- Химизм фотосинтеза

Содержание
- 2. Фотосинтез включает два главнейших этапа, последовательно связанных между собой. Этап поглощения и преобразования энергии (явление, получившее
- 3. Пигменты растений, участвующие в фотосинтезе, «упакованы» в тилакоиды хлоропластов в виде функциональных фотосинтетических единиц, называемых фотосистемами.
- 4. Все молекулы пигментов в фотосистемах способны улавливать энергию солнечного света. В случае антенных пигментов свет, поглощенный
- 5. Организация фотосинтетического аппарата, «реальная картинка».
- 6. Все пигменты фотосистемы могут поглощать частицы световой энергии, называемые фотонами или квантами света, но только одна
- 7. ФС I В ФС I реакционный центр образован особой молекулой хлорофилла а, обозначаемой как Р700, где
- 8. Фотосистема I в «реальном виде»
- 9. Все молекулы пигментов в фотосистемах способны улавливать энергию солнечного света. В случае антенных пигментов свет, поглощенный
- 10. ФС II В случае ФС II энергия света утилизируется реакционным центром Р680. Возбужденные энергизированные электроны центра
- 11. Фотосистема II: схема реакционного центра (РЦ) 25 белков: D1 + D2 – Гетеродимер, на нем: Р680
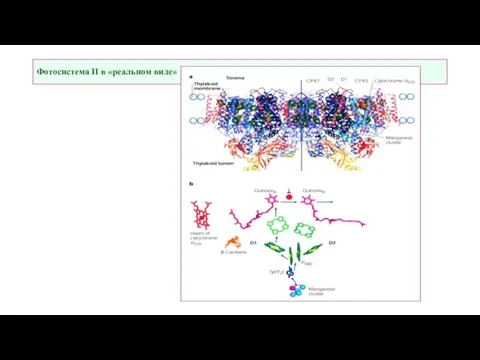
- 12. Фотосистема II в «реальном виде»
- 13. Электронотранспортная цепь между фотосистемами I и II устроена так, что АТФ может образовываться из АДФ и
- 14. Молекула Р680, потерявшая свои электроны (в момент потери происходит флюоресценция), заменяет их электронами донора. В аэробном
- 15. В ФС I энергия света, «уловленная» антенными пигментами фотосистемы, поступает в реакционный центр Р700. От Р700
- 16. ФС I может работать независимо от фотосистемы II. Этот процесс называют циклическим потоком электронов. В ходе
- 17. Темновая фаза фотосинтеза (цикл Кальвина). 6СО2 + 12Н2 + энергия АТФ - С6Н12О6 + 6Н2О (темновой
- 18. Восстановление СО2 до уровня углеводов может происходить в темноте. Этот цикл состоит из трех этапов: 1
- 19. 3 этап - регенерация. Эта фаза происходит с участием трех-, четырех-, пяти-, шести- и семиуглеродных сахаров.
- 20. 6СО2 + 12НАД • Н2 + 18АТФ --» С6Н12О6 + 12НАД + 18АДФ + 18Ф +
- 21. У некоторых растений первый продукт фиксации СО2 — не трехуглеродная молекула 3-глицерофосфата, а четырехуглеродное соединение —
- 22. С4-растения известны среди 19 семейств цветковых (из них 3 — однодольные и 16 — двудольные), и,
- 23. САМ-тип метаболизма. Помимо С3- и С4-путей, известен еще так называемый метаболизм органических кислот по типу толстянковых.
- 24. Завершается фотосинтез накоплением энергетически богатых запасных веществ, составляющих основу для поддержания жизнедеятельности клетки, а в итоге
- 25. Влияние внешних условий на интенсивность фотосинтеза Свет. Источником энергии для фотосинтеза является свет, и поэтому с
- 26. Углекислый газ. Количество СО2 в атмосфере, равное 0,03 %, не является оптимальным для растений. Опыты показали,
- 28. Скачать презентацию
Слайд 2Фотосинтез включает два главнейших этапа, последовательно связанных между собой.
Этап поглощения и
Фотосинтез включает два главнейших этапа, последовательно связанных между собой.
Этап поглощения и

