- Физиология возбудимых тканей. Биологические реакции организма

Содержание
- 2. Способность АДАПТИРОВАТЬСЯ к постоянно изменяющимся условиям внешней среды является одним из основных признаков живых систем
- 3. ОСНОВА ВСЕХ ФИЗИОЛОГИЧЕСКИХ РЕАКЦИЙ ОРГАНИЗМА – СПОСОБНОСТЬ ЖИВОЙ КЛЕТКИ РЕАГИРОВАТЬ НА РАЗДРАЖИТЕЛЬ
- 4. Все клетки и ткани живых систем в процессе эволюции приобрели способность отвечать на действие раздражителя специфической
- 5. В основе приспособительных (адаптационных) реакций организма лежит РАЗДРАЖИМОСТЬ. Под термином «раздражимость» понимают способность реагировать на действие
- 6. ВОЗБУЖДЕНИЕ – это сложный физиологический процесс временной деполяризации мембраны клеток, который проявляется специализированной реакцией ткани: проведение
- 7. ВОЗБУДИМОСТЬ различных тканей неодинакова. Величину ВОЗБУДИМОСТИ оценивают по порогу раздражения – минимальной силе раздражителя, кт способна
- 8. Раздражителями, вызывающими ВОЗБУЖДЕНИЕ, могут быть: любые внешние (действующие из окружающей среды) или внутренние (возникающие в самом
- 9. По ХАРАКТЕРУ ВОЗДЕЙСТВИЯ раздражители можно разделить на 3-и группы: ФИЗИЧЕСКИЕ: Механические Электрические Температурные Звуковые Световые ХИМИЧЕСКИЕ:
- 10. По СТЕПЕНИ ПРИСПОСОБЛЕННОСТИ биологических структур к восприятию раздражителя – раздражители можно разделить на: Адекватные Неадекватные
- 11. АДЕКВАТНЫМИ называются раздражители, к восприятию которых биологическая структура специально приспособлена в процессе эволюции. Например: адекватным раздражителем
- 12. НЕАДЕКВАТНЫМИ называются такие раздражители, которые действуют на структуру, специально не приспособленную для их восприятия. Например: мышца
- 13. Разделы Лекции: Строение и функции клеточной мембраны Биоэлектрические явления в возбудимых тканях Изменения мембранного потенциала. Потенциал
- 14. 1. СТРОЕНИЕ И ФУНКЦИИ КЛЕТОЧНОЙ МЕМБРАНЫ
- 15. Хотя в ПРОЦЕССЕ ВОЗБУЖДЕНИЯ участвуют все органеллы клетки, но главным местом этого процесса является КЛЕТОЧНАЯ МЕМБРАНА
- 16. СТРОЕНИЕ КЛЕТОЧНОЙ МЕМБРАНЫ (обеспечивает полупроницаемость):
- 17. СПОСОБЫ ПЕРЕНОСА ВЕЩЕСТВ ЧЕРЕЗ МЕМБРАНУ: пассивный транспорт активный транспорт, а также транспорт, связанный с нарушением структурной
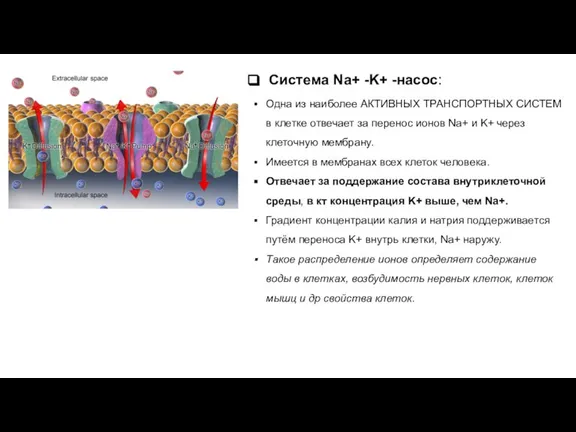
- 18. Система Na+ -K+ -насос: Одна из наиболее АКТИВНЫХ ТРАНСПОРТНЫХ СИСТЕМ в клетке отвечает за перенос ионов
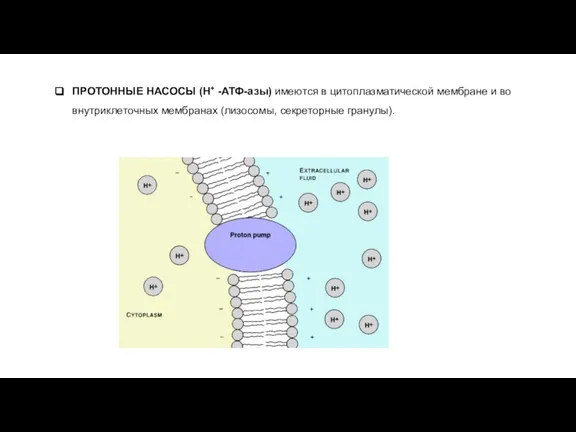
- 19. ПРОТОННЫЕ НАСОСЫ (H+ -АТФ-азы) имеются в цитоплазматической мембране и во внутриклеточных мембранах (лизосомы, секреторные гранулы).
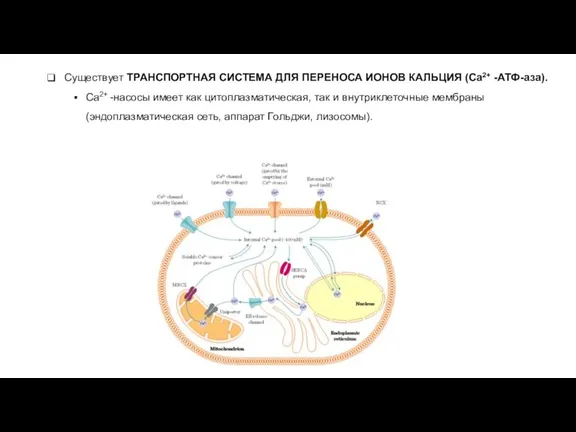
- 20. Существует ТРАНСПОРТНАЯ СИСТЕМА ДЛЯ ПЕРЕНОСА ИОНОВ КАЛЬЦИЯ (Ca2+ -АТФ-аза). Ca2+ -насосы имеет как цитоплазматическая, так и
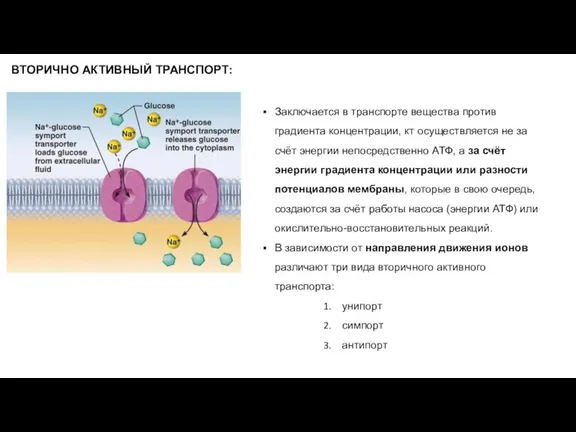
- 21. Заключается в транспорте вещества против градиента концентрации, кт осуществляется не за счёт энергии непосредственно АТФ, а
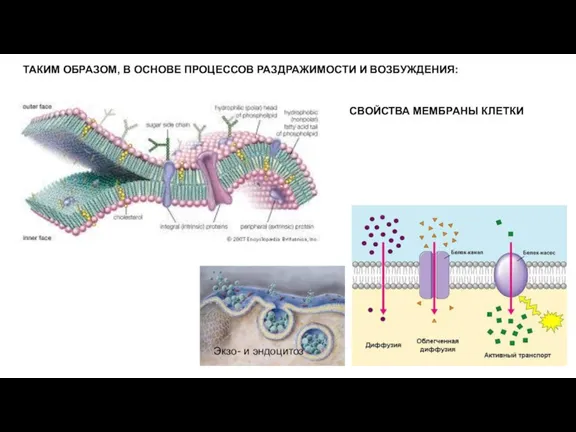
- 22. ТАКИМ ОБРАЗОМ, В ОСНОВЕ ПРОЦЕССОВ РАЗДРАЖИМОСТИ И ВОЗБУЖДЕНИЯ: Экзо- и эндоцитоз СВОЙСТВА МЕМБРАНЫ КЛЕТКИ
- 23. 2. БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ В ВОЗБУДИМЫХ ТКАНЯХ. МЕМБРАННЫЙ ПОТЕНЦИАЛ (МП).
- 24. ВОЗБУЖДЕНИЕ представляет собой сложную совокупность физических, химических и физико-химических процессов, в результате которых происходит быстрое и
- 25. Первые исследования электрической активности живых тканей были проведены Гальвани. Он обратил внимание на сокращение препарата задних
- 26. Физик Вольта, повторив этот опыт, пришёл к другому заключению. Источником тока, по его мнению, являются не
- 27. В ответ Гальвани усовершенствовал опыт, исключив из него металлы. Он препарировал седалищный нерв вдоль бедра лапки
- 28. Позднее Дюбуа-Реймоном было установлено, что повреждённый участок мышцы имеет отрицательный заряд, а неповреждённый участок – положительный.
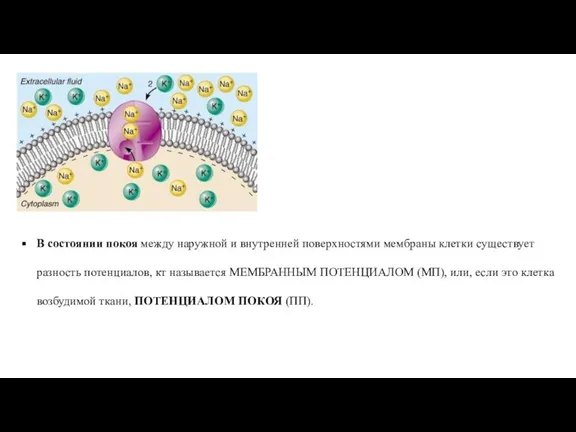
- 29. В состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность потенциалов, кт называется МЕМБРАННЫМ
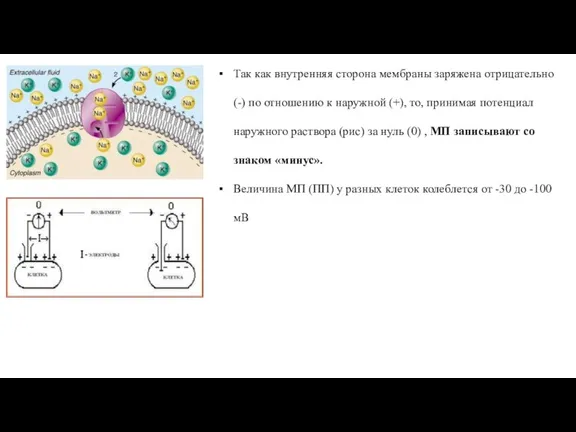
- 30. Так как внутренняя сторона мембраны заряжена отрицательно (-) по отношению к наружной (+), то, принимая потенциал
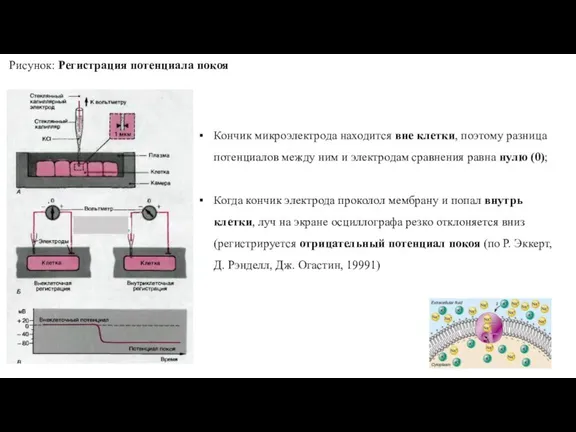
- 31. Рисунок: Регистрация потенциала покоя Кончик микроэлектрода находится вне клетки, поэтому разница потенциалов между ним и электродам
