- Физиология возбудимых тканей

Содержание
- 9. Диализ — очистка коллоидных растворов и субстанций высокомолекулярных веществ от растворённых в них низкомолекулярных соединений при
- 10. Осмос играет важную роль во многих биологических процессах. Мембрана, окружающая нормальную клетку крови, проницаема лишь для
- 12. Облегченная диффузии, так же, как и простая «диффузия» вещества, осуществляется по градиенту его концентрации, но в
- 15. Активный транспорт веществ через клеточные мембраны — это перенос веществ против их градиентов, осуществляющийся с затратой
- 16. Благодаря активному транспорту могут преодолеваться силы не только концентрационного, но и электрического, электрохимического и других градиентов
- 18. В результате изменения пространственной структуры белка места связывания ионов Na поворачиваются на внешнюю поверхность мембраны. Активность
- 19. Вещество строфантин (уабаин), выделяемое из растения наперстянка, обладает специфической способностью блокировать работу Na+-, К+ — насоса.
- 20. Кроме Na'-, К+-АТФ-азы имеются еще несколько типов транспортных АТФ-аз, или ионных насосов. Среди них насос, осуществляющий
- 21. Например, в клетках скелетных мышц и миокарда белок Са2+ -АТФ-аза встроен в мембраны саркоплазматического ретикулума и
- 22. Вторично-активный транспорт характеризуется тем, что перенос вещества через мембрану осуществляется за счет градиента концентрации другого вещества,
- 23. Из приведенных примеров видно, что вторично-активный транспорт осуществляется за счет использования сил градиента ионов Na+ или
- 24. Эксперименты, положенные в основу мембранной теории, были проведены в 40-х годах ХХ века на гигантских аксонах
- 26. 4.1. Концентрационный и электрический градиенты Для того, чтобы понять, как и почему возникает возбуждение в нервных
- 27. Равновесный потенциал Равновесный потенциал – такое значение трансмембранной разности электрических зарядов, при котором ток ионов в
- 28. Результаты измерений концентраций ионов в разных клетках животных, относящихся к разным видам, дают, разумеется и разные
- 29. Таким образом, если мембрана готова пропустить какие-либо ионы, то направление ионного тока будут определять два обстоятельства:
- 30. У находящихся в состоянии покоя клеток глии мембрана пропускает только ионы калия, поэтому реальная трансмембранная разность
- 31. 4.2. Активный транспорт Диффузия ионов должна уменьшать концентрационный градиент, но концентрационное равновесие означало бы для клетки
- 33. Таким образом, за один цикл работы насоса выносятся из клетки три иона натрия, вносятся в неё
- 34. Создаваемые работой насосов градиенты дают ионам возможность перемещаться через мембрану от большего энергетического уровня к меньшему
- 35. Производительность натрий-калиевого насоса зависит от концентрации в клетке ионов натрия: чем больше она, тем быстрее работает
- 36. 4.4. Управляемые каналы Только что описанные каналы, через которые проходят ионы калия, открыты всегда: и в
- 37. Во-вторых, пониманию свойств каналов способствовало построение их моделей на основе расшифрованного генетического кода многих канальных белков
- 38. Потенциалзависимые каналы Потенциалзависимые каналы открываются или закрываются в связи с определёнными изменениями мембранного потенциала. Например, натриевые
- 39. Хемозависимые каналы Хемозависимые каналы открываются вследствие присоединения нейромедиатора к выступающей наружу гликопротеиновой рецепторной области канального белка
- 41. Механически управляемые каналы Механически управляемые каналы характерны для чувствительных окончаний нейронов, реагирующих на растяжение и давление.
- 42. Во время инактивации каналов клетка перестаёт отвечать на возбуждающие её стимулы, что определяется термином рефрактерность, т.е.
- 44. 4.5. Блокаторы ионных каналов Существует довольно много веществ, способных обратимо или необратимо связываться с молекулами канальных
- 45. Так, например, во внутренностях некоторых сростночелюстных рыб (Tetrodontiformes) содержится тетродотоксин, блокирующий натриевые каналы. К этому отряду
- 46. . Натриевые каналы может блокировать и другой животный яд – батрахотоксин, который содержится в слизи некоторых
- 47. Другие южноамериканские индейцы готовили отравленные стрелы с помощью другого яда, растительного – это древесный сок кураре,
- 49. Скачать презентацию