Слайд 3Пигменты растений, участвующие в фотосинтезе, «упакованы» в тилакоиды хлоропластов в виде функциональных
Пигменты растений, участвующие в фотосинтезе, «упакованы» в тилакоиды хлоропластов в виде функциональных

Слайд 4Все молекулы пигментов в фотосистемах способны улавливать энергию солнечного света. В случае
Все молекулы пигментов в фотосистемах способны улавливать энергию солнечного света. В случае
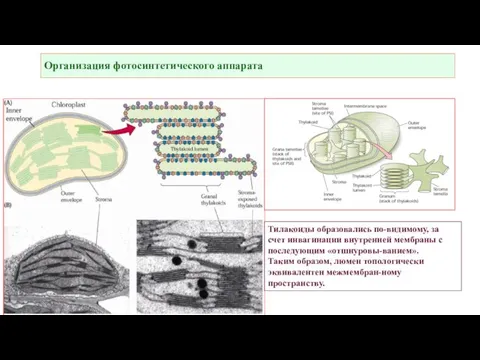
Слайд 5Организация фотосинтетического аппарата, «реальная картинка».
Организация фотосинтетического аппарата, «реальная картинка».
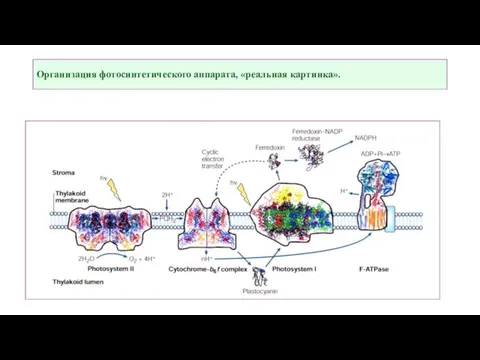
Слайд 6Все пигменты фотосистемы могут поглощать частицы световой энергии, называемые фотонами или квантами
Все пигменты фотосистемы могут поглощать частицы световой энергии, называемые фотонами или квантами

Слайд 7ФС I
В ФС I реакционный центр образован особой молекулой хлорофилла а, обозначаемой
ФС I
В ФС I реакционный центр образован особой молекулой хлорофилла а, обозначаемой

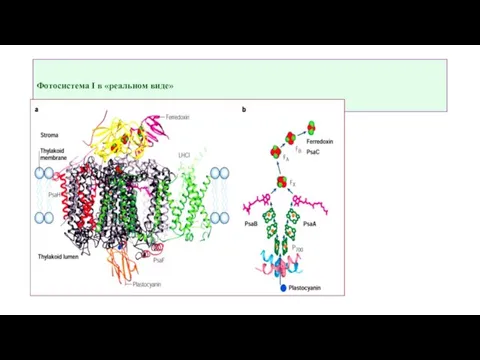
Слайд 8Фотосистема I в «реальном виде»
Фотосистема I в «реальном виде»
Слайд 9 Все молекулы пигментов в фотосистемах способны улавливать энергию солнечного света. В случае
Все молекулы пигментов в фотосистемах способны улавливать энергию солнечного света. В случае

Слайд 10ФС II
В случае ФС II энергия света утилизируется реакционным центром Р680. Возбужденные
ФС II
В случае ФС II энергия света утилизируется реакционным центром Р680. Возбужденные

