- Клетки места (продолжение). Клетки направления головы (head direction system)

Содержание
- 2. Обучение в условиях конфликта зрительной и обонятельной информации Muzzio et al. (2009). PLoS Biol 7(6): e1000140.
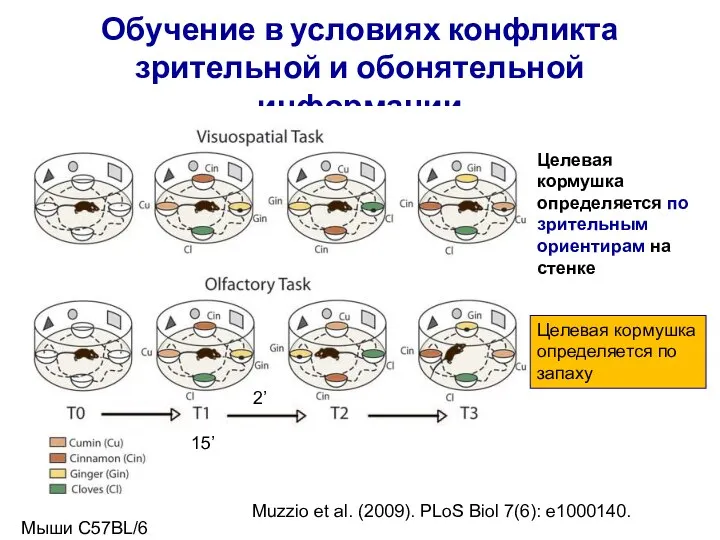
- 3. Обучение по стабильным зрительным ориентирам, стабильные поля разряда PC, несмотря на изменение положения запахов Нейрон А
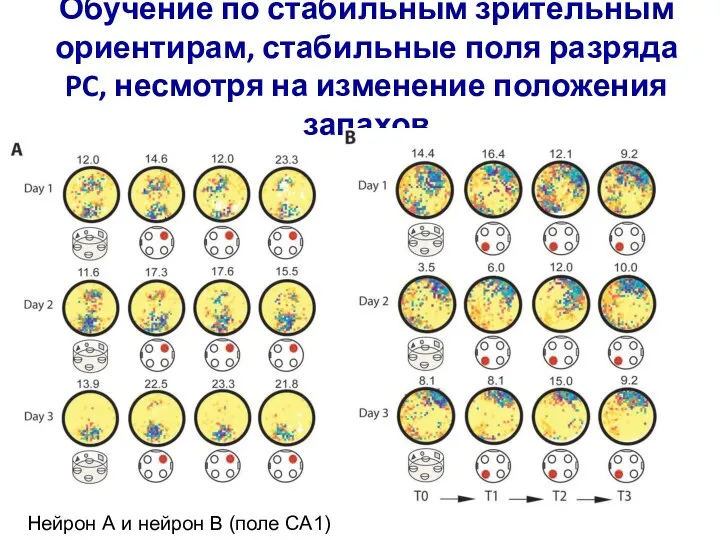
- 4. Дезорганизация полей разряда PC, появление дополнительных полей в области кормушки с целевым запахом
- 5. Феномен “replay” и память Что показывает одновременная регистрация активности многих клеток места
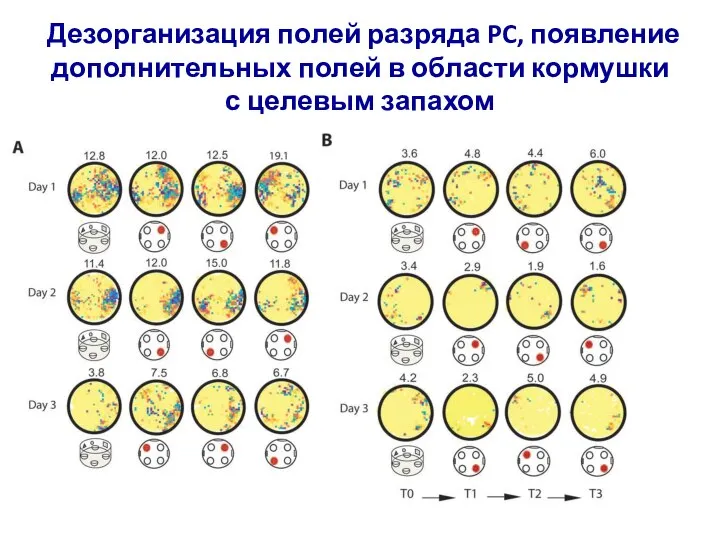
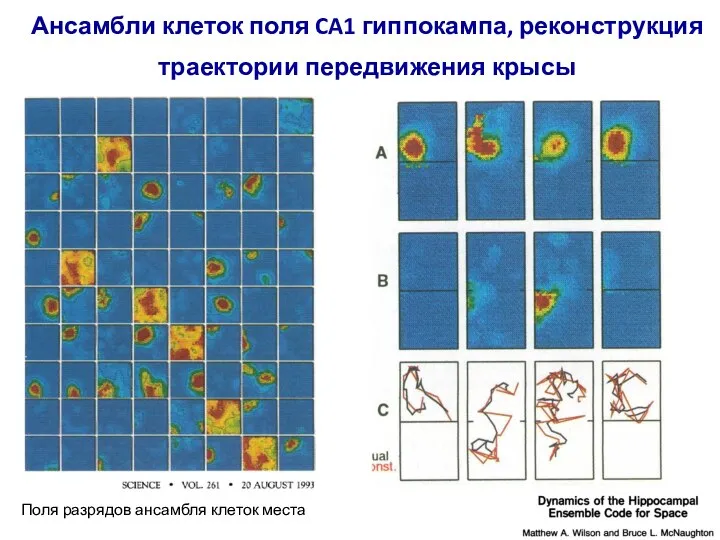
- 6. Ансамбли клеток поля CA1 гиппокампа, реконструкция траектории передвижения крысы Поля разрядов ансамбля клеток места
- 7. Так как клетки места гиппокампа активируются в определенных местах и в определенном порядке, то можно реконструировать
- 9. “Replay” – последовательное, организованное по времени повторное «проигрывание» активности группы нейронов гиппокампа в период низкой активности
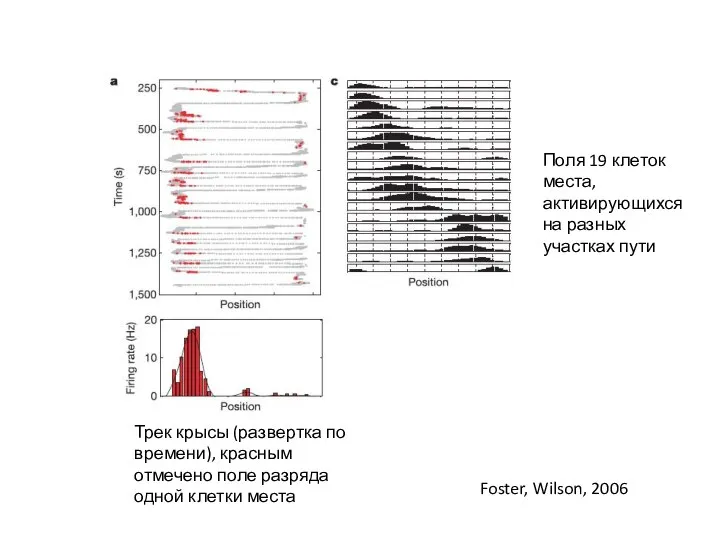
- 10. Трек крысы (развертка по времени), красным отмечено поле разряда одной клетки места Поля 19 клеток места,
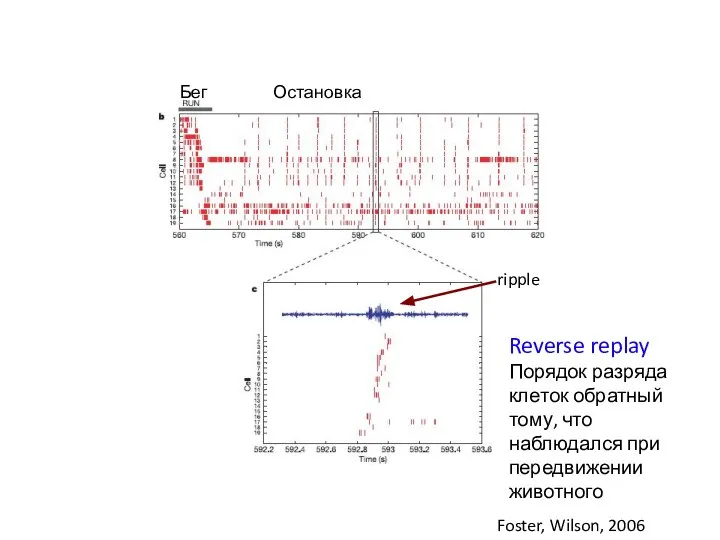
- 11. Reverse replay Порядок разряда клеток обратный тому, что наблюдался при передвижении животного Бег Остановка Foster, Wilson,
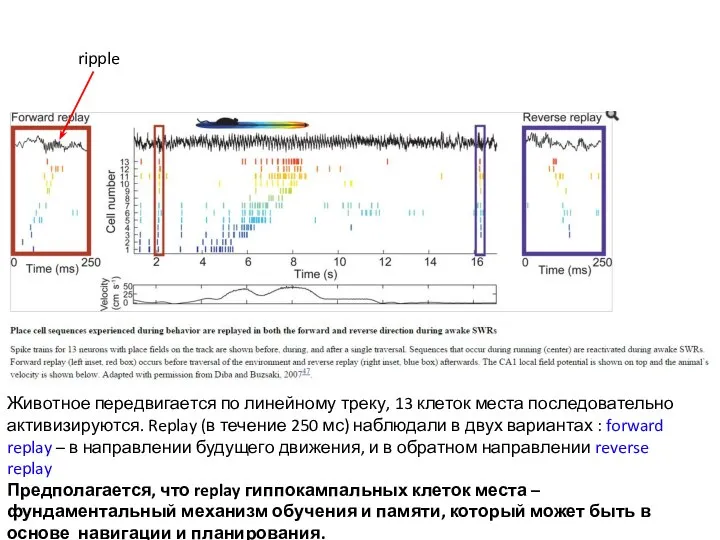
- 12. ripple Животное передвигается по линейному треку, 13 клеток места последовательно активизируются. Replay (в течение 250 мс)
- 13. Функциональная неоднородность гиппокампа вдоль септотемпоральной (рострокаудальной для грызунов) оси
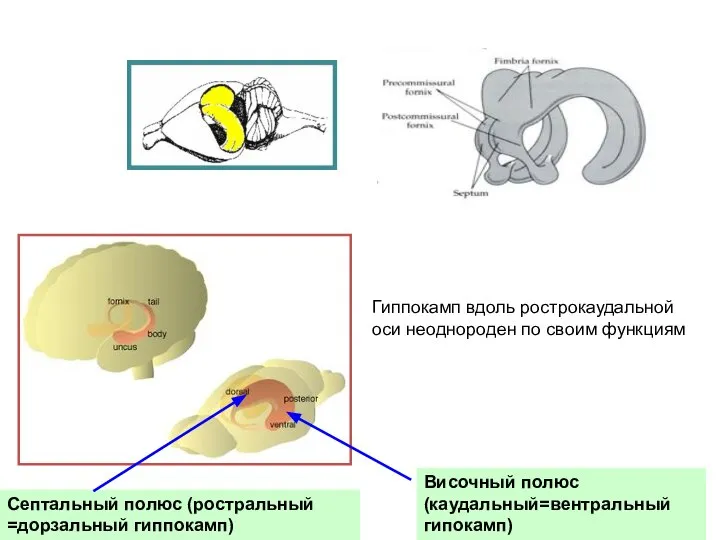
- 14. Септальный полюс (ростральный =дорзальный гиппокамп) Височный полюс (каудальный=вентральный гипокамп) Гиппокамп вдоль рострокаудальной оси неоднороден по своим
- 15. Поля гиппокампа (CA1, CA3 и зубчатая фасция), расположение на разных уровнях рострокаудальной оси
- 16. Молекулярные домены гиппокампа Вид с латеральной стороны Вид с медиальной стороны Fanselow, Dong, 2010 C помощью
- 17. Молекулярные домены гиппокампа Fanselow, Dong, 2010 C помощью набора генетических маркеров выявлены 3 функциональных домена: Дорзальный
- 18. Основные связи в гиппокампальной формации Зубчатая фасция Медиальн. Латеральн. септум Энторинальная кора
- 19. Парагиппокампальная область PER – периринальная кора, POR – постринальная кора, EC – энторинальная кора rs –
- 20. LEA – латеральная энторинальная кора, MEA – медиальная энторинальная кора, PR=PER – периринальная кора, POR –
- 21. Hartman, Lever, Burgess, O’Keefe, 2014
- 22. Топология проекций энторинальной коры (ЭК) в зубчатую извилину A – полушария (мозжечок снят), MEA – медиальная
- 23. Функциональная неоднородность гиппокампа Bast, 2007
- 24. (Bast et al., 2009) Области септальной и промежуточной части гиппокампа больше связаны с обработкой зрительной и
- 25. Свойства клеток места в разных субрегионах гиппокампа вдоль септотемпоральной оси Пространственно-специфичные нейроны обнаружены в промежуточной и
- 26. Клетки места при передвижении по дорожке на большие расстояния 18 м Kjelstrup et al., 2008
- 27. Характеристики клеток места вдоль септо-темпоральной оси гиппокампа 8 клеток места поля CA3 крысы, бежавшей по дорожке
- 28. Поля дорзального и вентрального гиппокампа различаются по пространственной специфичности
- 29. Особенности клеток места каудального (вентрального) гиппокампа Клетки места каудального (вентрального) гиппокампа имеют более широкие поля разрядов
- 30. Особенности клеток места каудального гиппокампа Пирамидные клетки и, в меньшей степени, интернейроны специфически разряжались в рукавах
- 31. Более каудально расположенные CA3 нейроны гиппокампа имеют большие поля, чем ростральные. Обнаружено участие нейронов средней и
- 32. Клетки дорзального гиппокампа сразу начинают избирательно активироваться в одном из контекстов Клетки промежуточной и вентральной области
- 33. Исследование функциональной гетерогенности гиппокампа K.V. Anokhin R. Deacon (Oxford, UK) V.M. Malygin P. A. Kuptsov, лаборатория
- 34. Функциональные особенности каудальных областей гиппокампа грызунов (наши данные) Картирование иммуногистохимическим методом (по уровню экспрессии с-Fos) активации
- 35. Области каудального гиппокампа значительно активировались у мышей и полевок, исследовавших арену или лабиринт (Купцов и др.
- 36. C57BL/6 Рыжая полевка Активное передвижение животных по арене «открытого поля» увеличивает активацию средних и задних отделов
- 37. Купцов, 2006 Введение новых зрительных ориентиров (синяя, красная и зеленая линия) усиливает активацию только в ростральных
- 38. Клетки места у представителей других видов
- 40. Клетки места дорзального гиппокампа шиншиллы Диам. 76 см Muir et al., 2009 Отр. Грызуны, сем. Шиншилловые
- 41. Клетки места у летучей мыши Ulanovsky, Moss, 2007, 2011 Клетки регистрировали в септальной части поля CA1
- 42. Форма поля разряда клетки места и биология вида Ulanovsky 2011 Hayman et al., 2011 ??? Будет
- 43. Объемные поля клеток места в гиппокампе крыланов в полете Yartsev, Ulanovsky, 2013 Клетки места (CA1) были
