- Характеристика вирусов. Бактериофаги

Содержание
- 2. Камышева К.С. Основы микробиологии и иммунологии Уч. пособие. «Феникс», 2015 г с.244-257 Подготовка реферативного сообщения по
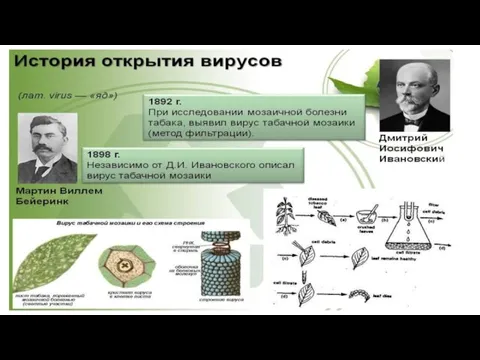
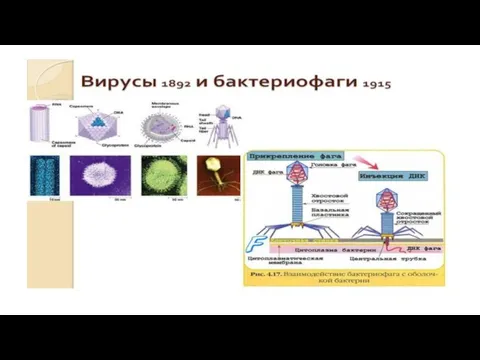
- 3. Открытие вирусов Д.И.Ивановским в 1892г. положило начало развитию науки вирусологии. Более быстрому ее развитию способствовали: изобретение
- 6. Слово “вирус” в переводе с латинского- яд (животного происхождения). Этот термин применяют для обозначения уникальных представителей
- 7. В настоящее время вирусология- бурно развивающаяся наука, что связано с рядом причин: - ведущей ролью вирусов
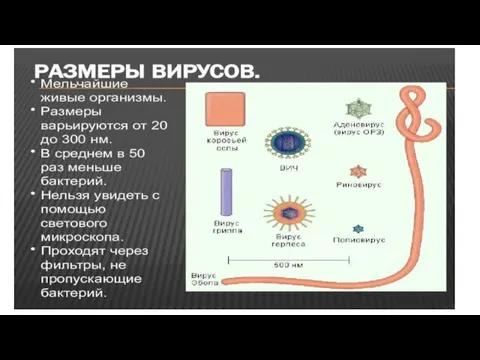
- 8. Основные свойства вирусов (и плазмид), по которым они отличаются от остального живого мира. 1.Ультрамикроскопические размеры (измеряются
- 9. 3.Вирусы не способны к росту и бинарному делению. 4.Вирусы размножаются путем воспроизводства себя в инфицированной клетке
- 10. 6.Средой обитания вирусов являются живые клетки- бактерии (это вирусы бактерий или бактериофаги), клетки растений, животных и
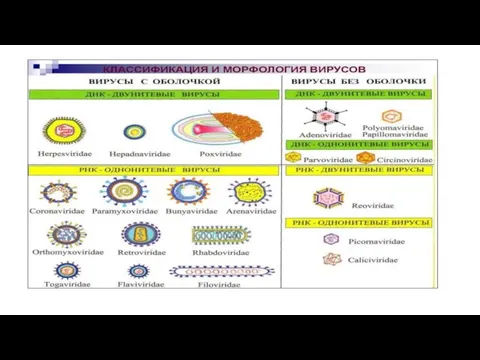
- 12. Классификация и таксономия вирусов. Вирусы составляют царство Vira, которое подразделено по типу нуклеиновой кислоты на два
- 13. В качестве таксономических характеристик первостепенное значение придается типу нуклеиновой кислоты и ее молекулярно-биологическим признакам: двунитевая, однонитевая
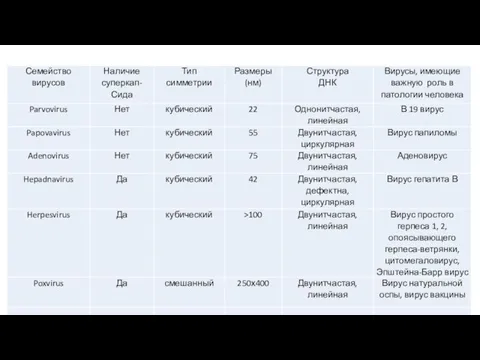
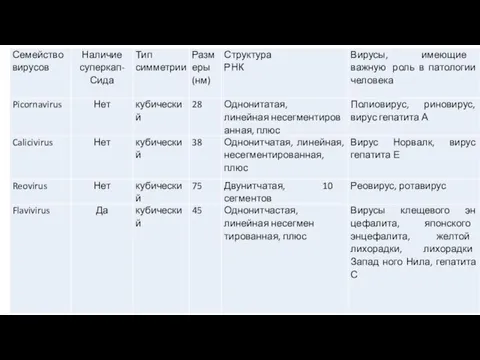
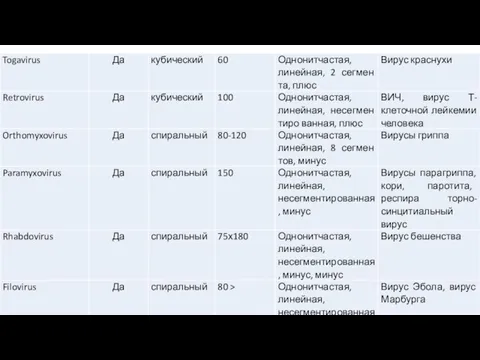
- 14. В настоящее время вирусы человека и животных включены в состав 18 семейств (табл. 1,2). Таблица 1.
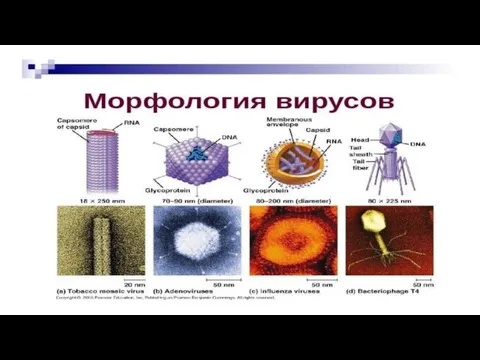
- 20. Строение (морфология) вирусов.
- 22. 1.Геном вирусов образуют нуклеиновые кислоты, представленные одноцепочечными молекулами РНК (у большинства РНК- вирусов) или двухцепочечными молекулами