- 32. Первая теория возникновения и поддержания мембранного потенциала была разработана Ю. Бернштейном (1902). Исходя из того, что
- 33. А. Ходжкин, Э. Хаксли, Б. Катц – создали современную мембранно-ионную теорию (1949-1952) : Согласно этой теории
- 34. Цитоплазма нервных и мышечных клеток по сравнению с внеклеточной жидкостью содержит: в 30-50 раз больше ионов
- 35. Проницаемость мембраны для ионов обусловлена ИОННЫМИ КАНАЛАМИ, макромолекулами белка, пронизывающими липидный слой: одни каналы открыты постоянно,
- 36. Потенциалзависимые каналы подразделятся на: Натриевые, Калиевые, Хлорные. Например: В состоянии физиологического покоя проницаемость мембраны нервных клеток
- 37. ТАКИМ ОБРАЗОМ, согласно обновлённой мембранной теории, возникновение и поддержание МП (ПП) обусловлено: Избирательной проницаемостью мембраны для
- 38. Поэтому более точно величину мембранного потенциала можно рассчитать по формуле (обновлённая формула Нернста): Eм – разность
- 39. Поляризация мембраны в покое объяснятся наличием открытых К-каналов и трансмембранным градиентом концентраций К, что приводит выходу
- 40. Переход ионов K и Na через мембрану по их концентрационному градиенту в конечном итоге должен был
- 41. Таким образом: возникновение и поддержание МП (ПП) обусловлено не только избирательной проницаемостью мембраны клетки, но и
- 42. 3. ИЗМЕНЕНИЯ МЕМБРАННОГО ПОТЕНЦИАЛА (МП/ПП). ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД). ПОРОГОВЫЕ И ПОДПОРОГОВЫЕ РАЗДРАЖИТЕЛИ.
- 43. Если раздражать нейрон через электрод, находящийся в цитоплазме, кратковременными импульсами деполяризующего электрического тока различной величины, то,
- 44. ЭЛЕКТРОТОНИЧЕСКИЙ ПОТЕНЦИАЛ: Если наносятся раздражения, величина которых не превышает 0.5 величины порогового раздражения, то деполяризация мембраны
- 45. ЛОКАЛЬНЫЙ ОТВЕТ: При увеличении амплитуды подпороговых раздражений от 0.5 до 0.9 пороговой величины развитие деполяризации мембраны
- 46. СВОЙСТВА ЛОКАЛЬНОГО ОТВЕТА: Возникает при действии подпороговых раздражителей; Находится в градуальной зависимости от силы стимула (не
- 47. В период развития локального ответа увеличивается поток ионов Na в клетку, что повышает её возбудимость. Локальный
- 48. ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД): ПД возникает на мембранах возбудимых клеток под влиянием раздражителя пороговой или сверхпороговой величины,
- 49. Ионы Na начинают входить внутрь клетки, что приводит к уменьшению величины МП – деполяризации мембраны. При
- 50. В результате проникновения ионов Na в цитоплазму и их взаимодействия с анионами разность потенциалов на мембране
- 51. В результате выхода K из клетки начинается процесс восстановления исходного уровня МП (ПП) – реполяризация мембраны.
- 52. Например: Если такое повышение проводимости для K предотвратить введением тетраэтиламмония, который избирательно блокирует K-каналы, мембрана реполяризуется
- 53. Таким образом: в основе ВОЗБУЖДЕНИЯ (генерации ПД) лежит повышение проницаемости мембраны для ионов Na, за счёт
- 54. В потенциале действия (ПД) различают следующие фазы: Предспайк – процесс медленной деполяризации мембраны (ДП) до критического
- 55. ПОТЕНЦИАЛ ДЕЙСТВИЯ В РАЗЛИЧНЫХ ТИПАХ КЛЕТОК ВОЗБУДИМЫХ ТКАНЕЙ
- 56. 4. ИЗМЕНЕНИЕ ВОЗБУДИМОСТИ ПРИ ВОЗБУЖДЕНИИ
- 57. ПРИ РАЗВИТИИ ПД ПРОИСХОДЯТ ФАЗНЫЕ ИЗМЕНЕНИЯ ВОЗБУДИМОСТИ ТКАНЕЙ: Фаза нормального уровня возбудимости Фаза повышенной возбудимости (первичной
- 58. 1. Фаза. Состоянию исходной поляризации мембраны (МП/ПП) соответствует нормальный уровень возбудимости (НВ). 2. Фаза. В период
- 59. 3. Фаза. В период развития спайка (пикового потенциала) идёт лавинообразное поступление ионов Na внутрь клетки, в
- 60. 4. Фаза. После окончания фазы перезарядки мембраны возбудимость её постепенно восстанавливается до исходного уровня – фаза
- 61. 5. Фаза. Периоду отрицательного следового потенциала соответствует повышенный уровень возбудимости (фаза вторичной экзальтации/ЭК). Так как МП
- 62. 6. Фаза. В период развития положительного следового потенциала возбудимость ткани понижена – фаза субнормальной возбудимости (вторичной
- 63. Таким образом:
- 64. Рефрактерность мембраны является следствием того, что Na-канал состоит из собственного канала (транспортной части) и воротного механизма,