- 44. Клетки места, зависимость от направления головы (направления движения) Rubin, Yartsev, Ulanovsky, 2014 Бурый кожан, Eptesicus fuscus
- 45. Клетки места, зависимость от направления головы (направления движения) Rubin, Yartsev, Ulanovsky, 2014 Клетка поля CA1 летучей
- 46. Кодирование пространства у обезьян Саймири свободно передвигались по камере (1.5 х 1.5 х 1.2), собирали фрукты,
- 47. Гиппокамп у мыши и голубя Butler et al., 2006
- 48. Пространственно-специфичные клетки гиппокампа почтового голубя Siegel, J. J., Nitz, D., & Bingman, V. P. (2005). Spatial
- 49. Человек (пациенты с вживленными по медицинским показаниям электродами в область эпилептического очага) Путешествие по виртуальному городу
- 50. Области мозга, где обнаружены пространственно специфичные клетки Гиппокамп (Аммонов рог, зубчатая фасция) Субикулюм Энторинальная кора
- 51. Для успешной навигации в пространстве необходимо не только определение своего местоположения, но и контроль направления передвижения
- 52. Клетки направления головы (head direction cells, HD cells)
- 53. Парагиппокампальная область PER – периринальная кора, POR – постринальная кора, EC – энторинальная кора rs –
- 54. Гиппокамп и соседние области Witter, Amaral, 2004
- 55. Subiculum, pre- и parasubiculum Крыса, горизонтальный срез переднего мозга, правое полушарие Дорзальная часть пресубикулюма часто выделяют
- 56. Энторинальная кора (медиальная, MEA и латеральная, LEA) Периринальная кора (A35 и A36)
- 57. Клетки направления головы (HD, head direction cells) Sharp, 2010 Частота генерации спайков зависит от направления головы
- 58. HD клетки, активность. Видео Taube, 2007
- 59. Jeffrey Taube Department of Psychological and Brain Sciences at Dartmouth 2005
- 60. Характеристики HD сигнала Предпочитаемое направление Направление головы, град Частота разряда HD клетка реагирует только на направление
- 61. HD клетки сначала были обнаружены в постсубикулуме, позже найдены в переднем дорзальном ядре таламуса (AND), каудальном
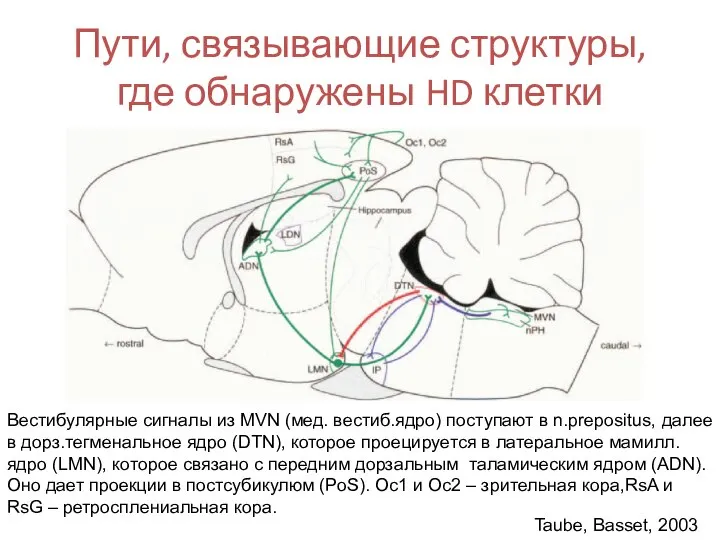
- 62. Пути, связывающие структуры, где обнаружены HD клетки Taube, Basset, 2003 Вестибулярные сигналы из MVN (мед. вестиб.ядро)
- 63. Доля клеток, демонстрирующих HD свойста, в разных структурах мозга Наиболее HD клетки представлены в переднем дорзальном
- 64. Характеристики клеток направления головы в разных структурах мозга Taube, Basset, 2003 Характеристики HD клеток могут различаться
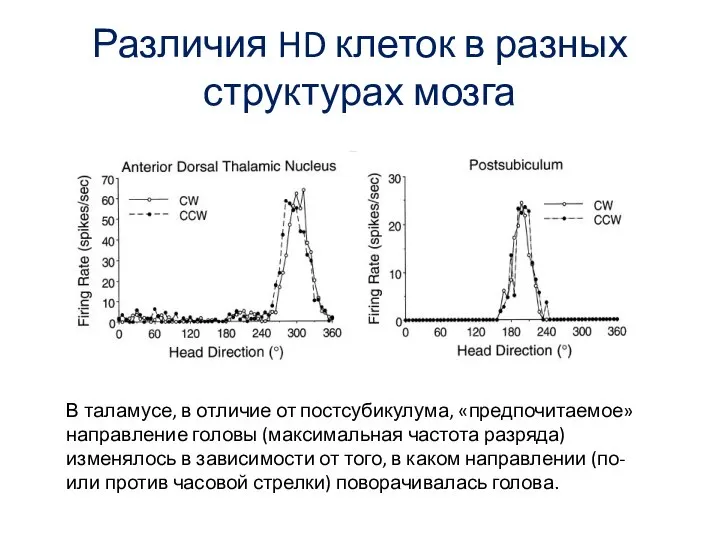
- 65. Различия HD клеток в разных структурах мозга В таламусе, в отличие от постсубикулума, «предпочитаемое» направление головы
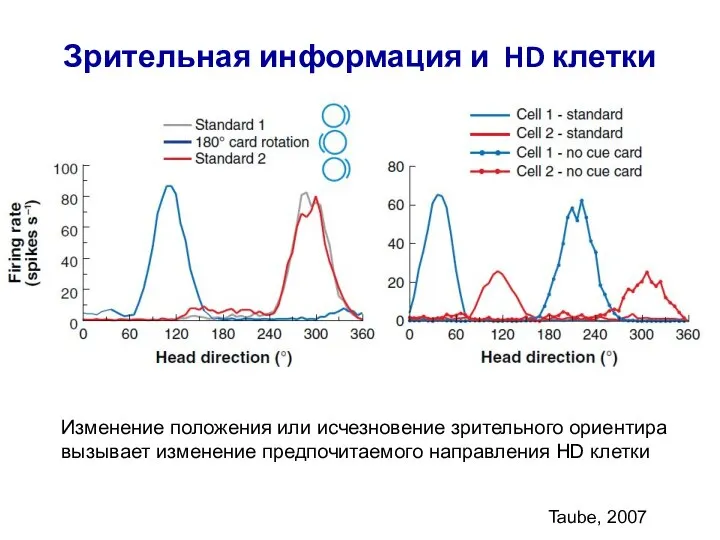
- 66. Зрительная информация и HD клетки Taube, 2007 Изменение положения или исчезновение зрительного ориентира вызывает изменение предпочитаемого
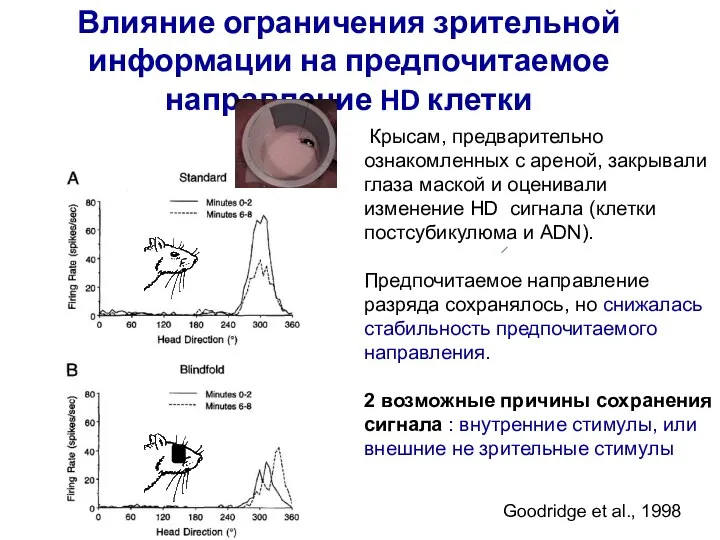
- 67. Влияние ограничения зрительной информации на предпочитаемое направление HD клетки Goodridge et al., 1998 Крысам, предварительно ознакомленных
- 68. Влияние длительности экспозиции нового зрительного ориентира Goodridge et al., 1998 Корректное переключение предпочитаемого направления HD клетки
- 69. Влияние «незрительной» информации Вращение цилиндра(стенок) и пола арены на 90° вызывало соответствующее смещение предпочитаемого направления. Возможное
- 70. Влияние звука Goodridge et al., 1998 Эксперименты проводили в темноте, через колонку подавали звуковой сигнал (щелчок
- 71. Предпочитаемое направление HD контролируется не только внешними, но и внутренними стимулами (от самого животного) При переходе
- 72. Предпочитаемое направление HD контролируется не только внешними, но и внутренними стимулами (от самого животного) Крыс высаживали
- 73. Характеристики HD клеток Разнообразные сенсорные сигналы могут определять предпочитаемое направление и изменять характеристики HD ответа Также
- 74. Эффекты повреждений вестибулярной системы на HD клетки Sodium arsanilate Eliminated HD signal (Stackman and Taube 1997)
- 75. Эффекты временной вестибулярной инактивации (тетродотоксином) на клетки места dCA1 и HD клетки (постсубикулум) Stackman, Clark, Taube,
- 76. HD клетки и передвижение вверх ногами Calton and Taube 2005 Чтобы получить корм в отсеке на
- 77. При передвижении крысы по потолку HD сигнал (anterodorsal thalamic nucleus) пропадал у половины зарегистрированных клеток, у
- 78. Клетки направления головы и вестибулярная система Yoder, Taube, 2014 Клетки направления головы обнаружены в разных структурах
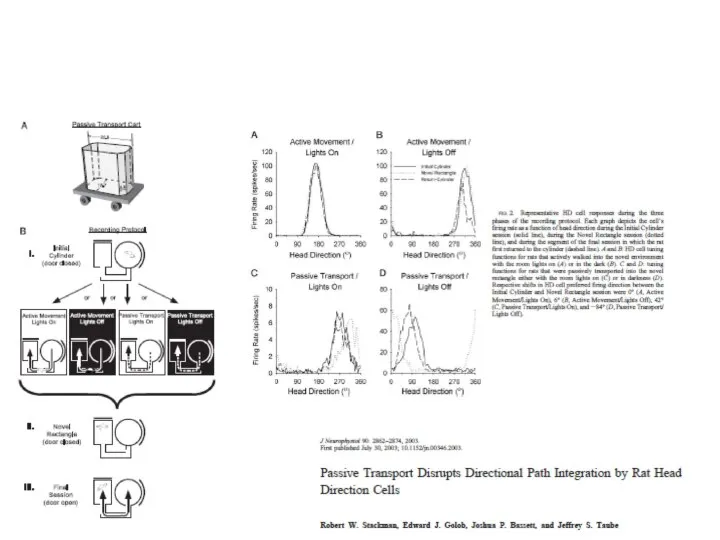
- 79. Интеграция пути (path integration) и HD клетки Stackman,…..Taube, 2003 Зрит. ориентиры Крыс или выпускали из знакомого
- 80. Предпочитаемое направление HD клеток у крыс, свободно передвигающихся или перевозимых на тележке в новый отсек Stackman,…..Taube,
- 81. Клетки, сочетающие признаки клеток места и HD (Theta-Modulated Place-by-Direction Cells) F. Cacucci, C. Lever, T. J.
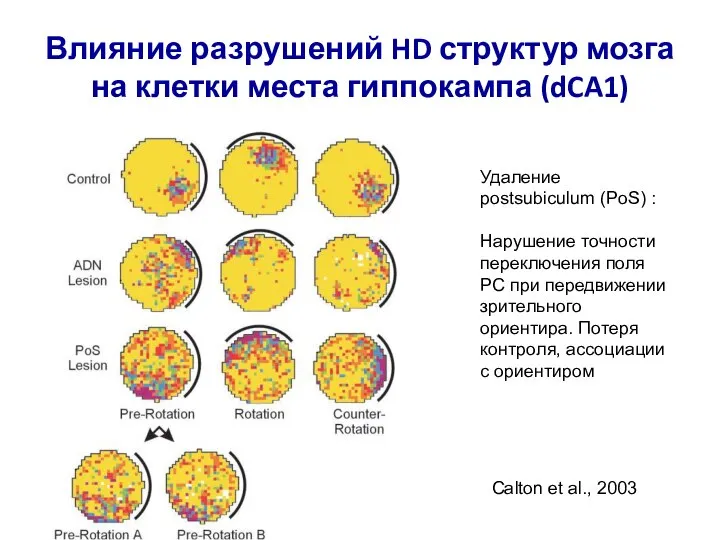
- 82. Влияние разрушений HD структур мозга на клетки места гиппокампа (dCA1) Уменьшение пространственной специфичности и стабильности разряда
