция 6)
Головной мозг человека. (Лекция 6) Птицы - наши друзья
Птицы - наши друзья Органы пищеварения человека. Строение и значение зубов
Органы пищеварения человека. Строение и значение зубов Презентация на тему ОСЕНЬ И ЗИМА В ЖИЗНИ РАСТЕНИЙ
Презентация на тему ОСЕНЬ И ЗИМА В ЖИЗНИ РАСТЕНИЙ 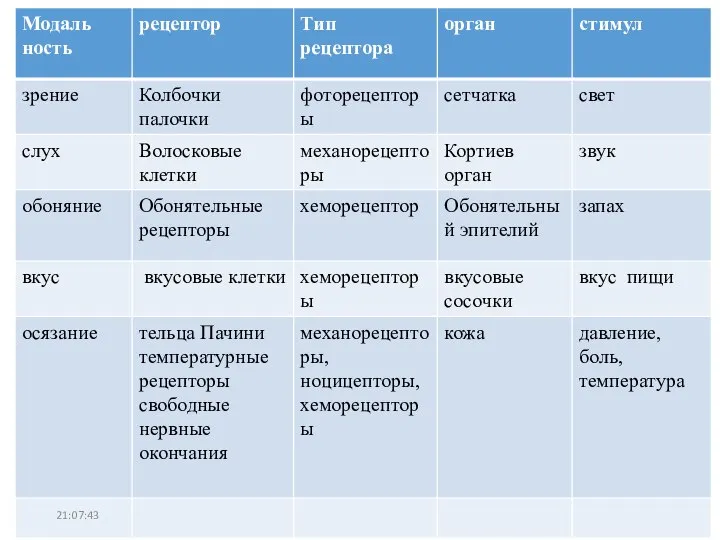 Механорецепторы. Тип рецептора
Механорецепторы. Тип рецептора Строение клетки и функции органоидов. Органеллы клетки
Строение клетки и функции органоидов. Органеллы клетки Удивительные и загадочные Ахатины (Achatina fulica)
Удивительные и загадочные Ахатины (Achatina fulica) Создание гербария Травки-муравки
Создание гербария Травки-муравки Коллекция как проект
Коллекция как проект Пропорции и строение фигуры человека
Пропорции и строение фигуры человека Внутриутробное развитие у млекопитающих. Виды плаценты млекопитающих
Внутриутробное развитие у млекопитающих. Виды плаценты млекопитающих Сеченовка экзамен
Сеченовка экзамен Ткани растений. 5 класс
Ткани растений. 5 класс Фотосинтез
Фотосинтез История генетики. Основные понятия генетики (урок 19)
История генетики. Основные понятия генетики (урок 19) Реципрокті (жинақталған) тежелу
Реципрокті (жинақталған) тежелу Органоиды клетки и их функции (§ 8)
Органоиды клетки и их функции (§ 8) Осьминог. Анотомия и физиология
Осьминог. Анотомия и физиология