- 65. «Ворота» могут быть полностью открыты или закрыты: например, в Na-канале в состоянии покоя «ворота» m закрыты,
- 66. Следовая гиперполяризация мембраны может возникать вследствие трёх причин: продолжающимся выходом ионов K; открытием каналов для ионов
- 67. 5. ЗАКОНЫ РАЗДРАЖЕНИЯ ВОЗБУДИМЫХ ТКАНЕЙ
- 68. Характер ответных реакций отдельных возбудимых структур и организма в целом зависит от: силы, длительности, скорости нарастания
- 69. ЗАКОНЫ раздражения возбудимых тканей отражают различные зависимости между действием раздражителя и характером ответной реакции возбудимой ткани.
- 70. Ответная реакция возбудимой ткани зависит от силы раздражения: При действии слабых (подпороговых) раздражителей ответной реакции не
- 71. ЗАКОНЫ раздражения возбудимых тканей: Закон силы Закон «всё или ничего» Закон силы-длительности Закон раздражения (аккомодации) Дюбуа-Реймона
- 72. 1. ЗАКОН СИЛЫ: Чем больше сила раздражителя, тем сильнее ответная реакция (как целого организма, так изолированного
- 73. Это обусловлено тем, что скелетная мышца состоит из множества мышечных волокон, имеющих различную возбудимость. На пороговые
- 74. 2. ЗАКОН «ВСЁ ИЛИ НИЧЕГО»: В физиологии существует представление о том, что ряд структур не подчиняется
- 75. Закон «всё или ничего» относителен, так как хотя на раздражители подпороговой силы и не возникает видимой
- 76. 3. ЗАКОН СИЛЫ-ДЛИТЕЛЬНОСТИ: В начале XX века интенсивно изучался вопрос о соотношении силы и длительности раздражения.
- 77. Исследования зависимости силы-длительности показали, что она имеет гиперболический характер. Ток меньше некоторой минимальной величины не вызывает
- 78. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой. Время, в течение
- 79. Хронаксиметрия (от греч. chrons – время, axia – цена, metron – мера) – метод изучения возбудимости
- 80. В клинической практике метод хронаксиметрии можно применять для определения степени снижения возбудимости нервной и мышечной ткани
- 81. 4. ЗАКОН РАЗДРАЖЕНИЯ (аккомодации) Дюбуа-Реймона: Стимулирующее действие постоянного тока зависит не только от абсолютной величины силы
- 82. При снижении скорости нарастания силы раздражителя ПД не возникает, тк деполяризация мембраны является пусковым стимулом к
- 83. При быстром нарастании тока повышение Na-проницаемости успевает достичь значительной величины, прежде чем наступит инактивация Na-проницаемости. При
- 84. Способность к аккомодации различных структур неодинакова: Наиболее высокая у двигательных нервных волокон; Значительно меньше – у
- 85. 5. ЗАКОН ПОЛЯРНОГО ДЕЙСТВИЯ ПОСТОЯННОГО ТОКА: При замыкании тока возбуждение возникает под катодом, а при размыкании
- 86. 6. ЗАКОН ФИЗИОЛОГИЧЕСКОГО ЭЛЕКТРОТОНА: Действие постоянного тока на ткань сопровождается изменением её возбудимости. При прохождении постоянного
- 87. Эти изменения возбудимости под катодом и анодом получили название электротона (электротоническое изменение возбудимости): повышение возбудимости под
- 88. При дальнейшем действии постоянного тока: первоначальное повышение возбудимости под катодом сменяется её понижением, развивается т.н. катодическая
- 89. 6. ОСНОВНЫЕ ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА ВОЗБУДИМЫХ ТКАНЕЙ: Возбудимость Проводимость Рефрактерность (невозбудимость) Лабильность (функциональная подвижность) Сократимость
- 90. Возбудимость – способность ткани отвечать на раздражение генерацией процесса возбуждения. Возбудимость зависит от уровня обменных процессов
- 91. 2. Проводимость – способность ткани проводить возбуждение по всей своей длине. Показатель проводимости – скорость проведения
- 92. 3. Рефрактерность (невозбудимость) - способность ткани резко снижать свою возбудимость при возбуждении. В момент самой активной
- 93. 4. Лабильность (функциональная подвижность) - способность ткани воспроизводить определенное число волн возбуждения в единицу времени в
- 94. 5. Сократимость – только для мышечной ткани.
- 95. ФИЗИОЛОГИЮ ВОЗБУДИМЫХ ТКАНЕЙ изучает электрофизиология – раздел физиологии, который исследует электрические проявления жизнедеятельности клеток, тканей и
- 97. Скачать презентацию
Слайд 2Способность АДАПТИРОВАТЬСЯ
к постоянно изменяющимся условиям внешней среды
является
одним из основных
Способность АДАПТИРОВАТЬСЯ
к постоянно изменяющимся условиям внешней среды
является
одним из основных