- 83. Влияние разрушений HD структур мозга на клетки места гиппокампа (dCA1) Удаление postsubiculum (PoS) : Нарушение точности
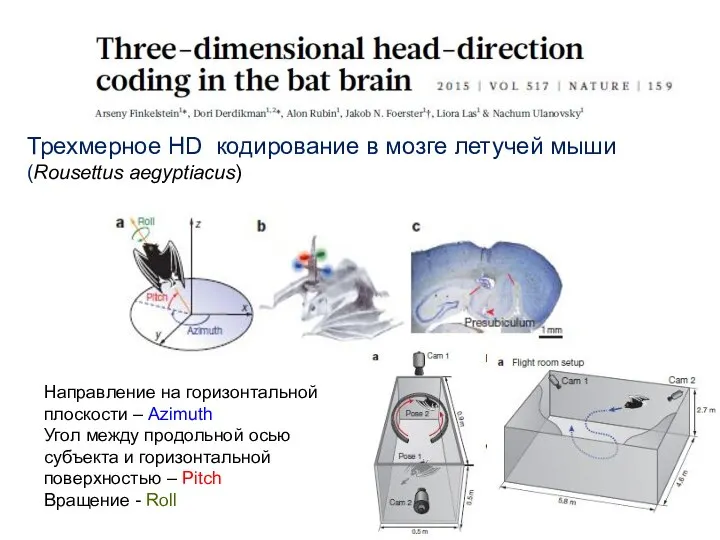
- 84. Трехмерное HD кодирование в мозге летучей мыши (Rousettus aegyptiacus) Направление на горизонтальной плоскости – Azimuth Угол
- 85. Типы HD клеток у летучих мышей Активность зависит только от наклона над горизонтальной плоскостью Взаимодействие факторов
- 86. «HD cells are similar to a compass in that their discharge is always tuned to a
- 87. Спасибо за внимание
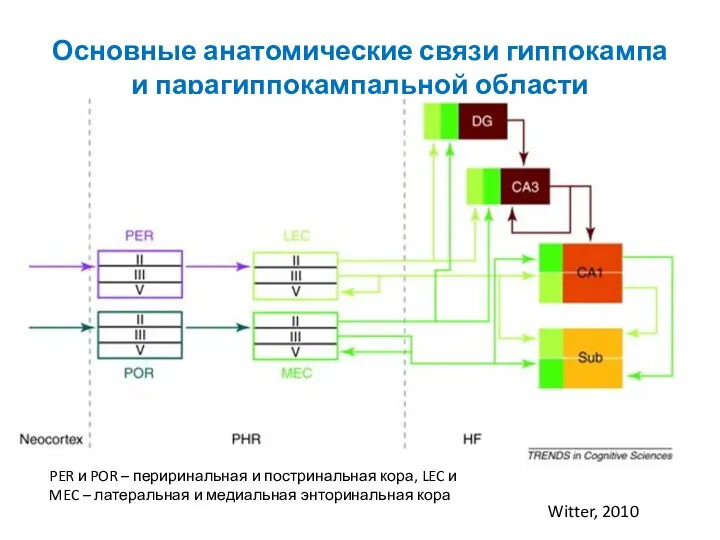
- 88. Основные анатомические связи гиппокампа и парагиппокампальной области Witter, 2010 PER и POR – периринальная и постринальная
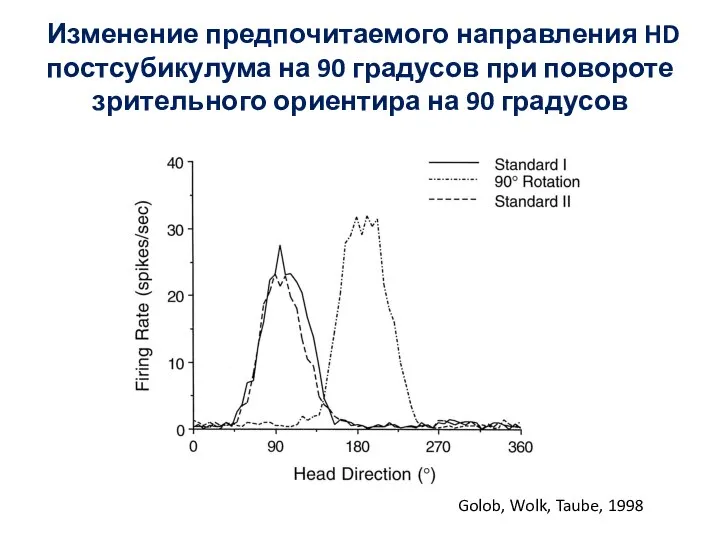
- 89. Изменение предпочитаемого направления HD постсубикулума на 90 градусов при повороте зрительного ориентира на 90 градусов Golob,
- 90. Три проекции поля CA1 Дорзальная часть CA1 Промежуточная и вентральная часть CA1 CA1 Sub EC Ретросплениальная
- 91. Ансамбли клеток поля CA1 гиппокампа, реконструкция траектории передвижения крысы Поля разрядов ансамбля клеток места
- 92. Краткое содержание предыдущей серии…. Ч.1 1. Эдвард Толмен: идея когнитивной карты 1.1 Ментальное представление среды 1.2
- 93. Когнитивные карты. Представления Толмена В процессе обучения в мозге крысы образуется нечто подобное карте окружающей среды.
- 94. Краткое содержание предыдущей серии…. Ч.2 2.3 Исследовательская активность – необходимый компонент построения и контроля карты 2.4
- 95. Place representation как часть когнитивной карты Строятся с использованием двух типов стимулов : На основе набора
- 97. Определение понятия «карты» по O’Keefe, Nadel,1978 The simplest definition of a map is that it is
- 98. Когнитивные карты (O’Keefe, Nadel, 1978) «Когда животное двигается, «внутренняя навигационная» система будет переключать фокус возбуждения внутри
- 99. История экспериментального изучения функций мозга Jean Pierre Flourens (1794-1867) Франция Обнаружил анестезирующий эффект хлороформа В 1825
- 100. Схема областей мозга, содержащих клетки, связанные с контролем навигации животного ADN- передне-дорзальное ядро переднего таламуса, DTN
- 101. Пути, связывающие структуры, где обнаружены HD клетки Taube, Basset, 2003 Вестибулярные сигналы из MVN (мед. вестиб.ядро)
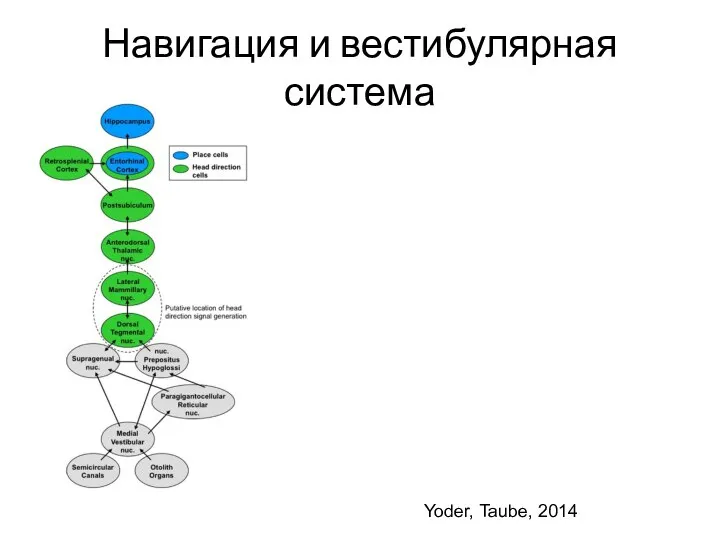
- 102. Навигация и вестибулярная система Yoder, Taube, 2014
- 105. Отличия hd в разных структурах Постсубикулуюм – есть реакция на место
- 106. When the shape of the animal's environment is changed, for example, from a cylinder to a
- 107. the hippocampus acts as a cognitive mapping system, which we shall call the locale system and
- 108. 75 cm 220 cm Лебедев, Плескачева, Анохин, ЖВНД, 2012 Ø 35см Ø 75см Ø 150см Ø
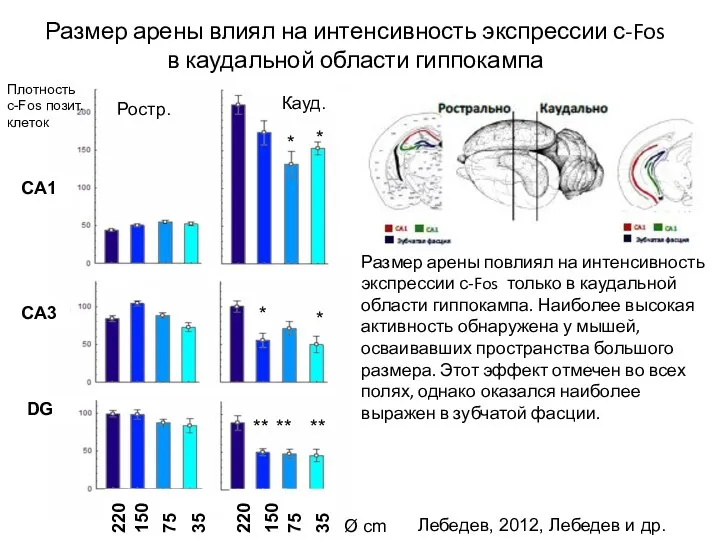
- 109. Размер арены влиял на интенсивность экспрессии с-Fos в каудальной области гиппокампа Лебедев, 2012, Лебедев и др.
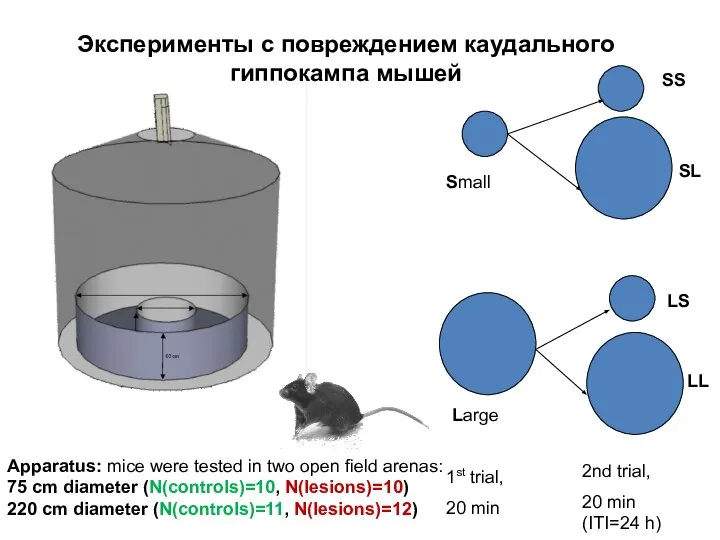
- 110. 1st trial, 20 min 2nd trial, 20 min (ITI=24 h) Small Large SS SL LL LS
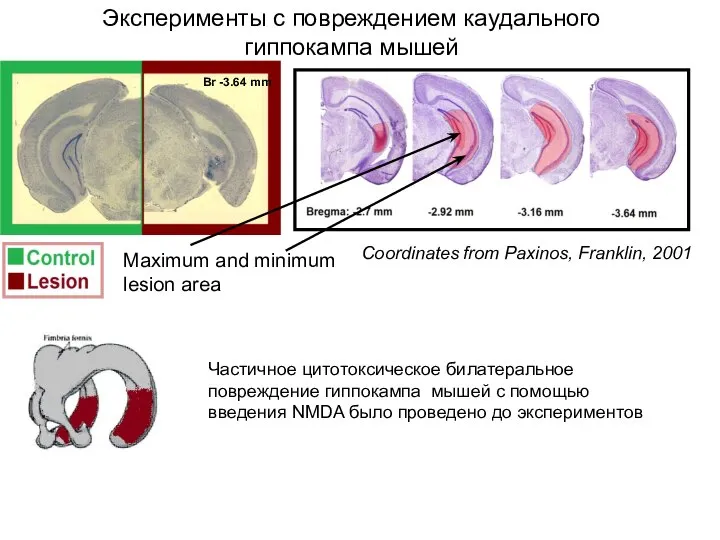
- 111. Частичное цитотоксическое билатеральное повреждение гиппокампа мышей с помощью введения NMDA было проведено до экспериментов Эксперименты с
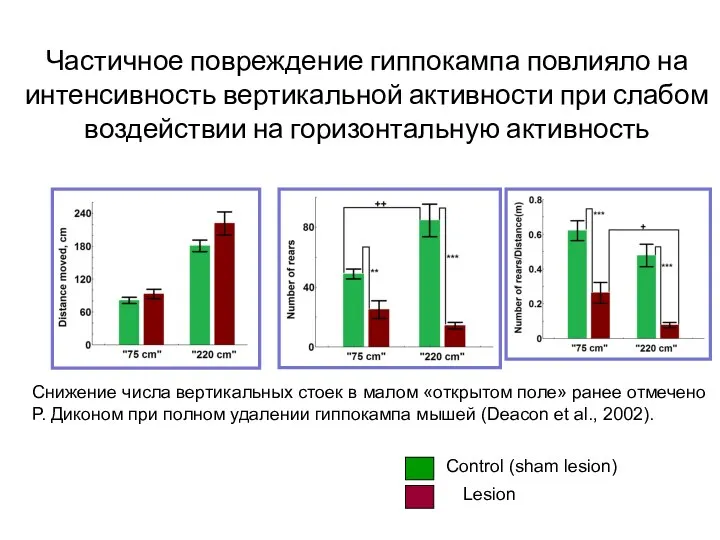
- 112. Частичное повреждение гиппокампа повлияло на интенсивность вертикальной активности при слабом воздействии на горизонтальную активность Снижение числа
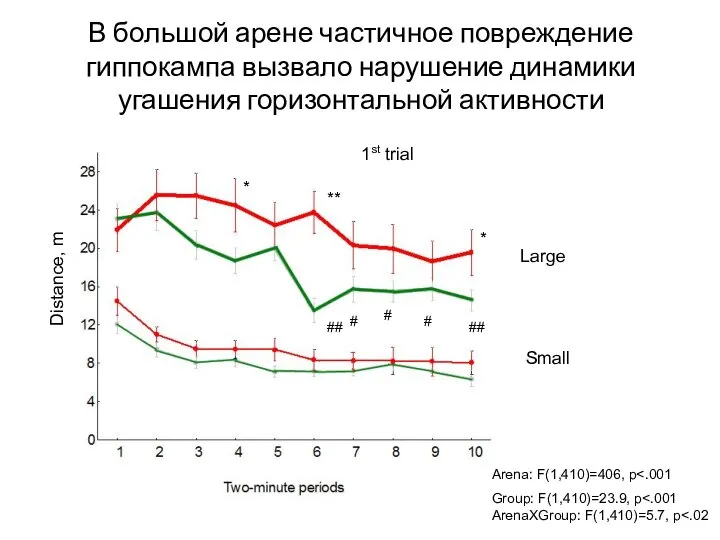
- 113. В большой арене частичное повреждение гиппокампа вызвало нарушение динамики угашения горизонтальной активности 1st trial Arena: F(1,410)=406,
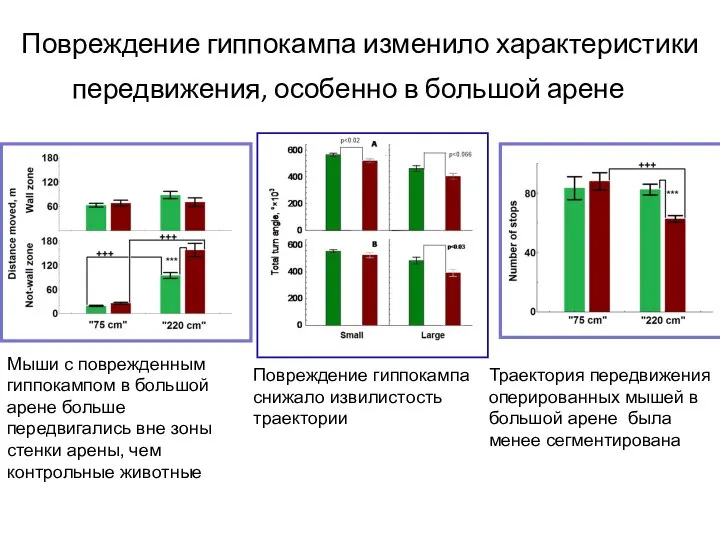
- 114. Повреждение гиппокампа изменило характеристики передвижения, особенно в большой арене Мыши с поврежденным гиппокампом в большой арене
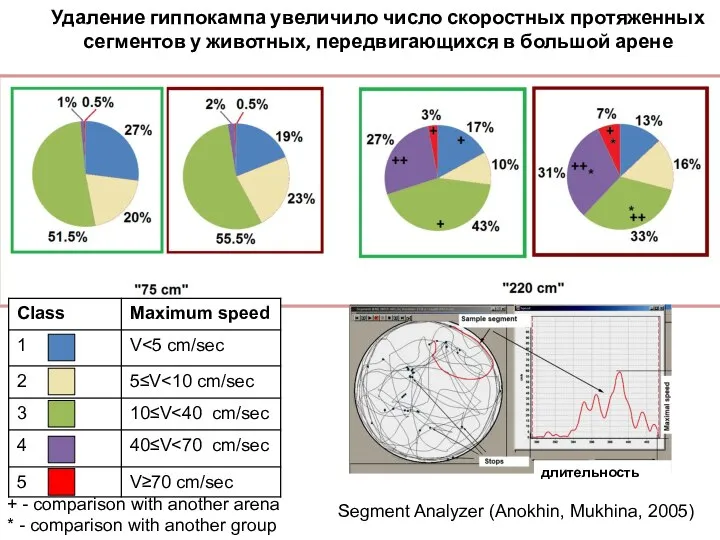
- 115. Удаление гиппокампа увеличило число скоростных протяженных сегментов у животных, передвигающихся в большой арене + - comparison
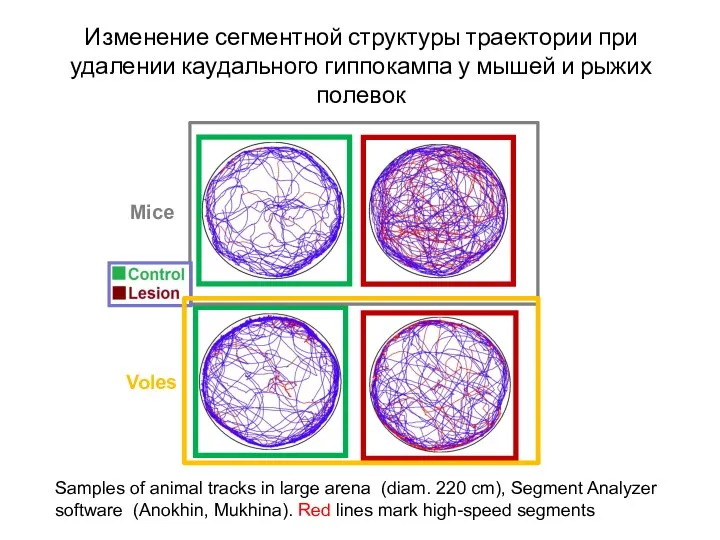
- 116. Mice Voles Samples of animal tracks in large arena (diam. 220 cm), Segment Analyzer software (Anokhin,
- 117. Изменение характеристик сегментов пути после повреждения каудальной части гиппокампа у мышей: увеличивается доля протяженных высокоскоростных сегментов
- 118. Выводы (Bast et al., 2009) Thus, residual circuitry at the septal pole can ‘‘learn’’rapidly, but cannot
- 119. Особенности входов в разные области энторинальной коры Корковые входы в ЭК организованны по «полоскам», особенно в
- 121. Скачать презентацию
Слайд 3Обучение по стабильным зрительным ориентирам, стабильные поля разряда PC, несмотря на изменение
Обучение по стабильным зрительным ориентирам, стабильные поля разряда PC, несмотря на изменение
Слайд 4Дезорганизация полей разряда PC, появление дополнительных полей в области кормушки с целевым
Дезорганизация полей разряда PC, появление дополнительных полей в области кормушки с целевым
Слайд 5Феномен “replay” и память
Что показывает одновременная регистрация активности многих клеток места
Феномен “replay” и память
Что показывает одновременная регистрация активности многих клеток места