- 23. При спиральной симметрии белковые субъединицы располагаются по спирали, а между ними, также по спирали, уложена геномная
- 24. 3.Просто устроенные вирусы имеют только нуклеокапсид, т.е. комплекс генома с капсидом и называются “голыми”. 4. У
- 26. ХИМИЧЕСКИЙ СОСТАВ ВИРИОНОВ В состав простых вирионов входит один тип нуклеиновой кислоты — РНК или ДНК
- 28. Основные этапы взаимодействия вируса с клеткой хозяина. 1.Адсорбция - пусковой механизм, связанный со взаимодействием специфических рецепторов
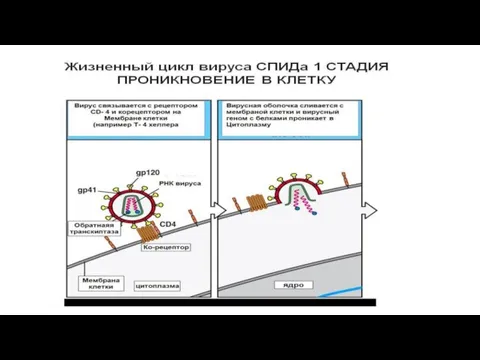
- 29. 2.Проникновение - путем слияния суперкапсида с мембраной клетки или путем эндоцитоза (пиноцитоза). 3.Освобождение нуклеиновых кислот -
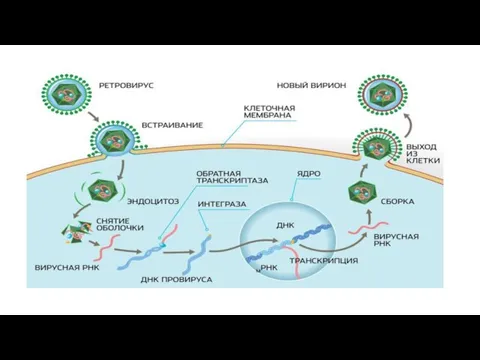
- 31. 4.Синтез нуклеиновых кислот и вирусных белков, т.е. подчинение систем клетки хозяина и их работа на воспроизводство
- 34. Исходы взаимодействия вирусов с клеткой хозяина. 1.Абортивный процесс - когда клетки освобождаются от вируса: - при
- 35. 2.Продуктивный процесс - репликация (продукция) вирусов: - гибель (лизис) клеток (цитопатический эффект)- результат интенсивного размножения и
- 36. 3.Интегративный процесс - интеграция вирусного генома с геномом клетки хозяина. Это особый вариант продуктивного процесса по
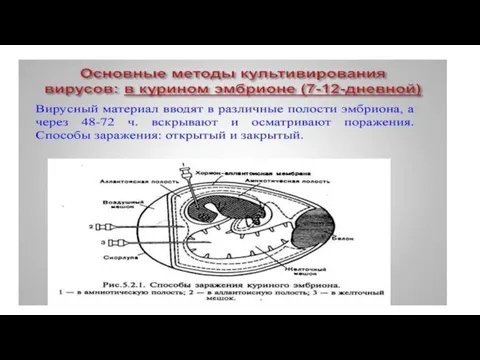
- 37. Методы культивирования вирусов. Вирусы являются облигатными внутриклеточными паразитами, потому они могут репродуцироваться только в живой клетке.
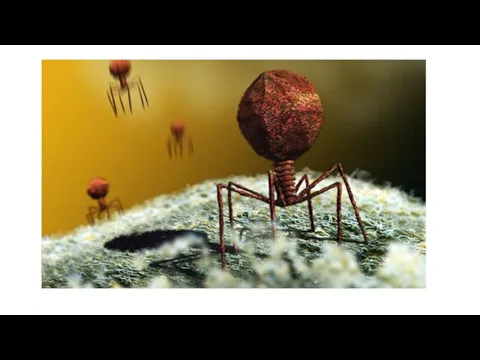
- 42. Вирусы бактерий (бактериофаги). В 1917 г. французский микробиолог Д'Эррель изучал возбудителя дизентерии, наблюдал лизис бактериальной культуры
- 43. Вместе с тем Д'Эррель правильно оценил биологический смысл открытого им феномена. Он высказал предположение, что бактериофаг
- 44. Посев этой же бактериальной культуры на жидкую среду приводит к просветлению среды. Позднее было показано, что