Слайд 3ОСНОВА
ВСЕХ ФИЗИОЛОГИЧЕСКИХ РЕАКЦИЙ ОРГАНИЗМА –
СПОСОБНОСТЬ
ЖИВОЙ КЛЕТКИ
РЕАГИРОВАТЬ НА РАЗДРАЖИТЕЛЬ
ОСНОВА
ВСЕХ ФИЗИОЛОГИЧЕСКИХ РЕАКЦИЙ ОРГАНИЗМА –
СПОСОБНОСТЬ
ЖИВОЙ КЛЕТКИ
РЕАГИРОВАТЬ НА РАЗДРАЖИТЕЛЬ

Слайд 4Все клетки и ткани живых систем в процессе эволюции приобрели способность отвечать
Все клетки и ткани живых систем в процессе эволюции приобрели способность отвечать
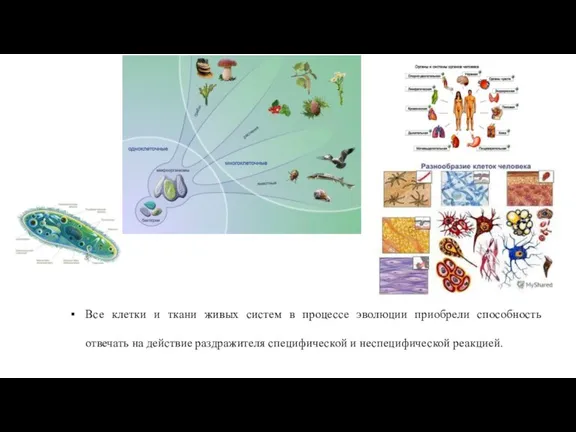
Слайд 5В основе приспособительных (адаптационных) реакций организма лежит РАЗДРАЖИМОСТЬ.
Под термином «раздражимость» понимают способность
В основе приспособительных (адаптационных) реакций организма лежит РАЗДРАЖИМОСТЬ.
Под термином «раздражимость» понимают способность

Слайд 6ВОЗБУЖДЕНИЕ – это сложный физиологический процесс временной деполяризации мембраны клеток, который проявляется
ВОЗБУЖДЕНИЕ – это сложный физиологический процесс временной деполяризации мембраны клеток, который проявляется

Слайд 7ВОЗБУДИМОСТЬ различных тканей неодинакова.
Величину ВОЗБУДИМОСТИ оценивают по порогу раздражения – минимальной
ВОЗБУДИМОСТЬ различных тканей неодинакова.
Величину ВОЗБУДИМОСТИ оценивают по порогу раздражения – минимальной

Слайд 8Раздражителями,
вызывающими ВОЗБУЖДЕНИЕ, могут быть:
любые внешние (действующие из окружающей среды) или
внутренние
Раздражителями,
вызывающими ВОЗБУЖДЕНИЕ, могут быть:
любые внешние (действующие из окружающей среды) или
внутренние

Слайд 9По ХАРАКТЕРУ ВОЗДЕЙСТВИЯ раздражители можно разделить на 3-и группы:
ФИЗИЧЕСКИЕ:
Механические
Электрические
Температурные
Звуковые
Световые
ХИМИЧЕСКИЕ:
Щелочи, кислоты, соли
По ХАРАКТЕРУ ВОЗДЕЙСТВИЯ раздражители можно разделить на 3-и группы:
ФИЗИЧЕСКИЕ:
Механические
Электрические
Температурные
Звуковые
Световые
ХИМИЧЕСКИЕ:
Щелочи, кислоты, соли

Слайд 10По СТЕПЕНИ ПРИСПОСОБЛЕННОСТИ биологических структур к восприятию раздражителя – раздражители можно разделить
По СТЕПЕНИ ПРИСПОСОБЛЕННОСТИ биологических структур к восприятию раздражителя – раздражители можно разделить

Слайд 11АДЕКВАТНЫМИ называются раздражители, к восприятию которых биологическая структура специально приспособлена в процессе
АДЕКВАТНЫМИ называются раздражители, к восприятию которых биологическая структура специально приспособлена в процессе

Слайд 12НЕАДЕКВАТНЫМИ называются такие раздражители, которые действуют на структуру, специально не приспособленную для
НЕАДЕКВАТНЫМИ называются такие раздражители, которые действуют на структуру, специально не приспособленную для

Слайд 13Разделы Лекции:
Строение и функции клеточной мембраны
Биоэлектрические явления в возбудимых тканях
Изменения мембранного потенциала.
Разделы Лекции:
Строение и функции клеточной мембраны
Биоэлектрические явления в возбудимых тканях
Изменения мембранного потенциала.

Слайд 141. СТРОЕНИЕ И ФУНКЦИИ КЛЕТОЧНОЙ МЕМБРАНЫ
1. СТРОЕНИЕ И ФУНКЦИИ КЛЕТОЧНОЙ МЕМБРАНЫ

Слайд 15Хотя в ПРОЦЕССЕ ВОЗБУЖДЕНИЯ участвуют все органеллы клетки,
но главным местом этого
Хотя в ПРОЦЕССЕ ВОЗБУЖДЕНИЯ участвуют все органеллы клетки,
но главным местом этого
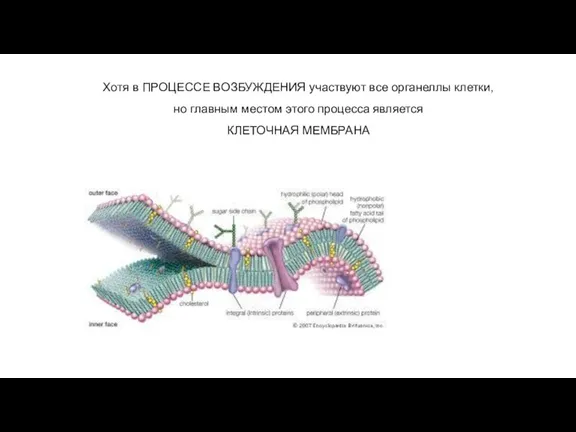
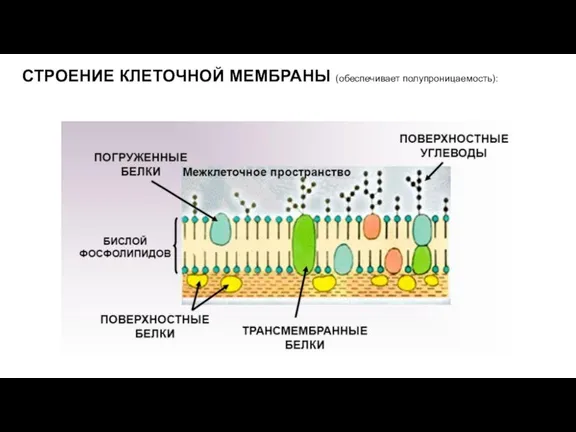
Слайд 16СТРОЕНИЕ КЛЕТОЧНОЙ МЕМБРАНЫ (обеспечивает полупроницаемость):
СТРОЕНИЕ КЛЕТОЧНОЙ МЕМБРАНЫ (обеспечивает полупроницаемость):
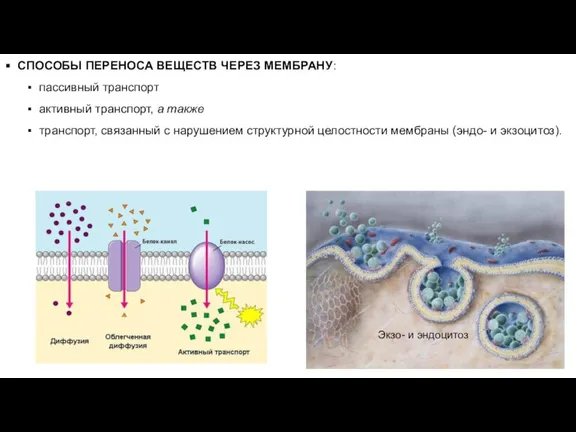
Слайд 17СПОСОБЫ ПЕРЕНОСА ВЕЩЕСТВ ЧЕРЕЗ МЕМБРАНУ:
пассивный транспорт
активный транспорт, а также
транспорт,
СПОСОБЫ ПЕРЕНОСА ВЕЩЕСТВ ЧЕРЕЗ МЕМБРАНУ:
пассивный транспорт
активный транспорт, а также
транспорт,
Слайд 18Система Na+ -K+ -насос:
Одна из наиболее АКТИВНЫХ ТРАНСПОРТНЫХ СИСТЕМ в клетке отвечает
Система Na+ -K+ -насос:
Одна из наиболее АКТИВНЫХ ТРАНСПОРТНЫХ СИСТЕМ в клетке отвечает
Слайд 19ПРОТОННЫЕ НАСОСЫ (H+ -АТФ-азы) имеются в цитоплазматической мембране и во внутриклеточных мембранах
ПРОТОННЫЕ НАСОСЫ (H+ -АТФ-азы) имеются в цитоплазматической мембране и во внутриклеточных мембранах
Слайд 20Существует ТРАНСПОРТНАЯ СИСТЕМА ДЛЯ ПЕРЕНОСА ИОНОВ КАЛЬЦИЯ (Ca2+ -АТФ-аза).
Ca2+ -насосы имеет как
Существует ТРАНСПОРТНАЯ СИСТЕМА ДЛЯ ПЕРЕНОСА ИОНОВ КАЛЬЦИЯ (Ca2+ -АТФ-аза).
Ca2+ -насосы имеет как
Слайд 21Заключается в транспорте вещества против градиента концентрации, кт осуществляется не за счёт
Заключается в транспорте вещества против градиента концентрации, кт осуществляется не за счёт
Слайд 22ТАКИМ ОБРАЗОМ, В ОСНОВЕ ПРОЦЕССОВ РАЗДРАЖИМОСТИ И ВОЗБУЖДЕНИЯ:
Экзо- и эндоцитоз
СВОЙСТВА МЕМБРАНЫ
ТАКИМ ОБРАЗОМ, В ОСНОВЕ ПРОЦЕССОВ РАЗДРАЖИМОСТИ И ВОЗБУЖДЕНИЯ:
Экзо- и эндоцитоз
СВОЙСТВА МЕМБРАНЫ
Слайд 232. БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ В ВОЗБУДИМЫХ ТКАНЯХ.
МЕМБРАННЫЙ ПОТЕНЦИАЛ (МП).
2. БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ В ВОЗБУДИМЫХ ТКАНЯХ.
МЕМБРАННЫЙ ПОТЕНЦИАЛ (МП).