Слайд 6Ансамбли клеток поля CA1 гиппокампа, реконструкция траектории передвижения крысы
Поля разрядов ансамбля
Ансамбли клеток поля CA1 гиппокампа, реконструкция траектории передвижения крысы
Поля разрядов ансамбля
Слайд 7Так как клетки места гиппокампа активируются в определенных местах и в определенном
Так как клетки места гиппокампа активируются в определенных местах и в определенном


Слайд 9“Replay” – последовательное, организованное по времени повторное «проигрывание» активности группы нейронов гиппокампа
“Replay” – последовательное, организованное по времени повторное «проигрывание» активности группы нейронов гиппокампа

Слайд 10Трек крысы (развертка по времени), красным отмечено поле разряда одной клетки места
Поля
Трек крысы (развертка по времени), красным отмечено поле разряда одной клетки места
Поля
Слайд 11Reverse replay
Порядок разряда клеток обратный тому, что наблюдался при передвижении животного
Бег
Reverse replay
Порядок разряда клеток обратный тому, что наблюдался при передвижении животного
Бег
Слайд 12ripple
Животное передвигается по линейному треку, 13 клеток места последовательно активизируются. Replay (в
ripple
Животное передвигается по линейному треку, 13 клеток места последовательно активизируются. Replay (в
Слайд 13Функциональная неоднородность гиппокампа вдоль септотемпоральной (рострокаудальной для грызунов) оси
Функциональная неоднородность гиппокампа вдоль септотемпоральной (рострокаудальной для грызунов) оси

Слайд 14Септальный полюс (ростральный =дорзальный гиппокамп)
Височный полюс (каудальный=вентральный гипокамп)
Гиппокамп вдоль рострокаудальной оси неоднороден
Септальный полюс (ростральный =дорзальный гиппокамп)
Височный полюс (каудальный=вентральный гипокамп)
Гиппокамп вдоль рострокаудальной оси неоднороден
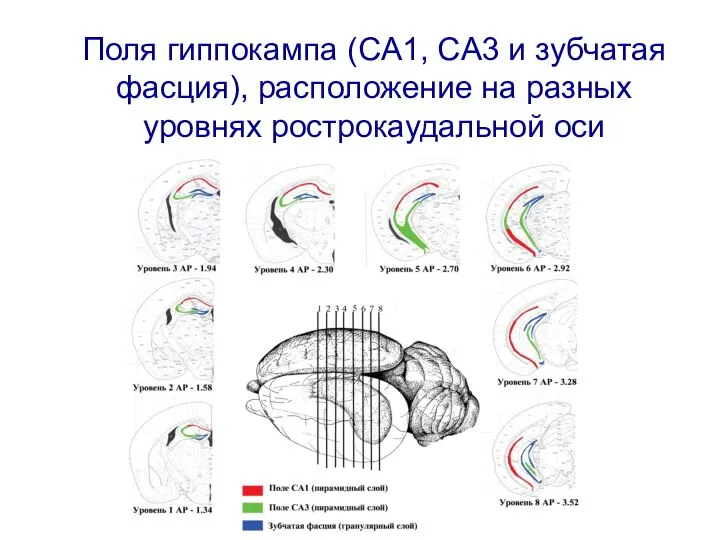
Слайд 15Поля гиппокампа (CA1, CA3 и зубчатая фасция), расположение на разных уровнях рострокаудальной
Поля гиппокампа (CA1, CA3 и зубчатая фасция), расположение на разных уровнях рострокаудальной
Слайд 16Молекулярные домены гиппокампа
Вид с латеральной стороны
Вид с медиальной стороны
Fanselow, Dong, 2010
C
Молекулярные домены гиппокампа
Вид с латеральной стороны
Вид с медиальной стороны
Fanselow, Dong, 2010
C
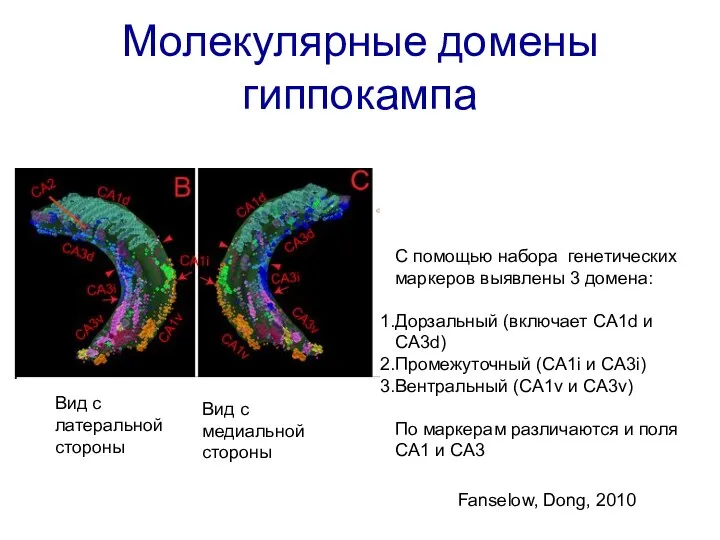
Слайд 17Молекулярные домены гиппокампа
Fanselow, Dong, 2010
C помощью набора генетических маркеров выявлены 3
Молекулярные домены гиппокампа
Fanselow, Dong, 2010
C помощью набора генетических маркеров выявлены 3

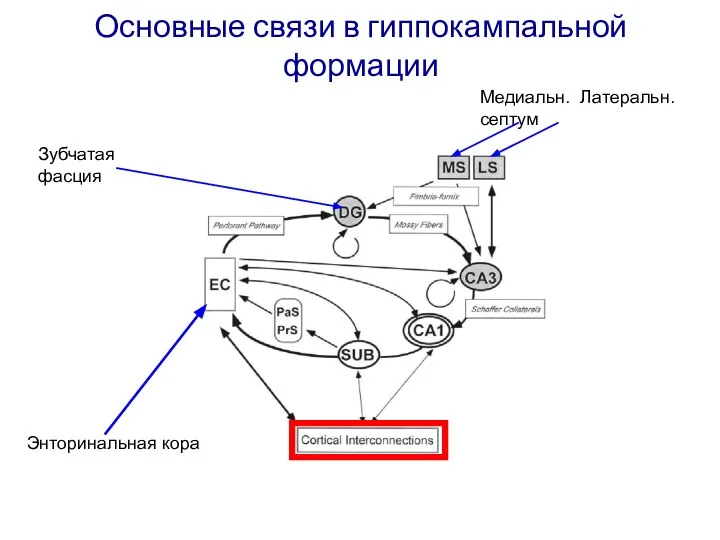
Слайд 18Основные связи в гиппокампальной формации
Зубчатая фасция
Медиальн. Латеральн. септум
Энторинальная кора
Основные связи в гиппокампальной формации
Зубчатая фасция
Медиальн. Латеральн. септум
Энторинальная кора
Слайд 19Парагиппокампальная область
PER – периринальная кора, POR – постринальная кора, EC – энторинальная
Парагиппокампальная область
PER – периринальная кора, POR – постринальная кора, EC – энторинальная