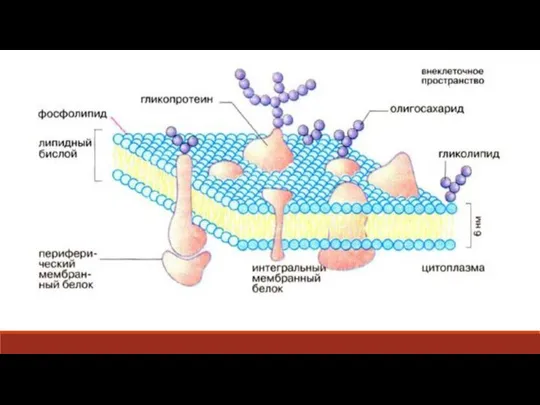



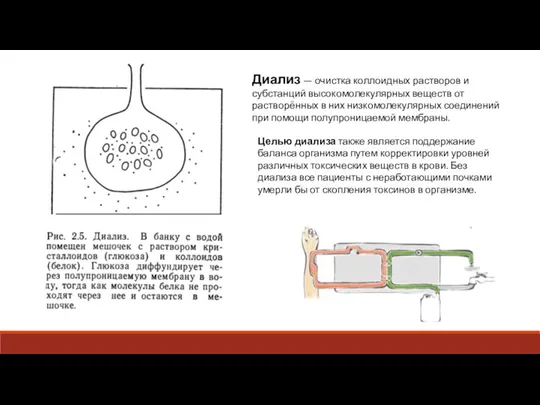
Слайд 9Диализ — очистка коллоидных растворов и субстанций высокомолекулярных веществ от растворённых в
Диализ — очистка коллоидных растворов и субстанций высокомолекулярных веществ от растворённых в
Слайд 10Осмос играет важную роль во многих биологических процессах. Мембрана, окружающая нормальную клетку
Осмос играет важную роль во многих биологических процессах. Мембрана, окружающая нормальную клетку


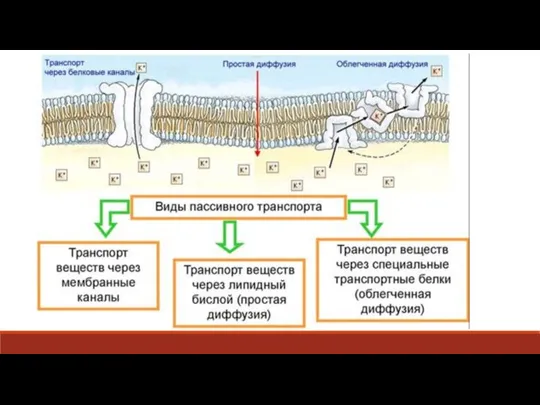
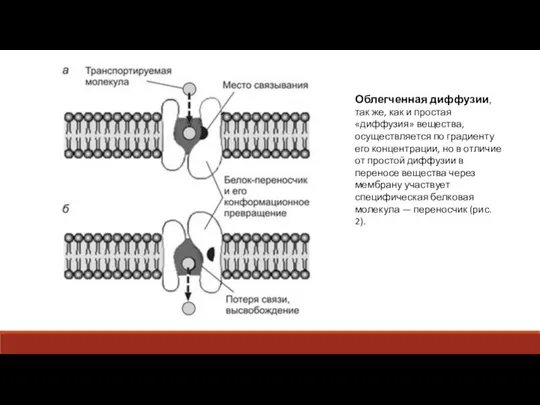
Слайд 12Облегченная диффузии, так же, как и простая «диффузия» вещества, осуществляется по градиенту
Облегченная диффузии, так же, как и простая «диффузия» вещества, осуществляется по градиенту


Слайд 15Активный транспорт веществ через клеточные мембраны — это перенос веществ против их
Активный транспорт веществ через клеточные мембраны — это перенос веществ против их

Слайд 16Благодаря активному транспорту могут преодолеваться силы не только концентрационного, но и электрического,
Благодаря активному транспорту могут преодолеваться силы не только концентрационного, но и электрического,