- 45. Номенклатура бактериофагов основана на видовом наименовании хозяина. Например, фаги, лизирующие дизентерийные бактерии, получили название дизентерийных бактериофагов,
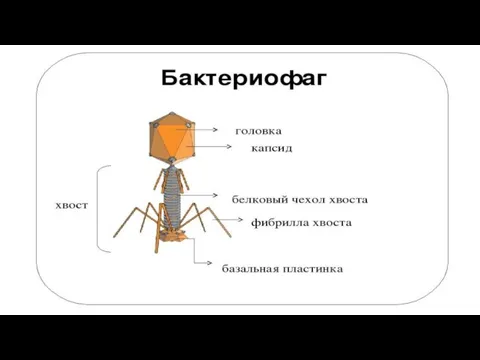
- 46. Структура. Большинство фагов имеют сперматозоидную форму. Они состоят из головки, которая содержит нуклеиновую кислоту, и отростка.
- 49. Химический состав. Фаги, как и другие вирусы, состоят из нуклеиновой кислоты и белка. Большинство их них
- 50. Резистентность к факторам окружающей среды. Фаги более устойчивы к действию физических и химических факторов, чем многие
- 51. Естественной средой обитания фагов является бактериальная клетка, поэтому фаги распространены повсеместно (например, в сточных водах). Фагам
- 53. Основные этапы взаимодействия фагов и бактерий. 1.Адсорбция (взаимодействие специфических рецепторов). 2.Внедрение вирусной ДНК (инъекция фага) осуществляется
- 55. Основные свойства фагов. Антигенные свойства. Бактериофаги содержат группоспецифические и типоспецифические антигены, обладают иммуногенными свойствами, вызывая синтез
- 56. Различают вирулентные фаги, способные вызвать продуктивную форму процесса, и умеренные фаги, вызывающие редуктивную фаговую инфекцию (редукцию
- 57. Бактериальную клетку, несущую в своем геноме профаг, называют лизогенной, поскольку профаг при нарушении синтеза особого белка-
- 58. Умеренные фаги имеют важное значение в обмене генетическим материалом между бактериями- в трансдукции (одна из форм
- 59. По спектру действия на бактерии фаги разделяют на : - поливалентные (лизируют близкородственные бактерии, например сальмонеллы);
- 60. На плотных средах фаги обнаруживают чаще с помощью спот (spot) - теста (образование негативного пятна при
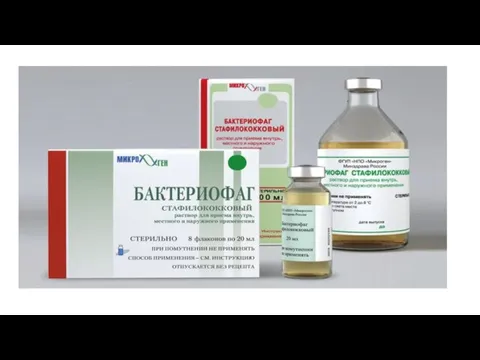
- 61. Практическое использование бактериофагов. 1.Для идентификации (определение фаготипа). 2.Для фагопрофилактики (купирование вспышек). 3.Для фаготерапии (лечение дисбактериозов). 4.Для
- 65. Скачать презентацию