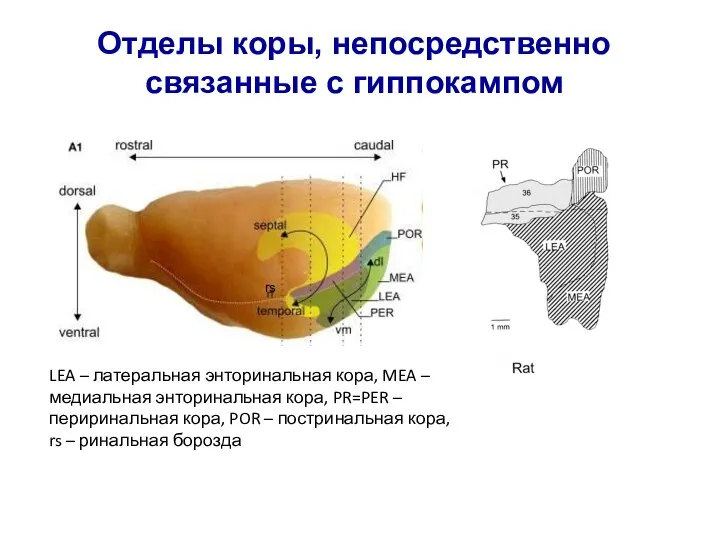
Слайд 20LEA – латеральная энторинальная кора, MEA – медиальная энторинальная кора, PR=PER –
LEA – латеральная энторинальная кора, MEA – медиальная энторинальная кора, PR=PER –
Слайд 21Hartman, Lever, Burgess, O’Keefe, 2014
Hartman, Lever, Burgess, O’Keefe, 2014
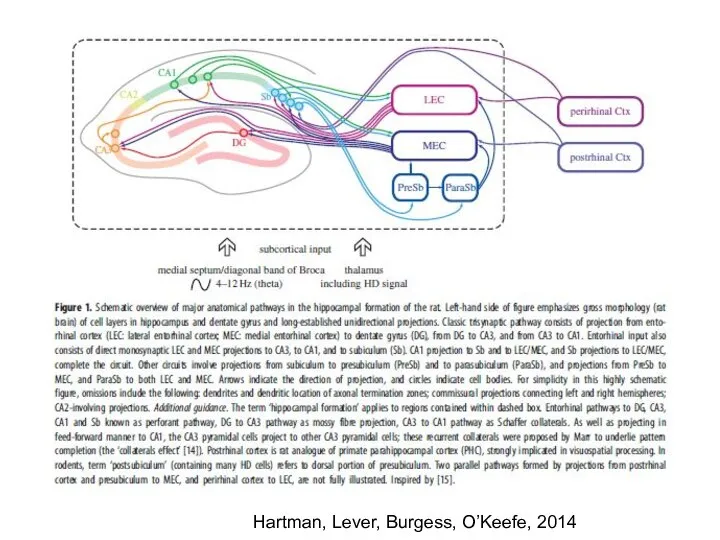
Слайд 22Топология проекций энторинальной коры (ЭК) в зубчатую извилину
A – полушария (мозжечок снят),
Топология проекций энторинальной коры (ЭК) в зубчатую извилину
A – полушария (мозжечок снят),

Слайд 23Функциональная неоднородность гиппокампа
Bast, 2007
Функциональная неоднородность гиппокампа
Bast, 2007
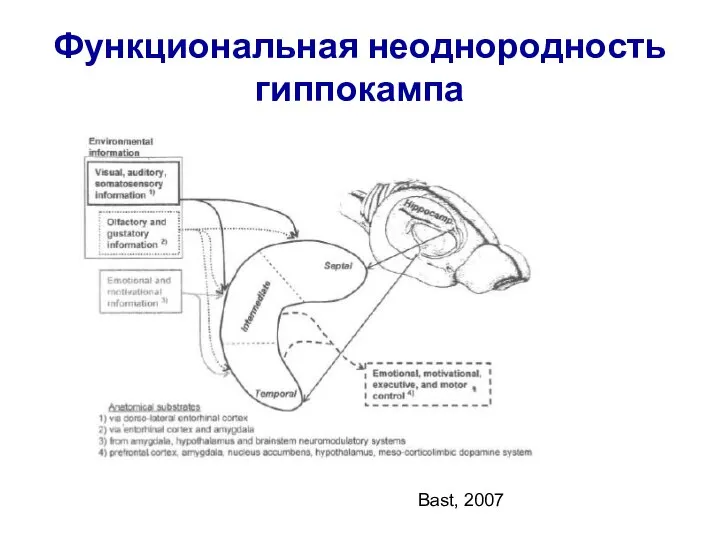
Слайд 24(Bast et al., 2009)
Области септальной и промежуточной части гиппокампа больше связаны с
(Bast et al., 2009)
Области септальной и промежуточной части гиппокампа больше связаны с
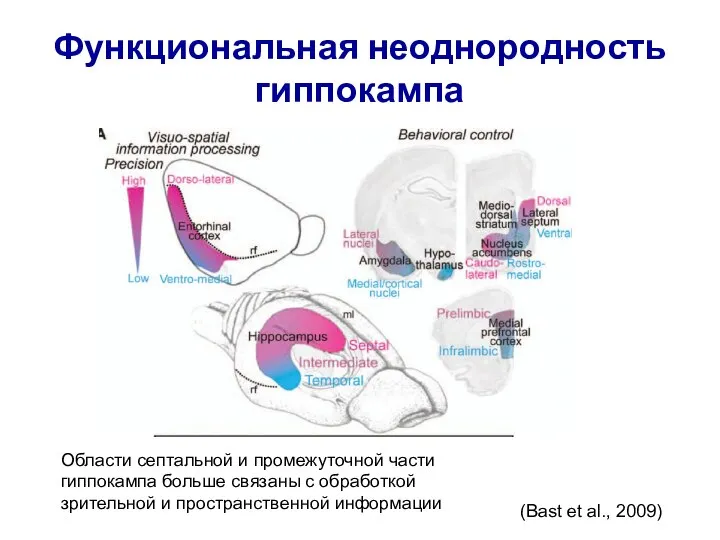
Слайд 25Свойства клеток места в разных субрегионах гиппокампа вдоль септотемпоральной оси
Пространственно-специфичные нейроны обнаружены
Свойства клеток места в разных субрегионах гиппокампа вдоль септотемпоральной оси
Пространственно-специфичные нейроны обнаружены

Слайд 26Клетки места при передвижении по дорожке на большие расстояния
18 м
Kjelstrup et al.,
Клетки места при передвижении по дорожке на большие расстояния
18 м
Kjelstrup et al.,
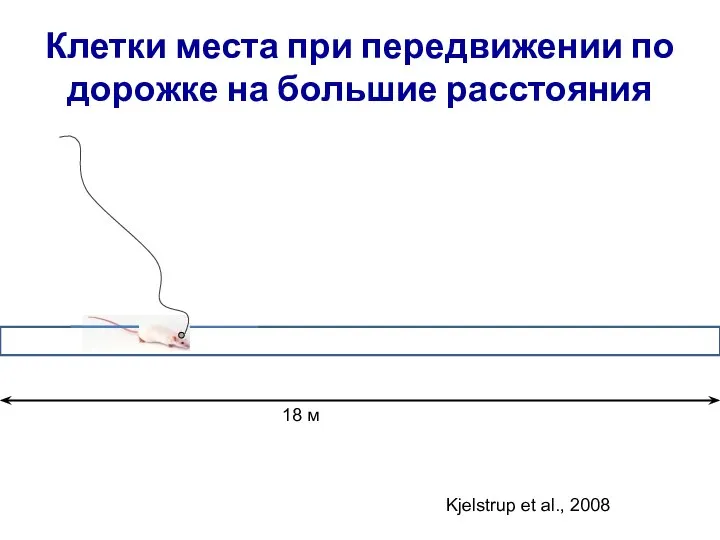
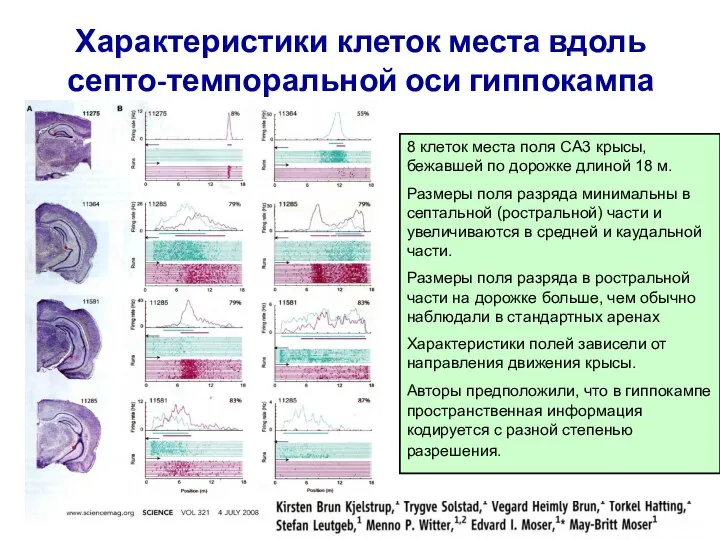
Слайд 27Характеристики клеток места вдоль септо-темпоральной оси гиппокампа
8 клеток места поля CA3 крысы,
Характеристики клеток места вдоль септо-темпоральной оси гиппокампа
8 клеток места поля CA3 крысы,
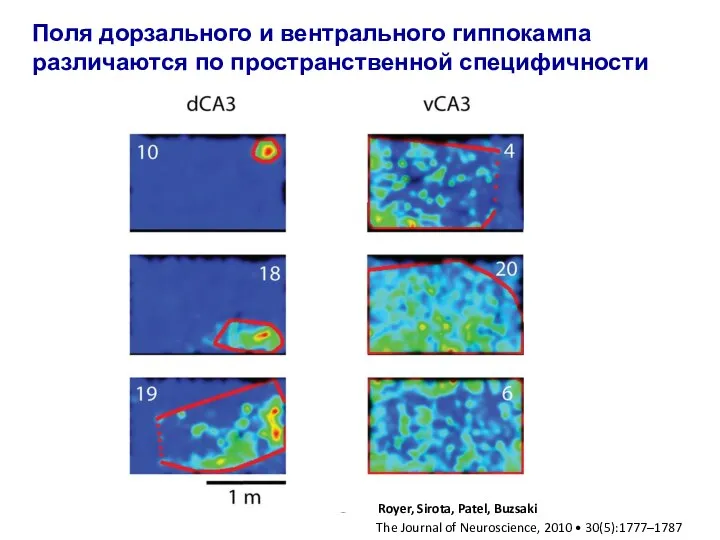
Слайд 28Поля дорзального и вентрального гиппокампа различаются по пространственной специфичности
Поля дорзального и вентрального гиппокампа различаются по пространственной специфичности
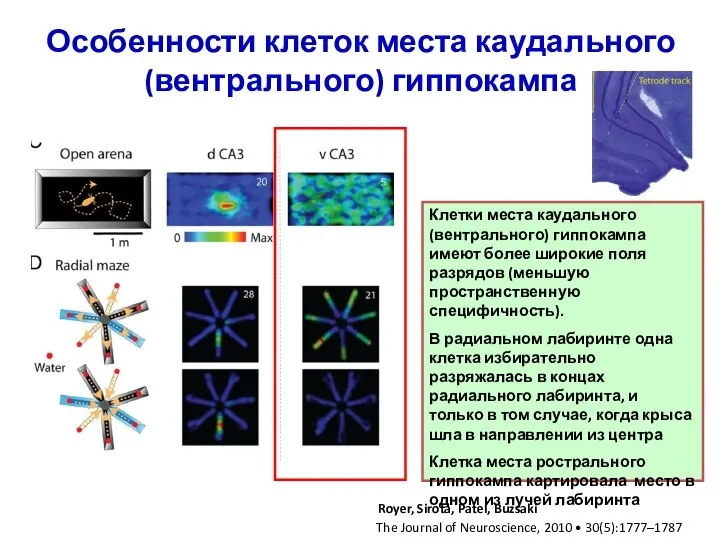
Слайд 29Особенности клеток места каудального (вентрального) гиппокампа
Клетки места каудального (вентрального) гиппокампа имеют более
Особенности клеток места каудального (вентрального) гиппокампа
Клетки места каудального (вентрального) гиппокампа имеют более
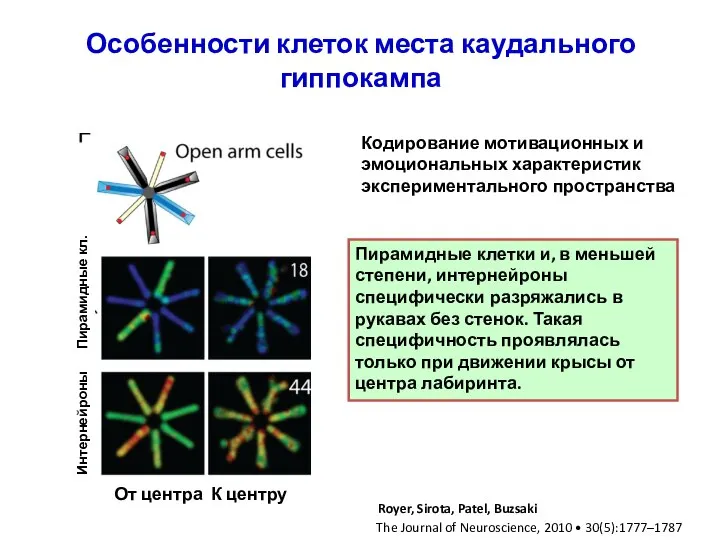
Слайд 30Особенности клеток места каудального гиппокампа
Пирамидные клетки и, в меньшей степени, интернейроны специфически
Особенности клеток места каудального гиппокампа
Пирамидные клетки и, в меньшей степени, интернейроны специфически
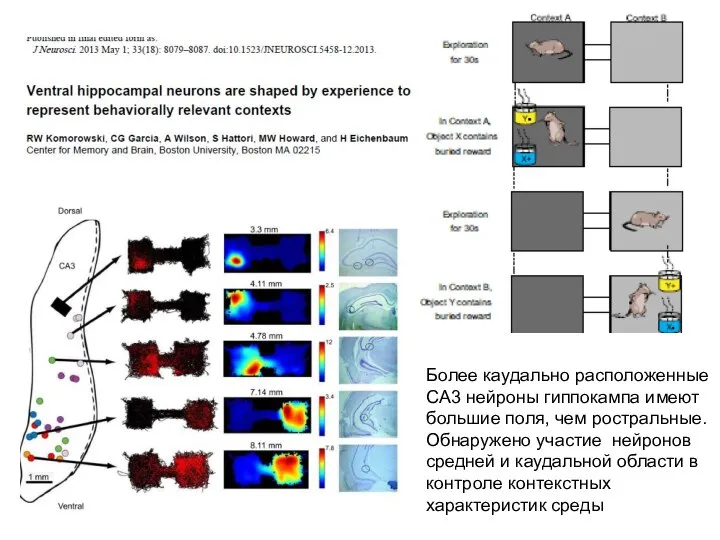
Слайд 31Более каудально расположенные CA3 нейроны гиппокампа имеют большие поля, чем ростральные.
Обнаружено
Более каудально расположенные CA3 нейроны гиппокампа имеют большие поля, чем ростральные.
Обнаружено
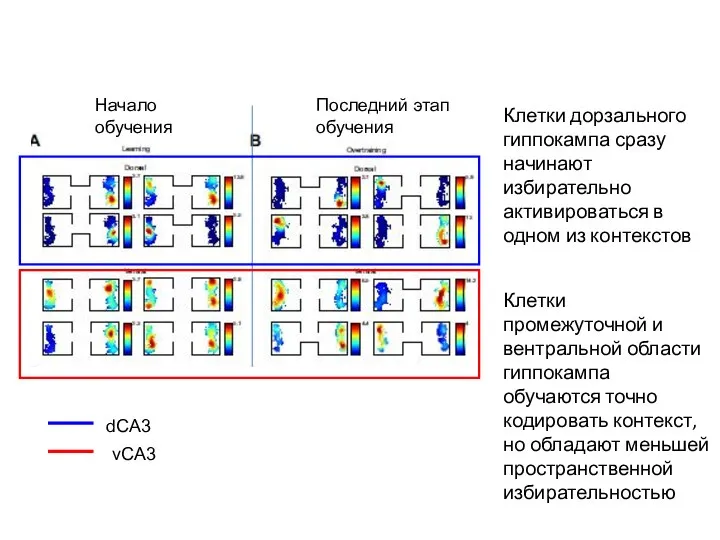
Слайд 32Клетки дорзального гиппокампа сразу начинают избирательно активироваться в одном из контекстов
Клетки промежуточной
Клетки дорзального гиппокампа сразу начинают избирательно активироваться в одном из контекстов
Клетки промежуточной
Слайд 33Исследование функциональной гетерогенности гиппокампа
K.V. Anokhin
R. Deacon (Oxford, UK)
V.M. Malygin
P. A. Kuptsov, лаборатория
Исследование функциональной гетерогенности гиппокампа
K.V. Anokhin
R. Deacon (Oxford, UK)
V.M. Malygin
P. A. Kuptsov, лаборатория