Слайд 24ВОЗБУЖДЕНИЕ
представляет собой сложную совокупность
физических, химических и физико-химических процессов,
в результате
ВОЗБУЖДЕНИЕ
представляет собой сложную совокупность
физических, химических и физико-химических процессов,
в результате

Слайд 25Первые исследования электрической активности живых тканей были проведены Гальвани.
Он обратил внимание
Первые исследования электрической активности живых тканей были проведены Гальвани.
Он обратил внимание

Слайд 26Физик Вольта, повторив этот опыт, пришёл к другому заключению.
Источником тока, по
Физик Вольта, повторив этот опыт, пришёл к другому заключению.
Источником тока, по

Слайд 27В ответ Гальвани усовершенствовал опыт, исключив из него металлы.
Он препарировал седалищный нерв
В ответ Гальвани усовершенствовал опыт, исключив из него металлы.
Он препарировал седалищный нерв

Слайд 28Позднее Дюбуа-Реймоном было установлено, что повреждённый участок мышцы имеет отрицательный заряд, а
Позднее Дюбуа-Реймоном было установлено, что повреждённый участок мышцы имеет отрицательный заряд, а

Слайд 29В состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность
В состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность
Слайд 30Так как внутренняя сторона мембраны заряжена отрицательно (-) по отношению к наружной
Так как внутренняя сторона мембраны заряжена отрицательно (-) по отношению к наружной
Слайд 31Рисунок: Регистрация потенциала покоя
Кончик микроэлектрода находится вне клетки, поэтому разница потенциалов между
Рисунок: Регистрация потенциала покоя
Кончик микроэлектрода находится вне клетки, поэтому разница потенциалов между
Слайд 32Первая теория возникновения и поддержания мембранного потенциала была разработана Ю. Бернштейном (1902).
Исходя
Первая теория возникновения и поддержания мембранного потенциала была разработана Ю. Бернштейном (1902).
Исходя

Слайд 33А. Ходжкин, Э. Хаксли, Б. Катц – создали современную мембранно-ионную теорию (1949-1952)
А. Ходжкин, Э. Хаксли, Б. Катц – создали современную мембранно-ионную теорию (1949-1952)

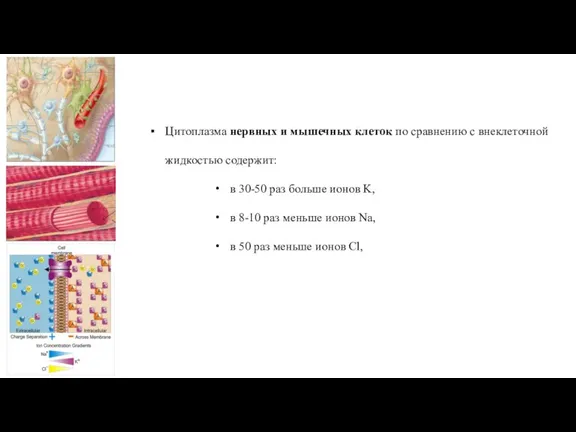
Слайд 34Цитоплазма нервных и мышечных клеток по сравнению с внеклеточной жидкостью содержит:
в 30-50
Цитоплазма нервных и мышечных клеток по сравнению с внеклеточной жидкостью содержит:
в 30-50
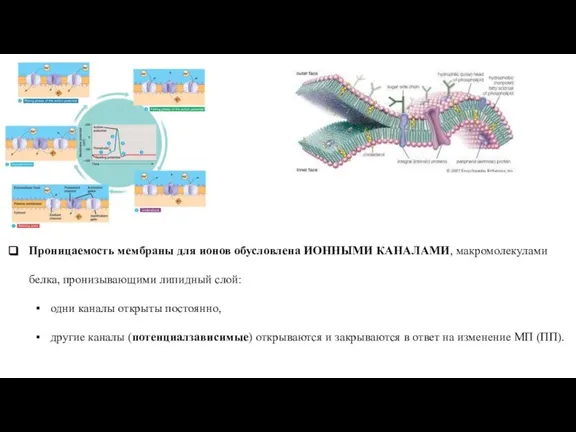
Слайд 35Проницаемость мембраны для ионов обусловлена ИОННЫМИ КАНАЛАМИ, макромолекулами белка, пронизывающими липидный слой:
одни
Проницаемость мембраны для ионов обусловлена ИОННЫМИ КАНАЛАМИ, макромолекулами белка, пронизывающими липидный слой:
одни
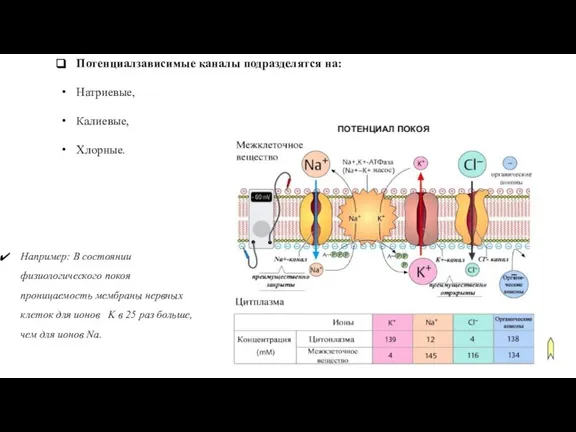
Слайд 36Потенциалзависимые каналы подразделятся на:
Натриевые,
Калиевые,
Хлорные.
Например: В состоянии физиологического покоя проницаемость
Потенциалзависимые каналы подразделятся на:
Натриевые,
Калиевые,
Хлорные.
Например: В состоянии физиологического покоя проницаемость
Слайд 37ТАКИМ ОБРАЗОМ, согласно обновлённой мембранной теории, возникновение и поддержание МП (ПП) обусловлено:
Избирательной
ТАКИМ ОБРАЗОМ, согласно обновлённой мембранной теории, возникновение и поддержание МП (ПП) обусловлено:
Избирательной