Слайд 3
Открытие вирусов Д.И.Ивановским в 1892г. положило начало развитию науки вирусологии. Более быстрому
Открытие вирусов Д.И.Ивановским в 1892г. положило начало развитию науки вирусологии. Более быстрому


Слайд 6Слово “вирус” в переводе с латинского- яд (животного происхождения). Этот термин применяют
Слово “вирус” в переводе с латинского- яд (животного происхождения). Этот термин применяют

Слайд 7В настоящее время вирусология- бурно развивающаяся наука, что связано с рядом причин:
-
В настоящее время вирусология- бурно развивающаяся наука, что связано с рядом причин:
-

Слайд 8Основные свойства вирусов (и плазмид), по которым они отличаются от остального живого
Основные свойства вирусов (и плазмид), по которым они отличаются от остального живого

Слайд 93.Вирусы не способны к росту и бинарному делению.
4.Вирусы размножаются путем воспроизводства себя
3.Вирусы не способны к росту и бинарному делению.
4.Вирусы размножаются путем воспроизводства себя

Слайд 106.Средой обитания вирусов являются живые клетки- бактерии (это вирусы бактерий или бактериофаги),
6.Средой обитания вирусов являются живые клетки- бактерии (это вирусы бактерий или бактериофаги),


Слайд 12Классификация и таксономия вирусов.
Вирусы составляют царство Vira, которое подразделено по типу нуклеиновой кислоты на
Классификация и таксономия вирусов.
Вирусы составляют царство Vira, которое подразделено по типу нуклеиновой кислоты на

Слайд 13В качестве таксономических характеристик первостепенное значение придается типу нуклеиновой кислоты и ее молекулярно-биологическим
В качестве таксономических характеристик первостепенное значение придается типу нуклеиновой кислоты и ее молекулярно-биологическим

Слайд 14В настоящее время вирусы человека и животных включены в состав 18 семейств (табл.
В настоящее время вирусы человека и животных включены в состав 18 семейств (табл.