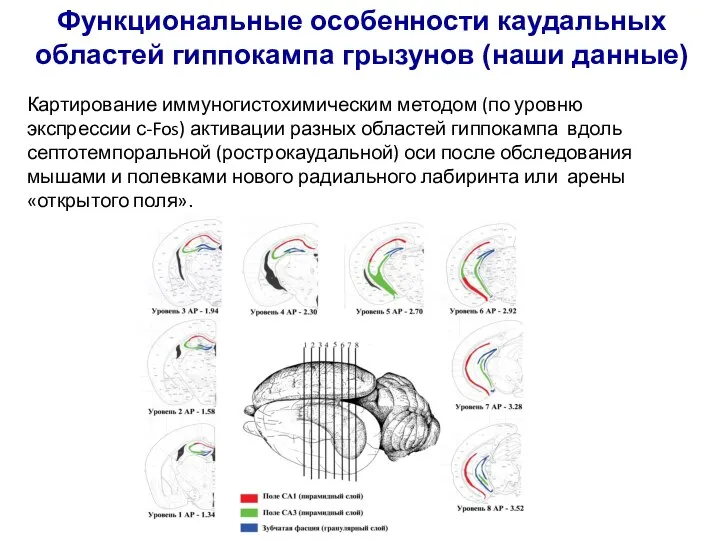
Слайд 34Функциональные особенности каудальных областей гиппокампа грызунов (наши данные)
Картирование иммуногистохимическим методом (по
Функциональные особенности каудальных областей гиппокампа грызунов (наши данные)
Картирование иммуногистохимическим методом (по
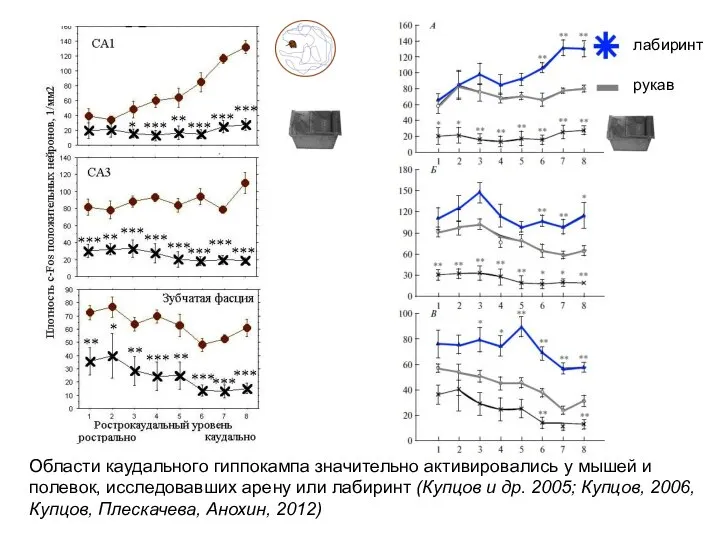
Слайд 35Области каудального гиппокампа значительно активировались у мышей и полевок, исследовавших арену или
Области каудального гиппокампа значительно активировались у мышей и полевок, исследовавших арену или
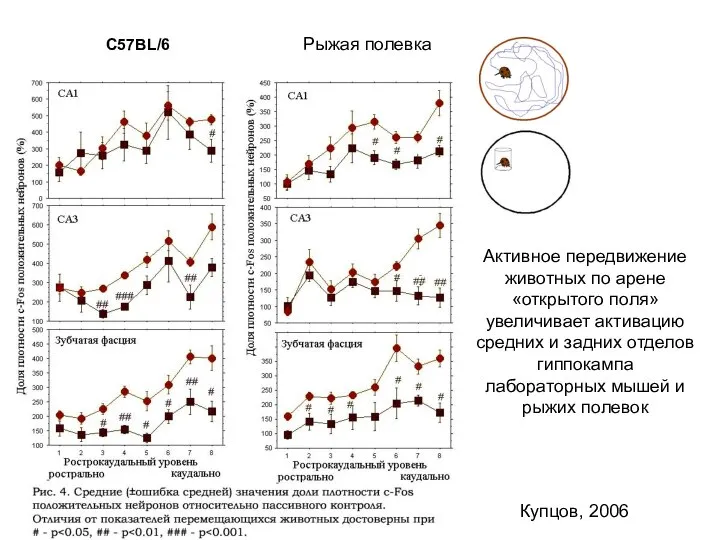
Слайд 36C57BL/6
Рыжая полевка
Активное передвижение животных по арене «открытого поля» увеличивает активацию средних и
C57BL/6
Рыжая полевка
Активное передвижение животных по арене «открытого поля» увеличивает активацию средних и
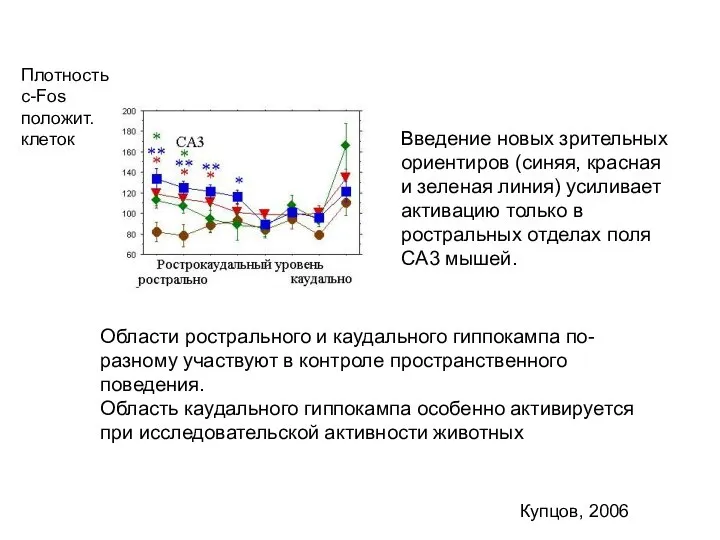
Слайд 37Купцов, 2006
Введение новых зрительных ориентиров (синяя, красная и зеленая линия) усиливает активацию
Купцов, 2006
Введение новых зрительных ориентиров (синяя, красная и зеленая линия) усиливает активацию
Слайд 38Клетки места у представителей других видов
Клетки места у представителей других видов


Слайд 40Клетки места дорзального гиппокампа шиншиллы
Диам. 76 см
Muir et al., 2009
Отр. Грызуны, сем.
Клетки места дорзального гиппокампа шиншиллы
Диам. 76 см
Muir et al., 2009
Отр. Грызуны, сем.

Слайд 41Клетки места у летучей мыши
Ulanovsky, Moss, 2007, 2011
Клетки регистрировали в септальной части
Клетки места у летучей мыши
Ulanovsky, Moss, 2007, 2011
Клетки регистрировали в септальной части

Слайд 42Форма поля разряда клетки места и биология вида
Ulanovsky 2011
Hayman et al., 2011
???
Будет
Форма поля разряда клетки места и биология вида
Ulanovsky 2011
Hayman et al., 2011
???
Будет

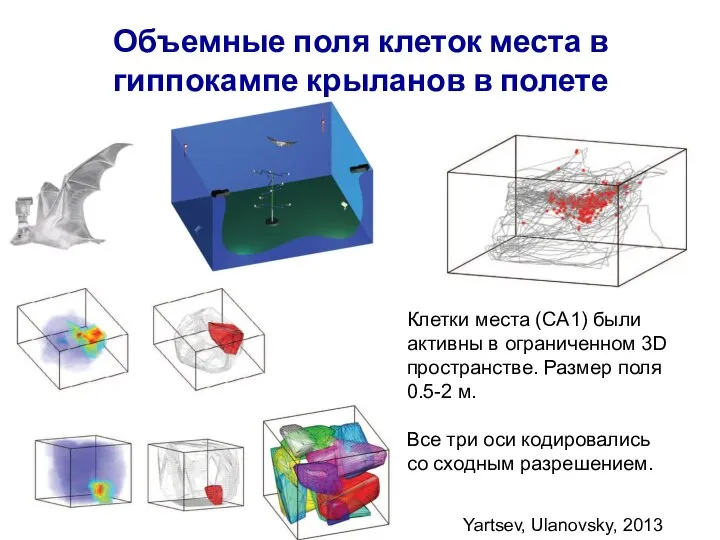
Слайд 43Объемные поля клеток места в гиппокампе крыланов в полете
Yartsev, Ulanovsky, 2013
Клетки места
Объемные поля клеток места в гиппокампе крыланов в полете
Yartsev, Ulanovsky, 2013
Клетки места
Слайд 44Клетки места, зависимость от направления головы (направления движения)
Rubin, Yartsev, Ulanovsky, 2014
Бурый кожан,
Клетки места, зависимость от направления головы (направления движения)
Rubin, Yartsev, Ulanovsky, 2014
Бурый кожан,

Слайд 45Клетки места, зависимость от направления головы (направления движения)
Rubin, Yartsev, Ulanovsky, 2014
Клетка поля
Клетки места, зависимость от направления головы (направления движения)
Rubin, Yartsev, Ulanovsky, 2014
Клетка поля

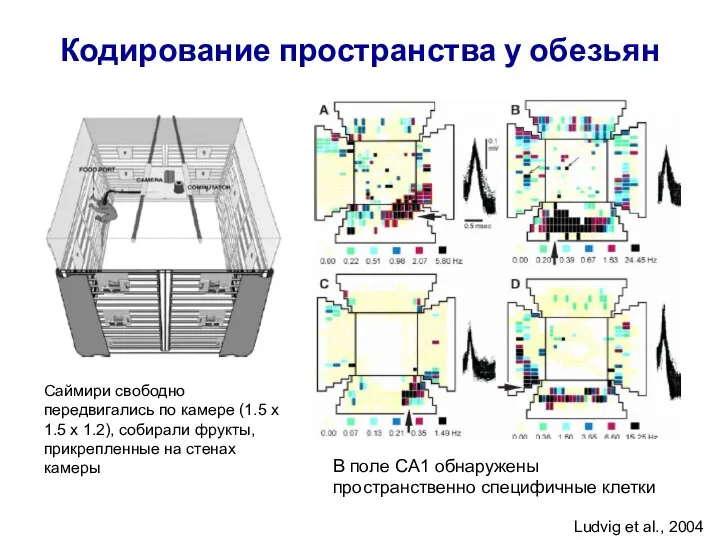
Слайд 46Кодирование пространства у обезьян
Саймири свободно передвигались по камере (1.5 х 1.5 х
Кодирование пространства у обезьян
Саймири свободно передвигались по камере (1.5 х 1.5 х
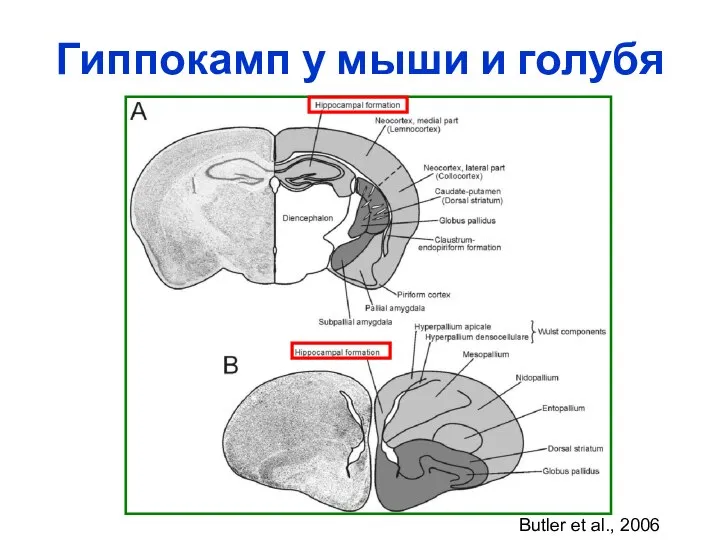
Слайд 47Гиппокамп у мыши и голубя
Butler et al., 2006
Гиппокамп у мыши и голубя
Butler et al., 2006
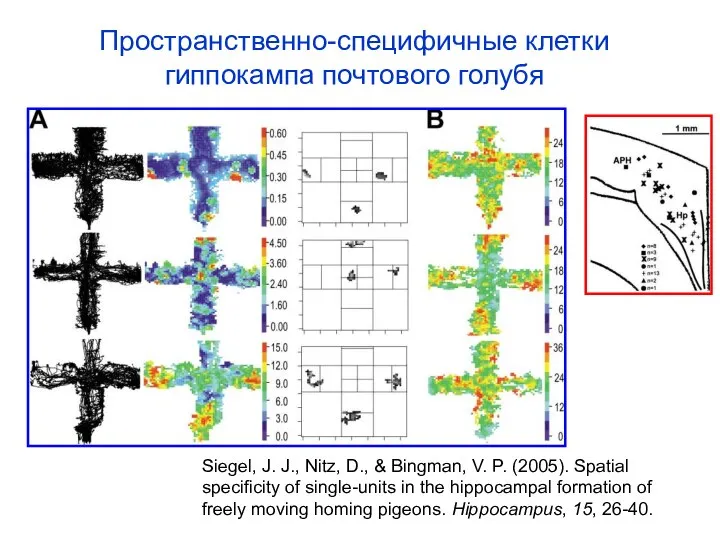
Слайд 48Пространственно-специфичные клетки гиппокампа почтового голубя
Siegel, J. J., Nitz, D., & Bingman, V.
Пространственно-специфичные клетки гиппокампа почтового голубя
Siegel, J. J., Nitz, D., & Bingman, V.
Слайд 49Человек
(пациенты с вживленными по медицинским показаниям электродами в область эпилептического очага)
Путешествие
Человек
(пациенты с вживленными по медицинским показаниям электродами в область эпилептического очага)
Путешествие