Слайд 38Поэтому более точно величину мембранного потенциала можно рассчитать по формуле (обновлённая формула
Поэтому более точно величину мембранного потенциала можно рассчитать по формуле (обновлённая формула

Слайд 39Поляризация мембраны в покое объяснятся наличием открытых К-каналов и трансмембранным градиентом концентраций
Поляризация мембраны в покое объяснятся наличием открытых К-каналов и трансмембранным градиентом концентраций

Слайд 40Переход ионов K и Na через мембрану по их концентрационному градиенту в
Переход ионов K и Na через мембрану по их концентрационному градиенту в

Слайд 41Таким образом:
возникновение и поддержание МП (ПП) обусловлено не только избирательной проницаемостью мембраны
Таким образом:
возникновение и поддержание МП (ПП) обусловлено не только избирательной проницаемостью мембраны

Слайд 423. ИЗМЕНЕНИЯ МЕМБРАННОГО ПОТЕНЦИАЛА (МП/ПП).
ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД).
ПОРОГОВЫЕ И ПОДПОРОГОВЫЕ РАЗДРАЖИТЕЛИ.
3. ИЗМЕНЕНИЯ МЕМБРАННОГО ПОТЕНЦИАЛА (МП/ПП).
ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД).
ПОРОГОВЫЕ И ПОДПОРОГОВЫЕ РАЗДРАЖИТЕЛИ.

Слайд 43Если раздражать нейрон через электрод, находящийся в цитоплазме, кратковременными импульсами деполяризующего электрического
Если раздражать нейрон через электрод, находящийся в цитоплазме, кратковременными импульсами деполяризующего электрического
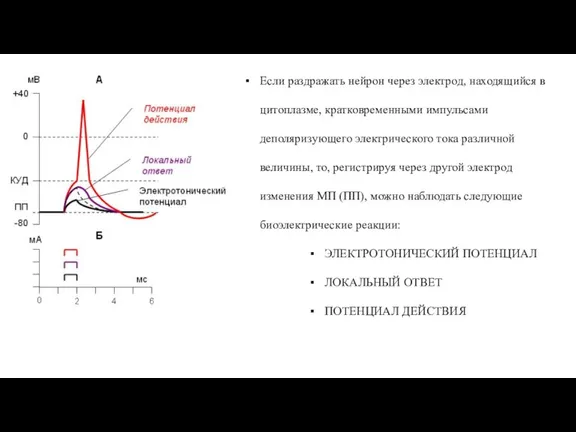
Слайд 44ЭЛЕКТРОТОНИЧЕСКИЙ ПОТЕНЦИАЛ:
Если наносятся раздражения, величина которых не превышает 0.5 величины порогового раздражения,
ЭЛЕКТРОТОНИЧЕСКИЙ ПОТЕНЦИАЛ:
Если наносятся раздражения, величина которых не превышает 0.5 величины порогового раздражения,

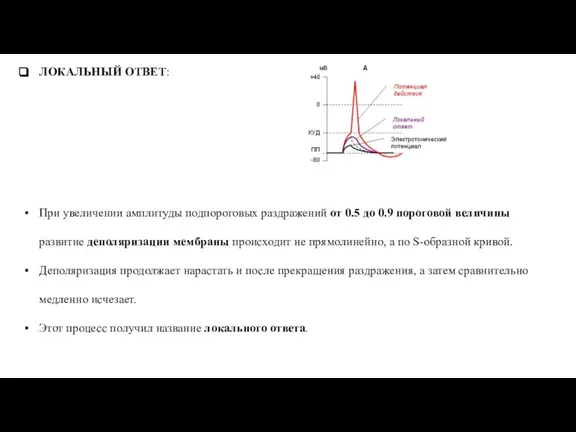
Слайд 45ЛОКАЛЬНЫЙ ОТВЕТ:
При увеличении амплитуды подпороговых раздражений от 0.5 до 0.9 пороговой величины
ЛОКАЛЬНЫЙ ОТВЕТ:
При увеличении амплитуды подпороговых раздражений от 0.5 до 0.9 пороговой величины
Слайд 46СВОЙСТВА ЛОКАЛЬНОГО ОТВЕТА:
Возникает при действии подпороговых раздражителей;
Находится в градуальной зависимости от силы
СВОЙСТВА ЛОКАЛЬНОГО ОТВЕТА:
Возникает при действии подпороговых раздражителей;
Находится в градуальной зависимости от силы

Слайд 47В период развития локального ответа увеличивается поток ионов Na в клетку, что
В период развития локального ответа увеличивается поток ионов Na в клетку, что

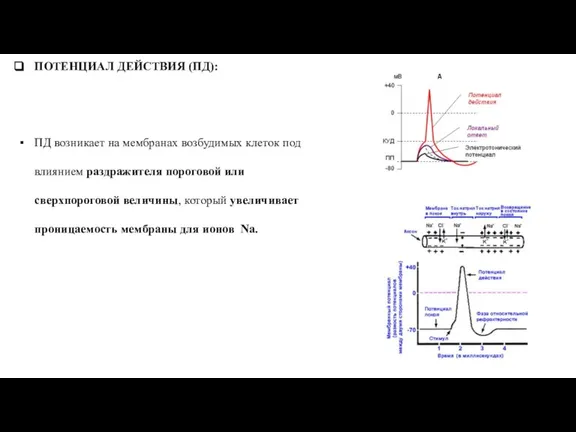
Слайд 48ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД):
ПД возникает на мембранах возбудимых клеток под влиянием раздражителя пороговой
ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД):
ПД возникает на мембранах возбудимых клеток под влиянием раздражителя пороговой
Слайд 49Ионы Na начинают входить внутрь клетки, что приводит к уменьшению величины МП
Ионы Na начинают входить внутрь клетки, что приводит к уменьшению величины МП

Слайд 50В результате проникновения ионов Na в цитоплазму и их взаимодействия с анионами
В результате проникновения ионов Na в цитоплазму и их взаимодействия с анионами

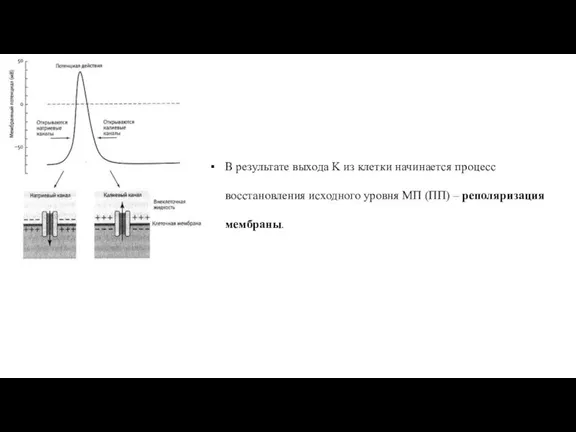
Слайд 51В результате выхода K из клетки начинается процесс восстановления исходного уровня МП
В результате выхода K из клетки начинается процесс восстановления исходного уровня МП
Слайд 52Например:
Если такое повышение проводимости для K предотвратить введением тетраэтиламмония, который избирательно блокирует
Например:
Если такое повышение проводимости для K предотвратить введением тетраэтиламмония, который избирательно блокирует