Слайд 20Строение (морфология) вирусов.
Строение (морфология) вирусов.
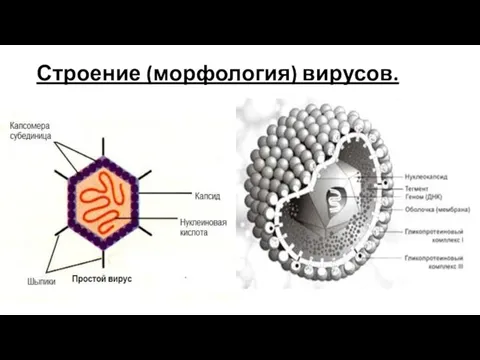
Слайд 221.Геном вирусов образуют нуклеиновые кислоты, представленные одноцепочечными молекулами РНК (у большинства РНК-
1.Геном вирусов образуют нуклеиновые кислоты, представленные одноцепочечными молекулами РНК (у большинства РНК-

Слайд 23При спиральной симметрии белковые субъединицы располагаются по спирали, а между ними, также
При спиральной симметрии белковые субъединицы располагаются по спирали, а между ними, также

Слайд 243.Просто устроенные вирусы имеют только нуклеокапсид, т.е. комплекс генома с капсидом и
3.Просто устроенные вирусы имеют только нуклеокапсид, т.е. комплекс генома с капсидом и

Слайд 26ХИМИЧЕСКИЙ СОСТАВ ВИРИОНОВ
В состав простых вирионов входит один тип нуклеиновой кислоты —
ХИМИЧЕСКИЙ СОСТАВ ВИРИОНОВ
В состав простых вирионов входит один тип нуклеиновой кислоты —


Слайд 28Основные этапы взаимодействия вируса с клеткой хозяина.
1.Адсорбция - пусковой механизм, связанный со
Основные этапы взаимодействия вируса с клеткой хозяина.
1.Адсорбция - пусковой механизм, связанный со

Слайд 292.Проникновение - путем слияния суперкапсида с мембраной клетки или путем эндоцитоза (пиноцитоза).
3.Освобождение
2.Проникновение - путем слияния суперкапсида с мембраной клетки или путем эндоцитоза (пиноцитоза).
3.Освобождение


Слайд 314.Синтез нуклеиновых кислот и вирусных белков, т.е. подчинение систем клетки хозяина и
4.Синтез нуклеиновых кислот и вирусных белков, т.е. подчинение систем клетки хозяина и

Слайд 34Исходы взаимодействия вирусов с клеткой хозяина.
1.Абортивный процесс - когда клетки освобождаются от
Исходы взаимодействия вирусов с клеткой хозяина.
1.Абортивный процесс - когда клетки освобождаются от

Слайд 352.Продуктивный процесс - репликация (продукция) вирусов:
- гибель (лизис) клеток (цитопатический эффект)- результат
2.Продуктивный процесс - репликация (продукция) вирусов:
- гибель (лизис) клеток (цитопатический эффект)- результат

Слайд 363.Интегративный процесс - интеграция вирусного генома с геномом клетки хозяина. Это особый
3.Интегративный процесс - интеграция вирусного генома с геномом клетки хозяина. Это особый

Слайд 37Методы культивирования вирусов.
Вирусы являются облигатными внутриклеточными паразитами, потому они могут репродуцироваться только
Методы культивирования вирусов.
Вирусы являются облигатными внутриклеточными паразитами, потому они могут репродуцироваться только




Слайд 42Вирусы бактерий (бактериофаги).
В 1917 г. французский микробиолог Д'Эррель изучал возбудителя дизентерии, наблюдал
Вирусы бактерий (бактериофаги).
В 1917 г. французский микробиолог Д'Эррель изучал возбудителя дизентерии, наблюдал

Слайд 43Вместе с тем Д'Эррель правильно оценил биологический смысл открытого им феномена.
Он высказал
Вместе с тем Д'Эррель правильно оценил биологический смысл открытого им феномена.
Он высказал

Слайд 44Посев этой же бактериальной культуры на жидкую среду приводит к просветлению среды.
Позднее
Посев этой же бактериальной культуры на жидкую среду приводит к просветлению среды.
Позднее