Слайд 50Области мозга, где обнаружены пространственно специфичные клетки
Гиппокамп (Аммонов рог, зубчатая фасция)
Области мозга, где обнаружены пространственно специфичные клетки
Гиппокамп (Аммонов рог, зубчатая фасция)

Слайд 51Для успешной навигации в пространстве необходимо не только определение своего местоположения, но
Для успешной навигации в пространстве необходимо не только определение своего местоположения, но

Слайд 52Клетки направления головы (head direction cells, HD cells)
Клетки направления головы (head direction cells, HD cells)

Слайд 53Парагиппокампальная область
PER – периринальная кора, POR – постринальная кора, EC – энторинальная
Парагиппокампальная область
PER – периринальная кора, POR – постринальная кора, EC – энторинальная

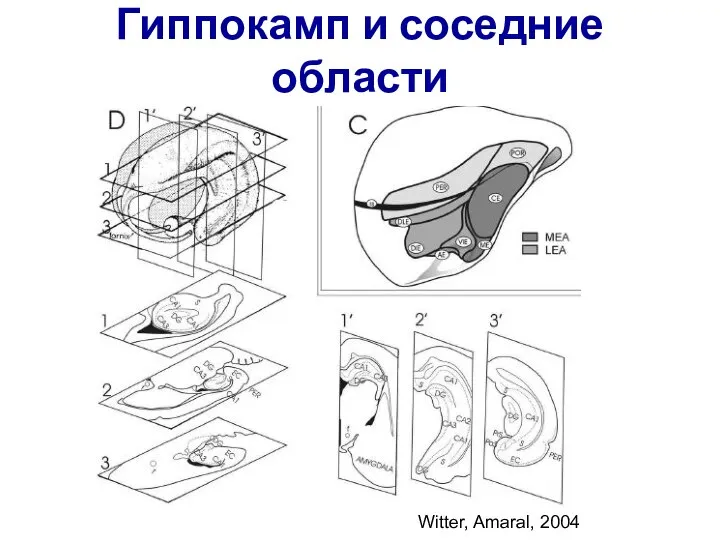
Слайд 54Гиппокамп и соседние области
Witter, Amaral, 2004
Гиппокамп и соседние области
Witter, Amaral, 2004
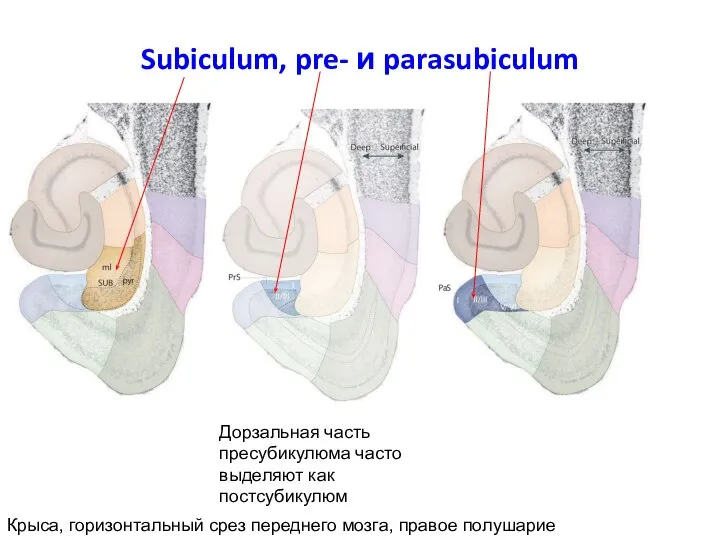
Слайд 55Subiculum, pre- и parasubiculum
Крыса, горизонтальный срез переднего мозга, правое полушарие
Дорзальная часть пресубикулюма
Subiculum, pre- и parasubiculum
Крыса, горизонтальный срез переднего мозга, правое полушарие
Дорзальная часть пресубикулюма
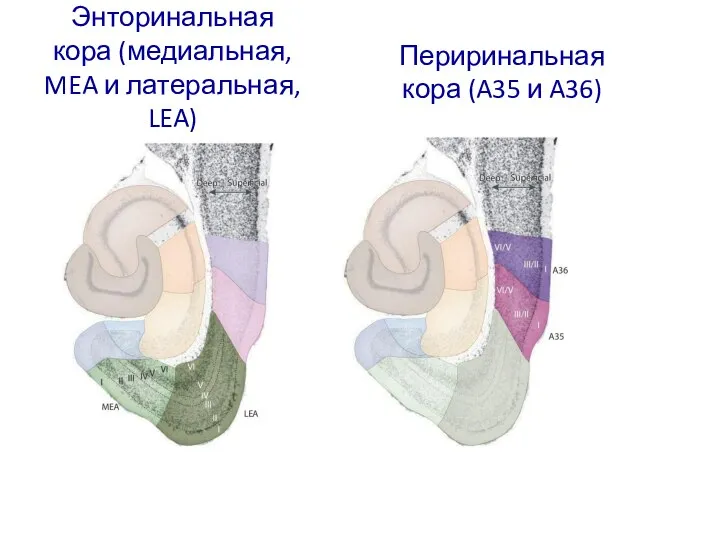
Слайд 56Энторинальная кора (медиальная, MEA и латеральная, LEA)
Периринальная кора (A35 и A36)
Энторинальная кора (медиальная, MEA и латеральная, LEA)
Периринальная кора (A35 и A36)
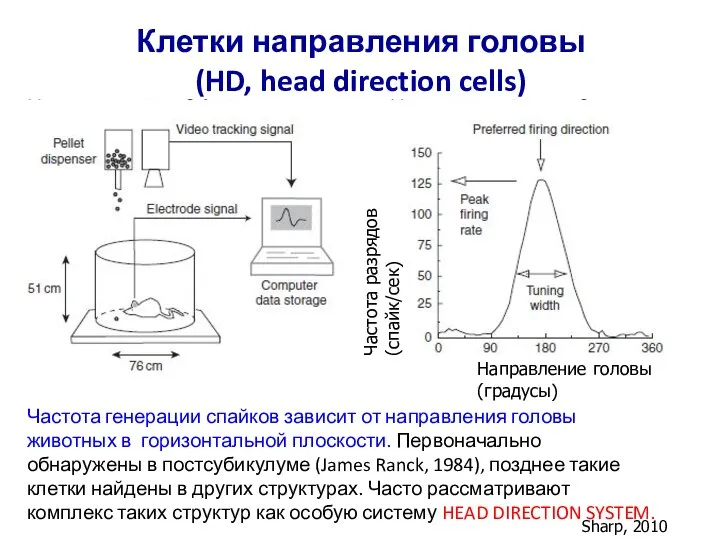
Слайд 57Клетки направления головы
(HD, head direction cells)
Sharp, 2010
Частота генерации спайков зависит от
Клетки направления головы
(HD, head direction cells)
Sharp, 2010
Частота генерации спайков зависит от
Слайд 58HD клетки, активность. Видео
Taube, 2007
HD клетки, активность. Видео
Taube, 2007

Слайд 59Jeffrey Taube
Department of Psychological and Brain Sciences at Dartmouth
2005
Jeffrey Taube
Department of Psychological and Brain Sciences at Dartmouth
2005

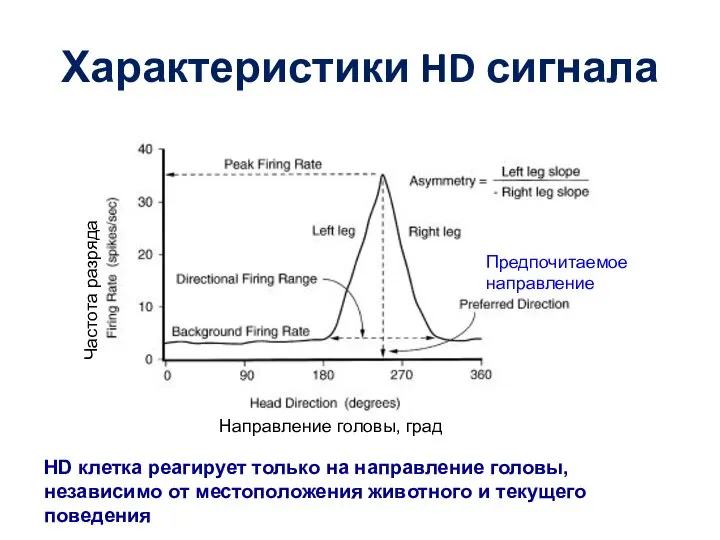
Слайд 60Характеристики HD сигнала
Предпочитаемое направление
Направление головы, град
Частота разряда
HD клетка реагирует только на направление
Характеристики HD сигнала
Предпочитаемое направление
Направление головы, град
Частота разряда
HD клетка реагирует только на направление
Слайд 61HD клетки сначала были обнаружены в постсубикулуме, позже найдены в переднем дорзальном
HD клетки сначала были обнаружены в постсубикулуме, позже найдены в переднем дорзальном

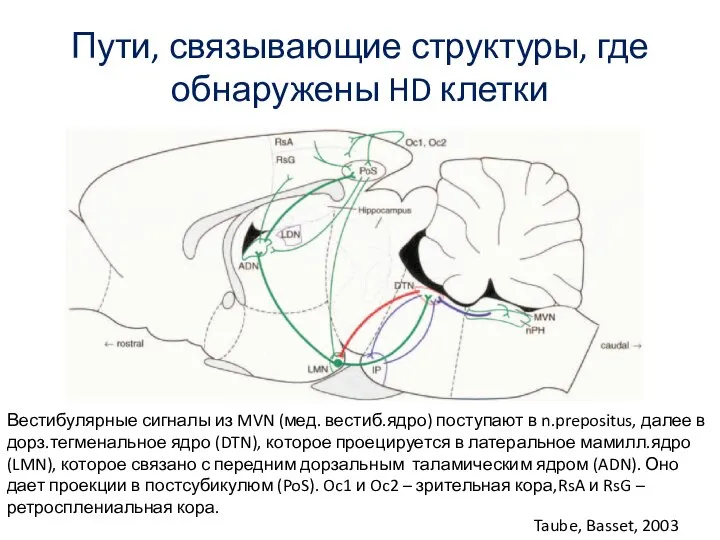
Слайд 62Пути, связывающие структуры, где обнаружены HD клетки
Taube, Basset, 2003
Вестибулярные сигналы из
Пути, связывающие структуры, где обнаружены HD клетки
Taube, Basset, 2003
Вестибулярные сигналы из
Слайд 63Доля клеток, демонстрирующих HD свойста, в разных структурах мозга
Наиболее HD клетки представлены
Доля клеток, демонстрирующих HD свойста, в разных структурах мозга
Наиболее HD клетки представлены