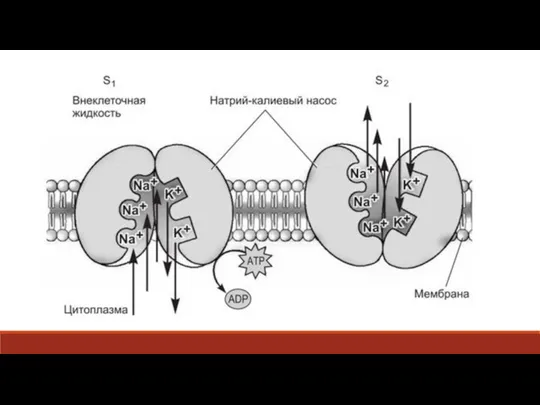
Слайд 18В результате изменения пространственной структуры белка места связывания ионов Na поворачиваются на
В результате изменения пространственной структуры белка места связывания ионов Na поворачиваются на

Слайд 19Вещество строфантин (уабаин), выделяемое из растения наперстянка, обладает специфической способностью блокировать работу
Вещество строфантин (уабаин), выделяемое из растения наперстянка, обладает специфической способностью блокировать работу

Слайд 20Кроме Na'-, К+-АТФ-азы имеются еще несколько типов транспортных АТФ-аз, или ионных насосов.
Кроме Na'-, К+-АТФ-азы имеются еще несколько типов транспортных АТФ-аз, или ионных насосов.
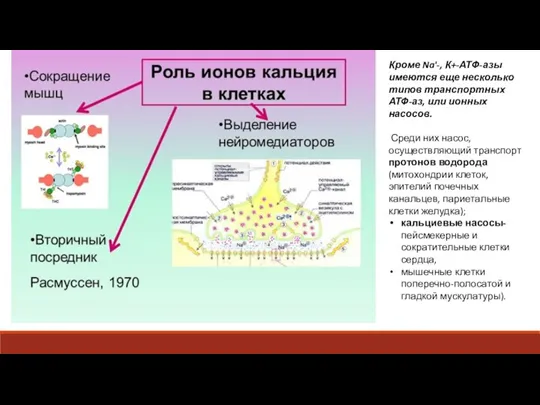
Слайд 21Например, в клетках скелетных мышц и миокарда белок Са2+ -АТФ-аза встроен в
Например, в клетках скелетных мышц и миокарда белок Са2+ -АТФ-аза встроен в
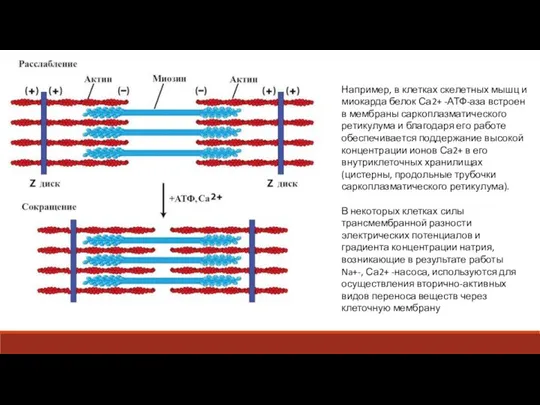
Слайд 22Вторично-активный транспорт характеризуется тем, что перенос вещества через мембрану осуществляется за счет
Вторично-активный транспорт характеризуется тем, что перенос вещества через мембрану осуществляется за счет

Слайд 23Из приведенных примеров видно, что вторично-активный транспорт осуществляется за счет использования сил
Из приведенных примеров видно, что вторично-активный транспорт осуществляется за счет использования сил

Слайд 24Эксперименты, положенные в основу мембранной теории, были проведены в 40-х годах ХХ
Эксперименты, положенные в основу мембранной теории, были проведены в 40-х годах ХХ