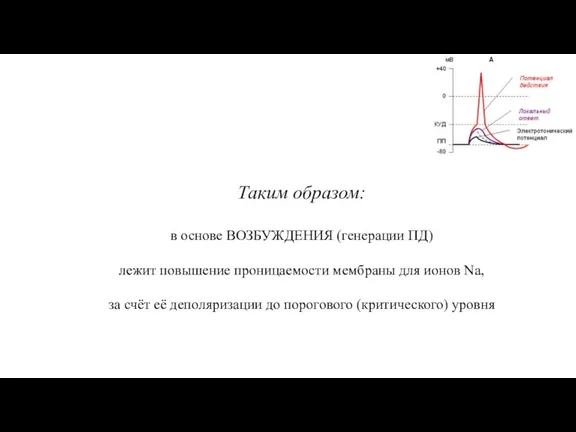
Слайд 53Таким образом:
в основе ВОЗБУЖДЕНИЯ (генерации ПД)
лежит повышение проницаемости мембраны для ионов
Таким образом:
в основе ВОЗБУЖДЕНИЯ (генерации ПД)
лежит повышение проницаемости мембраны для ионов
Слайд 54В потенциале действия (ПД) различают следующие фазы:
Предспайк – процесс медленной деполяризации мембраны
В потенциале действия (ПД) различают следующие фазы:
Предспайк – процесс медленной деполяризации мембраны

Слайд 55ПОТЕНЦИАЛ ДЕЙСТВИЯ В РАЗЛИЧНЫХ ТИПАХ КЛЕТОК ВОЗБУДИМЫХ ТКАНЕЙ
ПОТЕНЦИАЛ ДЕЙСТВИЯ В РАЗЛИЧНЫХ ТИПАХ КЛЕТОК ВОЗБУДИМЫХ ТКАНЕЙ
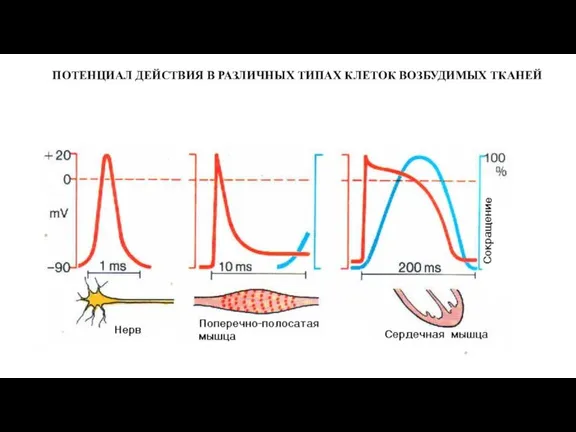
Слайд 564. ИЗМЕНЕНИЕ ВОЗБУДИМОСТИ ПРИ ВОЗБУЖДЕНИИ
4. ИЗМЕНЕНИЕ ВОЗБУДИМОСТИ ПРИ ВОЗБУЖДЕНИИ

Слайд 57ПРИ РАЗВИТИИ ПД ПРОИСХОДЯТ ФАЗНЫЕ ИЗМЕНЕНИЯ
ВОЗБУДИМОСТИ ТКАНЕЙ:
Фаза нормального уровня возбудимости
Фаза повышенной
ПРИ РАЗВИТИИ ПД ПРОИСХОДЯТ ФАЗНЫЕ ИЗМЕНЕНИЯ
ВОЗБУДИМОСТИ ТКАНЕЙ:
Фаза нормального уровня возбудимости
Фаза повышенной

Слайд 581. Фаза. Состоянию исходной поляризации мембраны (МП/ПП) соответствует нормальный уровень возбудимости (НВ).
2.
1. Фаза. Состоянию исходной поляризации мембраны (МП/ПП) соответствует нормальный уровень возбудимости (НВ).
2.

Слайд 593. Фаза. В период развития спайка (пикового потенциала) идёт лавинообразное поступление ионов
3. Фаза. В период развития спайка (пикового потенциала) идёт лавинообразное поступление ионов

Слайд 604. Фаза. После окончания фазы перезарядки мембраны возбудимость её постепенно восстанавливается до
4. Фаза. После окончания фазы перезарядки мембраны возбудимость её постепенно восстанавливается до

Слайд 615. Фаза. Периоду отрицательного следового потенциала соответствует повышенный уровень возбудимости (фаза вторичной
5. Фаза. Периоду отрицательного следового потенциала соответствует повышенный уровень возбудимости (фаза вторичной

Слайд 626. Фаза. В период развития положительного следового потенциала возбудимость ткани понижена –
6. Фаза. В период развития положительного следового потенциала возбудимость ткани понижена –

Слайд 63Таким образом:
Таким образом:
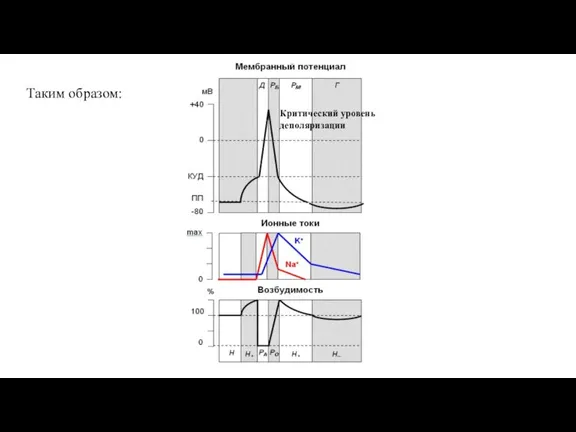
Слайд 64Рефрактерность мембраны является следствием того, что Na-канал состоит из собственного канала (транспортной
Рефрактерность мембраны является следствием того, что Na-канал состоит из собственного канала (транспортной

Слайд 65«Ворота» могут быть полностью открыты или закрыты:
например, в Na-канале в состоянии
«Ворота» могут быть полностью открыты или закрыты:
например, в Na-канале в состоянии

Слайд 66Следовая гиперполяризация мембраны может возникать вследствие трёх причин:
продолжающимся выходом ионов K;
открытием каналов
Следовая гиперполяризация мембраны может возникать вследствие трёх причин:
продолжающимся выходом ионов K;
открытием каналов

Слайд 675. ЗАКОНЫ РАЗДРАЖЕНИЯ ВОЗБУДИМЫХ ТКАНЕЙ
5. ЗАКОНЫ РАЗДРАЖЕНИЯ ВОЗБУДИМЫХ ТКАНЕЙ

Слайд 68 Характер ответных реакций отдельных возбудимых структур и организма в целом зависит
Характер ответных реакций отдельных возбудимых структур и организма в целом зависит

Слайд 69ЗАКОНЫ раздражения возбудимых тканей отражают различные зависимости между действием раздражителя и характером
ЗАКОНЫ раздражения возбудимых тканей отражают различные зависимости между действием раздражителя и характером

Слайд 70Ответная реакция возбудимой ткани зависит от силы раздражения:
При действии слабых (подпороговых) раздражителей
Ответная реакция возбудимой ткани зависит от силы раздражения:
При действии слабых (подпороговых) раздражителей

Слайд 71ЗАКОНЫ раздражения возбудимых тканей:
Закон силы
Закон «всё или ничего»
Закон силы-длительности
Закон раздражения (аккомодации) Дюбуа-Реймона
Закон
ЗАКОНЫ раздражения возбудимых тканей:
Закон силы
Закон «всё или ничего»
Закон силы-длительности
Закон раздражения (аккомодации) Дюбуа-Реймона
Закон

Слайд 721. ЗАКОН СИЛЫ:
Чем больше сила раздражителя, тем сильнее ответная реакция (как целого
1. ЗАКОН СИЛЫ:
Чем больше сила раздражителя, тем сильнее ответная реакция (как целого

Слайд 73Это обусловлено тем, что скелетная мышца состоит из множества мышечных волокон, имеющих
Это обусловлено тем, что скелетная мышца состоит из множества мышечных волокон, имеющих