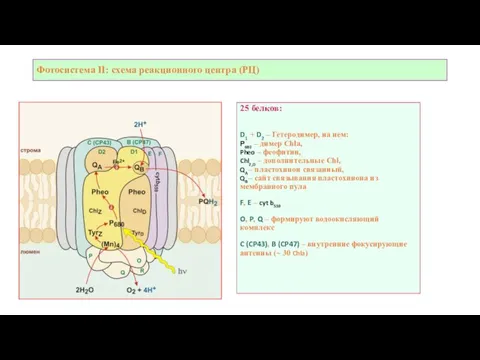
Слайд 11Фотосистема II: схема реакционного центра (РЦ)
25 белков:
D1 + D2 – Гетеродимер, на
Фотосистема II: схема реакционного центра (РЦ)
25 белков:
D1 + D2 – Гетеродимер, на
Слайд 12Фотосистема II в «реальном виде»
Фотосистема II в «реальном виде»
Слайд 13Электронотранспортная цепь между фотосистемами I и II устроена так, что АТФ может
Электронотранспортная цепь между фотосистемами I и II устроена так, что АТФ может

Слайд 14 Молекула Р680, потерявшая свои электроны (в момент потери происходит флюоресценция), заменяет их
Молекула Р680, потерявшая свои электроны (в момент потери происходит флюоресценция), заменяет их

Слайд 15 В ФС I энергия света, «уловленная» антенными пигментами фотосистемы, поступает в реакционный
В ФС I энергия света, «уловленная» антенными пигментами фотосистемы, поступает в реакционный

Слайд 16ФС I может работать независимо от фотосистемы II. Этот процесс называют циклическим
ФС I может работать независимо от фотосистемы II. Этот процесс называют циклическим

Слайд 17Темновая фаза фотосинтеза (цикл Кальвина).
6СО2 + 12Н2 + энергия АТФ -
Темновая фаза фотосинтеза (цикл Кальвина). 6СО2 + 12Н2 + энергия АТФ -

Слайд 18 Восстановление СО2 до уровня углеводов может происходить в темноте. Этот цикл состоит
Восстановление СО2 до уровня углеводов может происходить в темноте. Этот цикл состоит

Слайд 193 этап - регенерация.
Эта фаза происходит с участием трех-, четырех-, пяти-, шести-
3 этап - регенерация.
Эта фаза происходит с участием трех-, четырех-, пяти-, шести-

Слайд 206СО2 + 12НАД • Н2 + 18АТФ --» С6Н12О6 + 12НАД +
6СО2 + 12НАД • Н2 + 18АТФ --» С6Н12О6 + 12НАД +
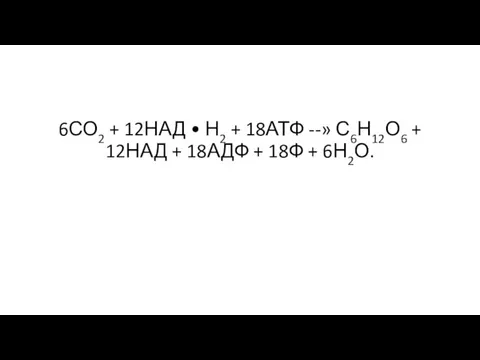
Слайд 21У некоторых растений первый продукт фиксации СО2 — не трехуглеродная молекула 3-глицерофосфата,
У некоторых растений первый продукт фиксации СО2 — не трехуглеродная молекула 3-глицерофосфата,

Слайд 22С4-растения известны среди 19 семейств цветковых (из них 3 — однодольные и
С4-растения известны среди 19 семейств цветковых (из них 3 — однодольные и

Слайд 23САМ-тип метаболизма.
Помимо С3- и С4-путей, известен еще так называемый метаболизм органических
САМ-тип метаболизма.
Помимо С3- и С4-путей, известен еще так называемый метаболизм органических

Слайд 24Завершается фотосинтез накоплением энергетически богатых запасных веществ, составляющих основу для поддержания жизнедеятельности
Завершается фотосинтез накоплением энергетически богатых запасных веществ, составляющих основу для поддержания жизнедеятельности

Слайд 25Влияние внешних условий на интенсивность фотосинтеза
Свет. Источником энергии для фотосинтеза является свет,
Влияние внешних условий на интенсивность фотосинтеза
Свет. Источником энергии для фотосинтеза является свет,

Слайд 26Углекислый газ.
Количество СО2 в атмосфере, равное 0,03 %, не является оптимальным
Углекислый газ.
Количество СО2 в атмосфере, равное 0,03 %, не является оптимальным