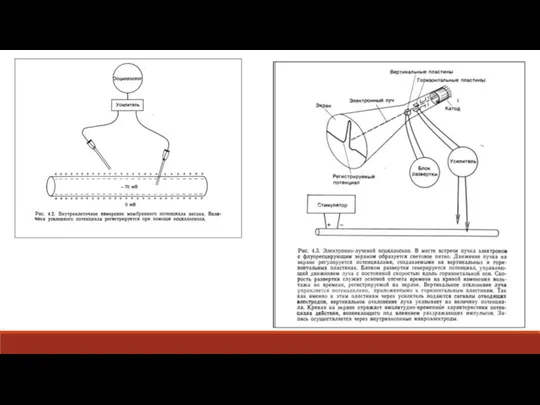
Слайд 264.1. Концентрационный и электрический градиенты
Для того, чтобы понять, как и почему возникает
4.1. Концентрационный и электрический градиенты
Для того, чтобы понять, как и почему возникает

Слайд 27Равновесный потенциал
Равновесный потенциал – такое значение трансмембранной разности электрических зарядов, при
Равновесный потенциал
Равновесный потенциал – такое значение трансмембранной разности электрических зарядов, при

Слайд 28Результаты измерений концентраций ионов в разных клетках животных, относящихся к разным видам,
Результаты измерений концентраций ионов в разных клетках животных, относящихся к разным видам,

Слайд 29 Таким образом, если мембрана готова пропустить какие-либо ионы, то направление ионного
Таким образом, если мембрана готова пропустить какие-либо ионы, то направление ионного

Слайд 30У находящихся в состоянии покоя клеток глии мембрана пропускает только ионы калия,
У находящихся в состоянии покоя клеток глии мембрана пропускает только ионы калия,

Слайд 314.2. Активный транспорт
Диффузия ионов должна уменьшать концентрационный градиент, но концентрационное равновесие
4.2. Активный транспорт
Диффузия ионов должна уменьшать концентрационный градиент, но концентрационное равновесие

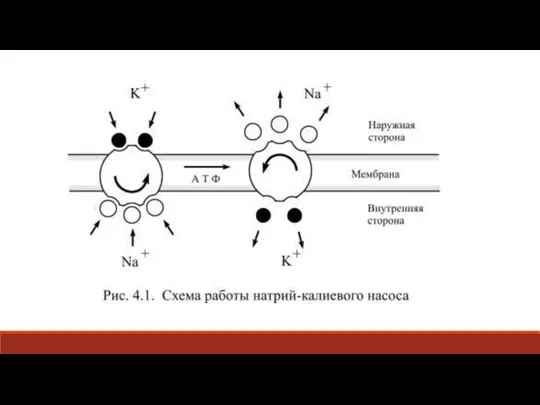
Слайд 33Таким образом, за один цикл работы насоса выносятся из клетки три иона
Таким образом, за один цикл работы насоса выносятся из клетки три иона

Слайд 34Создаваемые работой насосов градиенты дают ионам возможность перемещаться через мембрану от большего
Создаваемые работой насосов градиенты дают ионам возможность перемещаться через мембрану от большего

Слайд 35Производительность натрий-калиевого насоса зависит от концентрации в клетке ионов натрия: чем больше
Производительность натрий-калиевого насоса зависит от концентрации в клетке ионов натрия: чем больше

Слайд 364.4. Управляемые каналы
Только что описанные каналы, через которые проходят ионы калия,
4.4. Управляемые каналы
Только что описанные каналы, через которые проходят ионы калия,

Слайд 37Во-вторых, пониманию свойств каналов способствовало построение их моделей на основе расшифрованного генетического
Во-вторых, пониманию свойств каналов способствовало построение их моделей на основе расшифрованного генетического

Слайд 38Потенциалзависимые каналы
Потенциалзависимые каналы открываются или закрываются в связи с определёнными изменениями
Потенциалзависимые каналы
Потенциалзависимые каналы открываются или закрываются в связи с определёнными изменениями

Слайд 39Хемозависимые каналы
Хемозависимые каналы открываются вследствие присоединения нейромедиатора к выступающей наружу гликопротеиновой
Хемозависимые каналы
Хемозависимые каналы открываются вследствие присоединения нейромедиатора к выступающей наружу гликопротеиновой


Слайд 41Механически управляемые каналы
Механически управляемые каналы характерны для чувствительных окончаний нейронов, реагирующих
Механически управляемые каналы
Механически управляемые каналы характерны для чувствительных окончаний нейронов, реагирующих