Слайд 45 Номенклатура бактериофагов основана на видовом наименовании хозяина. Например, фаги, лизирующие дизентерийные
Номенклатура бактериофагов основана на видовом наименовании хозяина. Например, фаги, лизирующие дизентерийные

Слайд 46Структура. Большинство фагов имеют сперматозоидную форму. Они состоят из головки, которая содержит нуклеиновую кислоту, и
Структура. Большинство фагов имеют сперматозоидную форму. Они состоят из головки, которая содержит нуклеиновую кислоту, и


Слайд 49Химический состав. Фаги, как и другие вирусы, состоят из нуклеиновой кислоты и белка.
Большинство
Химический состав. Фаги, как и другие вирусы, состоят из нуклеиновой кислоты и белка.
Большинство

Слайд 50Резистентность к факторам окружающей среды. Фаги более устойчивы к действию физических и химических факторов,
Резистентность к факторам окружающей среды. Фаги более устойчивы к действию физических и химических факторов,

Слайд 51Естественной средой обитания фагов является бактериальная клетка, поэтому фаги распространены повсеместно (например,
Естественной средой обитания фагов является бактериальная клетка, поэтому фаги распространены повсеместно (например,

Слайд 53Основные этапы взаимодействия фагов и бактерий.
1.Адсорбция (взаимодействие специфических рецепторов).
2.Внедрение вирусной ДНК (инъекция
Основные этапы взаимодействия фагов и бактерий.
1.Адсорбция (взаимодействие специфических рецепторов).
2.Внедрение вирусной ДНК (инъекция

Слайд 55Основные свойства фагов.
Антигенные свойства.
Бактериофаги содержат группоспецифические и типоспецифические антигены, обладают иммуногенными
Основные свойства фагов.
Антигенные свойства.
Бактериофаги содержат группоспецифические и типоспецифические антигены, обладают иммуногенными

Слайд 56Различают вирулентные фаги, способные вызвать продуктивную форму процесса, и умеренные фаги, вызывающие
Различают вирулентные фаги, способные вызвать продуктивную форму процесса, и умеренные фаги, вызывающие

Слайд 57 Бактериальную клетку, несущую в своем геноме профаг, называют лизогенной, поскольку профаг
Бактериальную клетку, несущую в своем геноме профаг, называют лизогенной, поскольку профаг

Слайд 58Умеренные фаги имеют важное значение в обмене генетическим материалом между бактериями- в
Умеренные фаги имеют важное значение в обмене генетическим материалом между бактериями- в

Слайд 59По спектру действия на бактерии фаги разделяют на :
- поливалентные (лизируют близкородственные
По спектру действия на бактерии фаги разделяют на :
- поливалентные (лизируют близкородственные

Слайд 60На плотных средах фаги обнаруживают чаще с помощью спот (spot) - теста
На плотных средах фаги обнаруживают чаще с помощью спот (spot) - теста

Слайд 61Практическое использование бактериофагов.
1.Для идентификации (определение фаготипа).
2.Для фагопрофилактики (купирование вспышек).
3.Для фаготерапии (лечение дисбактериозов).
4.Для
Практическое использование бактериофагов.
1.Для идентификации (определение фаготипа).
2.Для фагопрофилактики (купирование вспышек).
3.Для фаготерапии (лечение дисбактериозов).
4.Для