Слайд 742. ЗАКОН «ВСЁ ИЛИ НИЧЕГО»:
В физиологии существует представление о том, что ряд
2. ЗАКОН «ВСЁ ИЛИ НИЧЕГО»:
В физиологии существует представление о том, что ряд

Слайд 75Закон «всё или ничего» относителен, так как хотя на раздражители подпороговой силы
Закон «всё или ничего» относителен, так как хотя на раздражители подпороговой силы

Слайд 763. ЗАКОН СИЛЫ-ДЛИТЕЛЬНОСТИ:
В начале XX века интенсивно изучался вопрос о соотношении силы
3. ЗАКОН СИЛЫ-ДЛИТЕЛЬНОСТИ:
В начале XX века интенсивно изучался вопрос о соотношении силы

Слайд 77Исследования зависимости силы-длительности показали, что она имеет гиперболический характер.
Ток меньше некоторой минимальной
Исследования зависимости силы-длительности показали, что она имеет гиперболический характер.
Ток меньше некоторой минимальной

Слайд 78Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется
Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется

Слайд 79Хронаксиметрия (от греч. chrons – время, axia – цена, metron – мера)
Хронаксиметрия (от греч. chrons – время, axia – цена, metron – мера)

Слайд 80В клинической практике метод хронаксиметрии можно применять для определения степени снижения возбудимости
В клинической практике метод хронаксиметрии можно применять для определения степени снижения возбудимости

Слайд 814. ЗАКОН РАЗДРАЖЕНИЯ (аккомодации) Дюбуа-Реймона:
Стимулирующее действие постоянного тока зависит не только от
4. ЗАКОН РАЗДРАЖЕНИЯ (аккомодации) Дюбуа-Реймона:
Стимулирующее действие постоянного тока зависит не только от

Слайд 82При снижении скорости нарастания силы раздражителя ПД не возникает, тк деполяризация мембраны
При снижении скорости нарастания силы раздражителя ПД не возникает, тк деполяризация мембраны

Слайд 83При быстром нарастании тока повышение Na-проницаемости успевает достичь значительной величины, прежде чем
При быстром нарастании тока повышение Na-проницаемости успевает достичь значительной величины, прежде чем

Слайд 84Способность к аккомодации различных структур неодинакова:
Наиболее высокая у двигательных нервных волокон;
Значительно меньше
Способность к аккомодации различных структур неодинакова:
Наиболее высокая у двигательных нервных волокон;
Значительно меньше

Слайд 855. ЗАКОН ПОЛЯРНОГО ДЕЙСТВИЯ ПОСТОЯННОГО ТОКА:
При замыкании тока возбуждение возникает под катодом,
5. ЗАКОН ПОЛЯРНОГО ДЕЙСТВИЯ ПОСТОЯННОГО ТОКА:
При замыкании тока возбуждение возникает под катодом,

Слайд 866. ЗАКОН ФИЗИОЛОГИЧЕСКОГО ЭЛЕКТРОТОНА:
Действие постоянного тока на ткань сопровождается изменением её возбудимости.
При
6. ЗАКОН ФИЗИОЛОГИЧЕСКОГО ЭЛЕКТРОТОНА:
Действие постоянного тока на ткань сопровождается изменением её возбудимости.
При

Слайд 87Эти изменения возбудимости под катодом и анодом получили название электротона (электротоническое изменение
Эти изменения возбудимости под катодом и анодом получили название электротона (электротоническое изменение

Слайд 88При дальнейшем действии постоянного тока:
первоначальное повышение возбудимости под катодом сменяется её понижением,
При дальнейшем действии постоянного тока:
первоначальное повышение возбудимости под катодом сменяется её понижением,

Слайд 896. ОСНОВНЫЕ ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА ВОЗБУДИМЫХ ТКАНЕЙ:
Возбудимость
Проводимость
Рефрактерность (невозбудимость)
Лабильность (функциональная подвижность)
Сократимость
6. ОСНОВНЫЕ ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА ВОЗБУДИМЫХ ТКАНЕЙ:
Возбудимость
Проводимость
Рефрактерность (невозбудимость)
Лабильность (функциональная подвижность)
Сократимость

Слайд 90Возбудимость – способность ткани отвечать на раздражение генерацией процесса возбуждения.
Возбудимость зависит
Возбудимость – способность ткани отвечать на раздражение генерацией процесса возбуждения.
Возбудимость зависит

Слайд 912. Проводимость – способность ткани проводить возбуждение по всей своей длине.
Показатель
2. Проводимость – способность ткани проводить возбуждение по всей своей длине.
Показатель

Слайд 923. Рефрактерность (невозбудимость) - способность ткани резко снижать свою возбудимость при возбуждении. В
3. Рефрактерность (невозбудимость) - способность ткани резко снижать свою возбудимость при возбуждении. В

Слайд 934. Лабильность (функциональная подвижность) - способность ткани воспроизводить определенное число волн возбуждения в
4. Лабильность (функциональная подвижность) - способность ткани воспроизводить определенное число волн возбуждения в

Слайд 945. Сократимость – только для мышечной ткани.
5. Сократимость – только для мышечной ткани.

Слайд 95ФИЗИОЛОГИЮ ВОЗБУДИМЫХ ТКАНЕЙ
изучает электрофизиология – раздел физиологии, который исследует
электрические проявления
ФИЗИОЛОГИЮ ВОЗБУДИМЫХ ТКАНЕЙ
изучает электрофизиология – раздел физиологии, который исследует
электрические проявления

 Этапы работы с задачами при обучении одарённных старшеклассников
Этапы работы с задачами при обучении одарённных старшеклассников Принцип обратной связи на примере регулирования уровня углекислого газа
Принцип обратной связи на примере регулирования уровня углекислого газа Ферменты - биологические катализаторы
Ферменты - биологические катализаторы 832648_№3-realizaciya-nasledstvennoi-informacii
832648_№3-realizaciya-nasledstvennoi-informacii Холмогорская порода коров
Холмогорская порода коров Что такое растения
Что такое растения Презентация на тему Дыхание. Органы дыхательной системы. Легкие
Презентация на тему Дыхание. Органы дыхательной системы. Легкие  Презентация на тему Разнообразие пауков
Презентация на тему Разнообразие пауков  Строение, химический состав_
Строение, химический состав_ 20180111_rybalka_na_sogozhe
20180111_rybalka_na_sogozhe Витамины
Витамины Хозяйственно-биологические особенности лошадей
Хозяйственно-биологические особенности лошадей Презентация на тему Сульфатредуцирующие бактерии
Презентация на тему Сульфатредуцирующие бактерии  Презентация на тему Строение и работа мышц
Презентация на тему Строение и работа мышц  Высшие позвоночные – рептилии, птицы, млекопитающие
Высшие позвоночные – рептилии, птицы, млекопитающие Взаимодействие аллельных генов. Кодоминирование
Взаимодействие аллельных генов. Кодоминирование Путешествие вглубь листа
Путешествие вглубь листа Глоссарий
Глоссарий Бобр обыкновенный
Бобр обыкновенный Простейшие. Характерные признаки простейших
Простейшие. Характерные признаки простейших Фотосинтез. Структурные компоненты хлоропласта и их функции
Фотосинтез. Структурные компоненты хлоропласта и их функции Птицы водоёмов и побережья
Птицы водоёмов и побережья Роль диатомей в процессе осадкообразования в морях и континентальных водоемах
Роль диатомей в процессе осадкообразования в морях и континентальных водоемах Принцип сокращение мышц
Принцип сокращение мышц Генотип и фенотип
Генотип и фенотип Анализ нуклеиновых кислот
Анализ нуклеиновых кислот Цитология и генетика
Цитология и генетика Индивидуальное развитие организма
Индивидуальное развитие организма