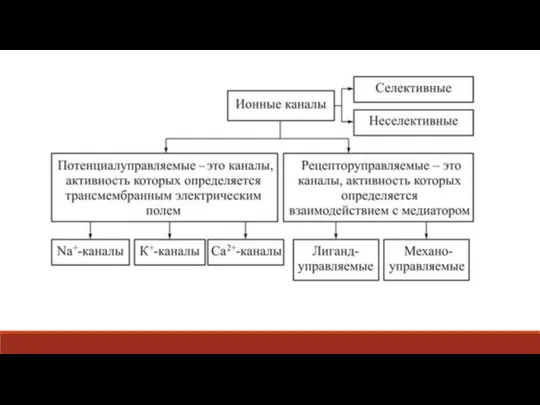
Слайд 42Во время инактивации каналов клетка перестаёт отвечать на возбуждающие её стимулы, что
Во время инактивации каналов клетка перестаёт отвечать на возбуждающие её стимулы, что

Слайд 444.5. Блокаторы ионных каналов
Существует довольно много веществ, способных обратимо или необратимо связываться
4.5. Блокаторы ионных каналов
Существует довольно много веществ, способных обратимо или необратимо связываться

Слайд 45Так, например, во внутренностях некоторых сростночелюстных рыб (Tetrodontiformes) содержится тетродотоксин, блокирующий натриевые
Так, например, во внутренностях некоторых сростночелюстных рыб (Tetrodontiformes) содержится тетродотоксин, блокирующий натриевые

Слайд 46. Натриевые каналы может блокировать и другой животный яд – батрахотоксин, который
. Натриевые каналы может блокировать и другой животный яд – батрахотоксин, который

Слайд 47Другие южноамериканские индейцы готовили отравленные стрелы с помощью другого яда, растительного –
Другие южноамериканские индейцы готовили отравленные стрелы с помощью другого яда, растительного –

 Презентация на тему "Генетический тест" - презентации по Биологии
Презентация на тему "Генетический тест" - презентации по Биологии Круглые черви (нематоды)
Круглые черви (нематоды) Человек и животные. Домашние животные
Человек и животные. Домашние животные Птицы России
Птицы России Птицы Республики Коми
Птицы Республики Коми Минеральное питание растений
Минеральное питание растений Корни.Корневые системы
Корни.Корневые системы Проращивание овса
Проращивание овса Типы взаимодействия между популяциями
Типы взаимодействия между популяциями Фауна родного края
Фауна родного края Эволюция огранов дыхания
Эволюция огранов дыхания Бактерии – разрушители органических веществ
Бактерии – разрушители органических веществ Нервная система. Работа с гистологическими препаратами
Нервная система. Работа с гистологическими препаратами Жизнь в пресных водах
Жизнь в пресных водах Чому людина не може жити без рослин
Чому людина не може жити без рослин Водоросли. Свечение моря
Водоросли. Свечение моря Методы диагностики в бактериологии
Методы диагностики в бактериологии Микробалдырлар биотехнологиясы
Микробалдырлар биотехнологиясы Секреты растительной клетки. Почему желтеют листья?
Секреты растительной клетки. Почему желтеют листья? Отчет по летней практике. Приготовление среды Мурасиге-Скуга. Микроклональное размножение
Отчет по летней практике. Приготовление среды Мурасиге-Скуга. Микроклональное размножение Khimicheskiy_sostav_kletki_11_09
Khimicheskiy_sostav_kletki_11_09 Городской парк
Городской парк Крахмал и его свойства. Исследовательская работа
Крахмал и его свойства. Исследовательская работа Морские жители
Морские жители Отдел Папоротникообразные
Отдел Папоротникообразные Конкурентное и неконкурентное ингибирование ферментов
Конкурентное и неконкурентное ингибирование ферментов Закономерности изменчивости. Модификационная изменчивость. Норма реакции
Закономерности изменчивости. Модификационная изменчивость. Норма реакции Презентация на тему МНОГООБРАЗИЕ НАСЕКОМЫХ
Презентация на тему МНОГООБРАЗИЕ НАСЕКОМЫХ