 Скат Манта
Скат Манта Wireless Recording in the Peripheral Nervous System with Ultrasonic Neural Dust
Wireless Recording in the Peripheral Nervous System with Ultrasonic Neural Dust Кровоснабжение органов брюшной полости
Кровоснабжение органов брюшной полости Презентация на тему Особенности организации моллюсков
Презентация на тему Особенности организации моллюсков  Зоопсихолог. Зоопсихология
Зоопсихолог. Зоопсихология Физиология желез внутренней секреции (частная)
Физиология желез внутренней секреции (частная) Выведение породы животных
Выведение породы животных Общая классификация отдела Плауны
Общая классификация отдела Плауны Презентация на тему Прибрежно-водные растения
Презентация на тему Прибрежно-водные растения  Как животные защищаются от врагов? Выполнила учитель лицея№180 г.Нижнего Новгорода Бахметова Марина Валерьевна
Как животные защищаются от врагов? Выполнила учитель лицея№180 г.Нижнего Новгорода Бахметова Марина Валерьевна Транспортные системы организма
Транспортные системы организма Подготовительный этап развития биологии. Древнеримский период
Подготовительный этап развития биологии. Древнеримский период Мифические лекарственные растения. Семейство Сложноцветные и Розоцветные
Мифические лекарственные растения. Семейство Сложноцветные и Розоцветные Спинной мозг Форма, топография, основные отделы спинного мозга
Спинной мозг Форма, топография, основные отделы спинного мозга Тема урока: Органические молекулы. Белки, структура и свойства, функции.
Тема урока: Органические молекулы. Белки, структура и свойства, функции. Видоизменения корня
Видоизменения корня Пировиноградная кислота
Пировиноградная кислота Класс Птицы
Класс Птицы Презентация на тему Первые вестники весны - Первоцветы
Презентация на тему Первые вестники весны - Первоцветы  Рептилії. Життєві форми. Будова тіла
Рептилії. Життєві форми. Будова тіла Обрезка плодовых деревьев и кустарников
Обрезка плодовых деревьев и кустарников Ферментативная функция
Ферментативная функция Биогеноценоз и экосистемы - сходства и различия
Биогеноценоз и экосистемы - сходства и различия Корень. Корневые системы
Корень. Корневые системы Орлык төзелеше. 6 класс
Орлык төзелеше. 6 класс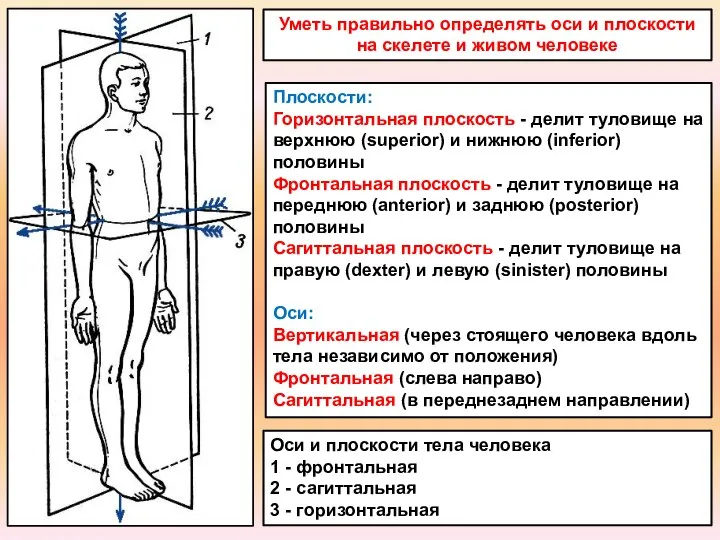 Оси и плоскости.Строение трубчатой кости
Оси и плоскости.Строение трубчатой кости Фенотипическая изменчивость
Фенотипическая изменчивость Надкласс Рыбы. Класс Костные рыбы
Надкласс Рыбы. Класс Костные рыбы