 obmen_veschestv
obmen_veschestv Гликолиз
Гликолиз Половозрастной состав населения России
Половозрастной состав населения России Гаструляция. Типы гаструляции. Характер гаструляции у человека
Гаструляция. Типы гаструляции. Характер гаструляции у человека Мой домашний питомец
Мой домашний питомец Выращивание мелиссы лимонной в деревне Хийденсельга
Выращивание мелиссы лимонной в деревне Хийденсельга Кто и как готовится к зиме
Кто и как готовится к зиме Виды корней и типы корневых систем
Виды корней и типы корневых систем История изучения клетки
История изучения клетки сущность жизни и свойства живого презентация
сущность жизни и свойства живого презентация Цветоводство : виды и значение
Цветоводство : виды и значение Индивидуальное развитие организмов (онтогенез). 6 класс
Индивидуальное развитие организмов (онтогенез). 6 класс Брюхоногие моллюски
Брюхоногие моллюски Мейоз
Мейоз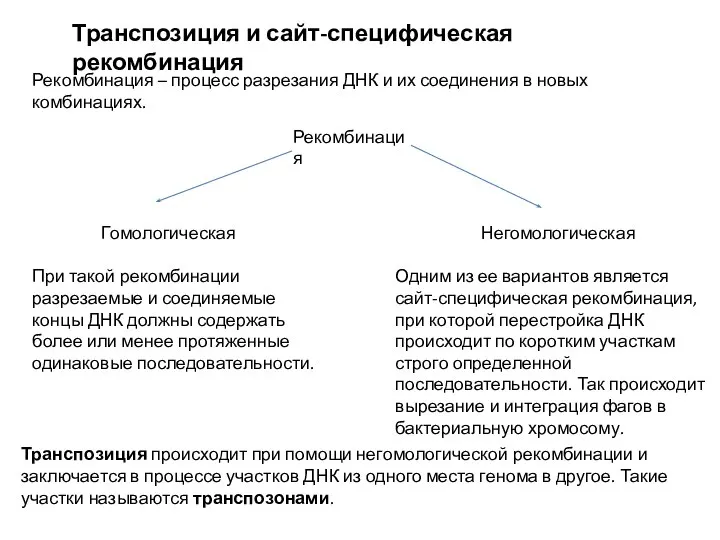 Транспозиция и сайт - специфическая рекомбинация
Транспозиция и сайт - специфическая рекомбинация 6 сыйныф чәчәк һәм җимеш
6 сыйныф чәчәк һәм җимеш 01. Клеточная теория
01. Клеточная теория Системы органов в организме
Системы органов в организме Всемирный день сна
Всемирный день сна Физиология пищеварения в желудке. Лекция № 19
Физиология пищеварения в желудке. Лекция № 19 Физиология спорта Повседневная деятельность человека предусматривает два вида активности: психологическую и физическую. Психол
Физиология спорта Повседневная деятельность человека предусматривает два вида активности: психологическую и физическую. Психол Класс Птицы. Общая характеристика класса
Класс Птицы. Общая характеристика класса Средиземноморская черепаха Никольского
Средиземноморская черепаха Никольского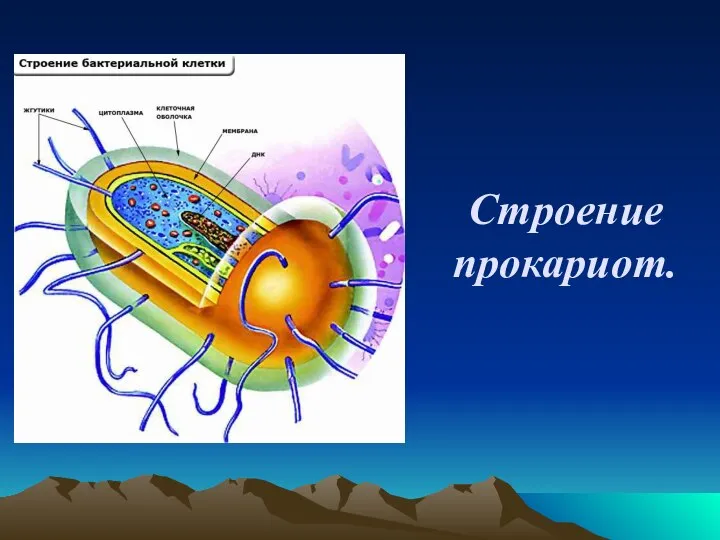 Строение прокариот
Строение прокариот Хемосинтез
Хемосинтез Строение пищеварительной системы
Строение пищеварительной системы Генетика. Пов'язаність генетики з іншими науками та підрозділи генетики. Методи генетики
Генетика. Пов'язаність генетики з іншими науками та підрозділи генетики. Методи генетики Рыбки для маленького аквариума
Рыбки для маленького аквариума