Слайд 64Характеристики клеток направления головы в разных структурах мозга
Taube, Basset, 2003
Характеристики HD
Характеристики клеток направления головы в разных структурах мозга
Taube, Basset, 2003
Характеристики HD
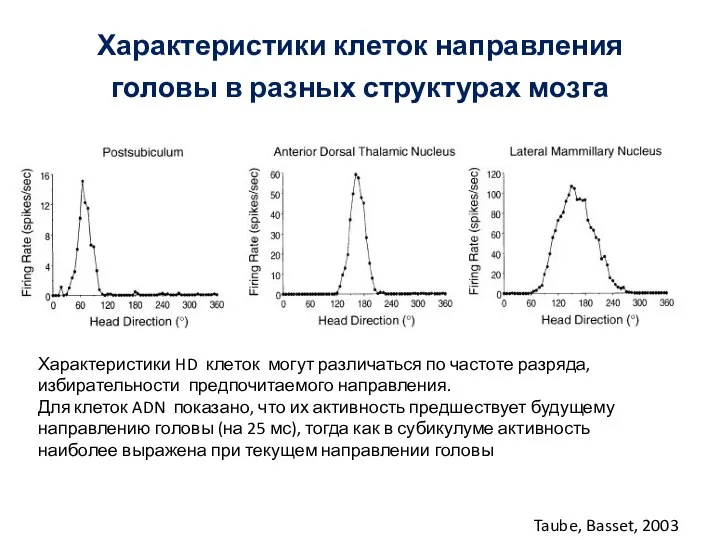
Слайд 65Различия HD клеток в разных структурах мозга
В таламусе, в отличие от постсубикулума,
Различия HD клеток в разных структурах мозга
В таламусе, в отличие от постсубикулума,
Слайд 66Зрительная информация и HD клетки
Taube, 2007
Изменение положения или исчезновение зрительного ориентира вызывает
Зрительная информация и HD клетки
Taube, 2007
Изменение положения или исчезновение зрительного ориентира вызывает
Слайд 67Влияние ограничения зрительной информации на предпочитаемое направление HD клетки
Goodridge et al., 1998
Влияние ограничения зрительной информации на предпочитаемое направление HD клетки
Goodridge et al., 1998
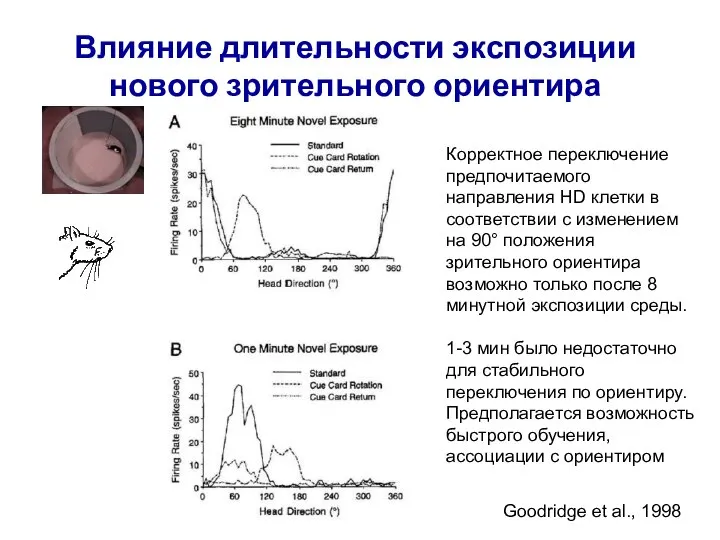
Слайд 68Влияние длительности экспозиции нового зрительного ориентира
Goodridge et al., 1998
Корректное переключение предпочитаемого направления
Влияние длительности экспозиции нового зрительного ориентира
Goodridge et al., 1998
Корректное переключение предпочитаемого направления
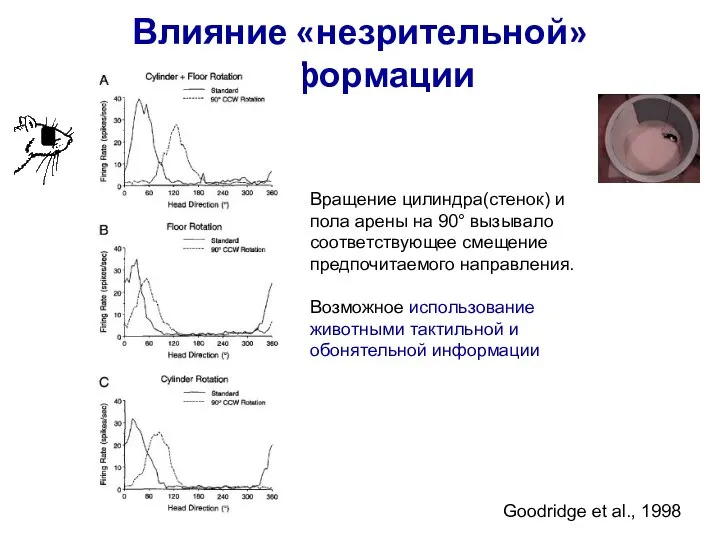
Слайд 69Влияние «незрительной» информации
Вращение цилиндра(стенок) и пола арены на 90° вызывало соответствующее смещение
Влияние «незрительной» информации
Вращение цилиндра(стенок) и пола арены на 90° вызывало соответствующее смещение
Слайд 70Влияние звука
Goodridge et al., 1998
Эксперименты проводили в темноте, через колонку подавали звуковой
Влияние звука
Goodridge et al., 1998
Эксперименты проводили в темноте, через колонку подавали звуковой

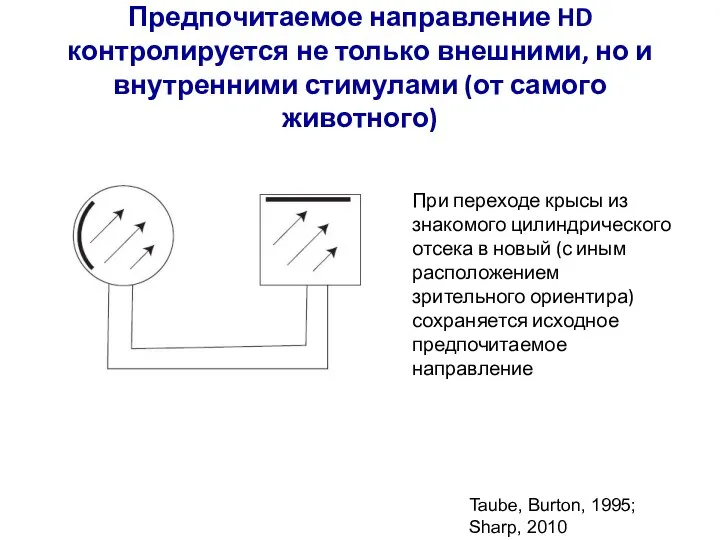
Слайд 71Предпочитаемое направление HD контролируется не только внешними, но и внутренними стимулами (от
Предпочитаемое направление HD контролируется не только внешними, но и внутренними стимулами (от
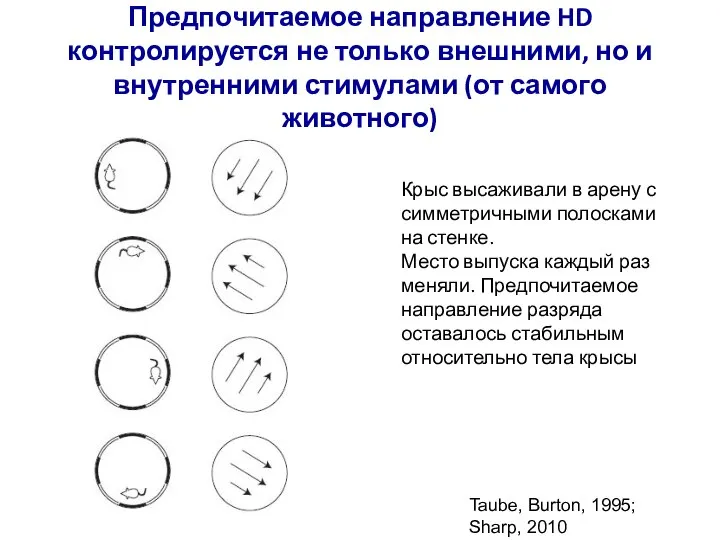
Слайд 72Предпочитаемое направление HD контролируется не только внешними, но и внутренними стимулами (от
Предпочитаемое направление HD контролируется не только внешними, но и внутренними стимулами (от
Слайд 73Характеристики HD клеток
Разнообразные сенсорные сигналы могут определять предпочитаемое направление и изменять
Характеристики HD клеток
Разнообразные сенсорные сигналы могут определять предпочитаемое направление и изменять

Слайд 74Эффекты повреждений вестибулярной системы на HD клетки
Sodium arsanilate Eliminated HD signal
Эффекты повреждений вестибулярной системы на HD клетки
Sodium arsanilate Eliminated HD signal

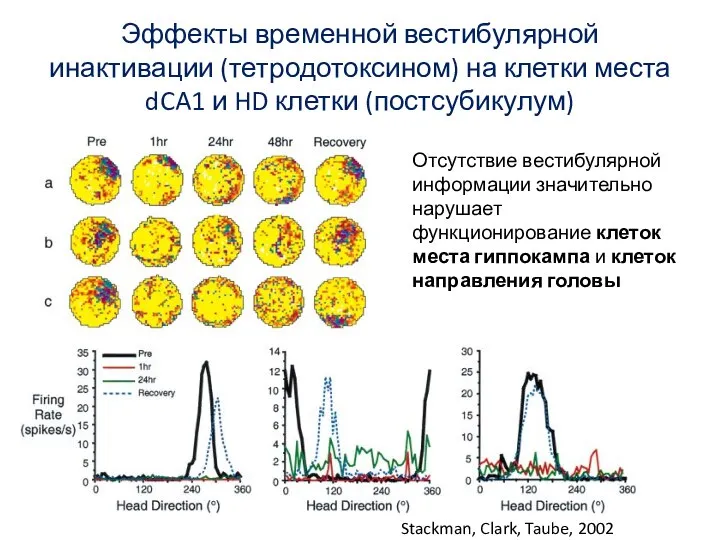
Слайд 75Эффекты временной вестибулярной инактивации (тетродотоксином) на клетки места dCA1 и HD клетки
Эффекты временной вестибулярной инактивации (тетродотоксином) на клетки места dCA1 и HD клетки
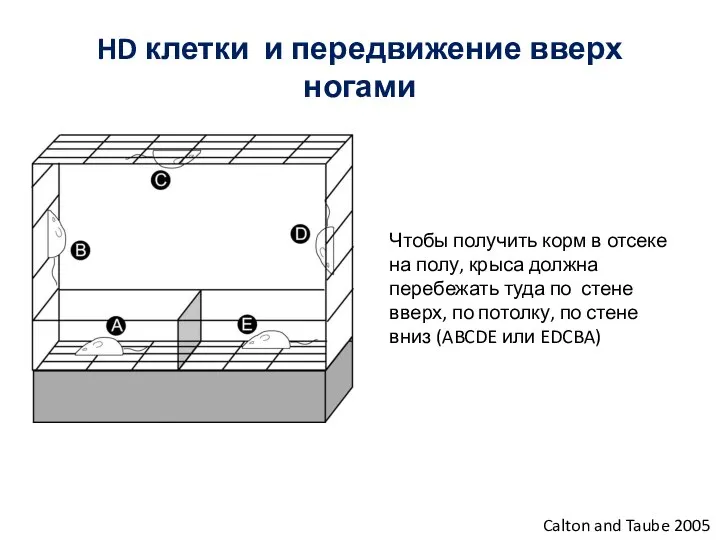
Слайд 76HD клетки и передвижение вверх ногами
Calton and Taube 2005
Чтобы получить корм в
HD клетки и передвижение вверх ногами
Calton and Taube 2005
Чтобы получить корм в
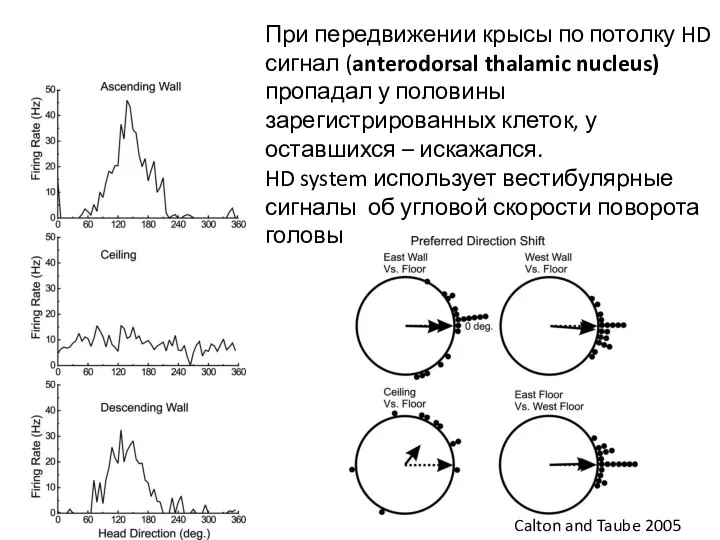
Слайд 77При передвижении крысы по потолку HD сигнал (anterodorsal thalamic nucleus) пропадал у
При передвижении крысы по потолку HD сигнал (anterodorsal thalamic nucleus) пропадал у
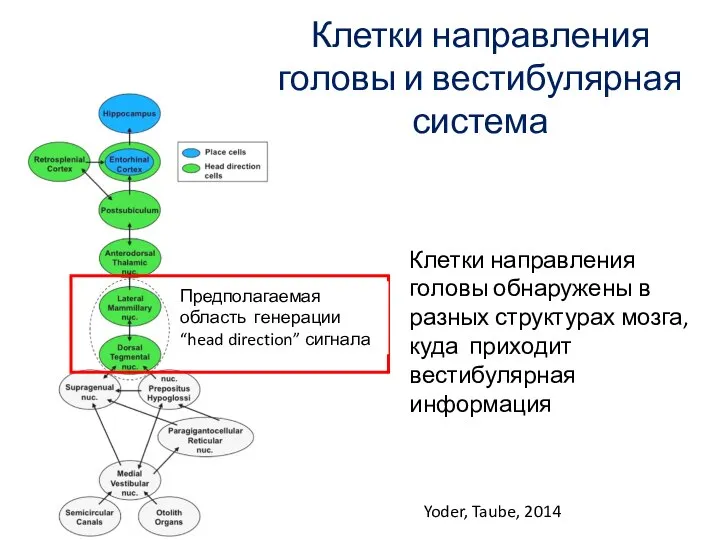
Слайд 78Клетки направления головы и вестибулярная система
Yoder, Taube, 2014
Клетки направления головы обнаружены в
Клетки направления головы и вестибулярная система
Yoder, Taube, 2014
Клетки направления головы обнаружены в
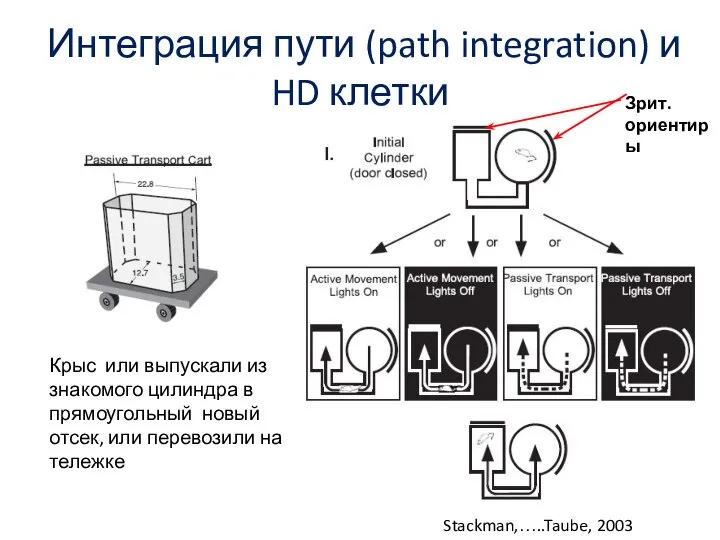
Слайд 79Интеграция пути (path integration) и HD клетки
Stackman,…..Taube, 2003
Зрит. ориентиры
Крыс или выпускали
Интеграция пути (path integration) и HD клетки
Stackman,…..Taube, 2003
Зрит. ориентиры
Крыс или выпускали
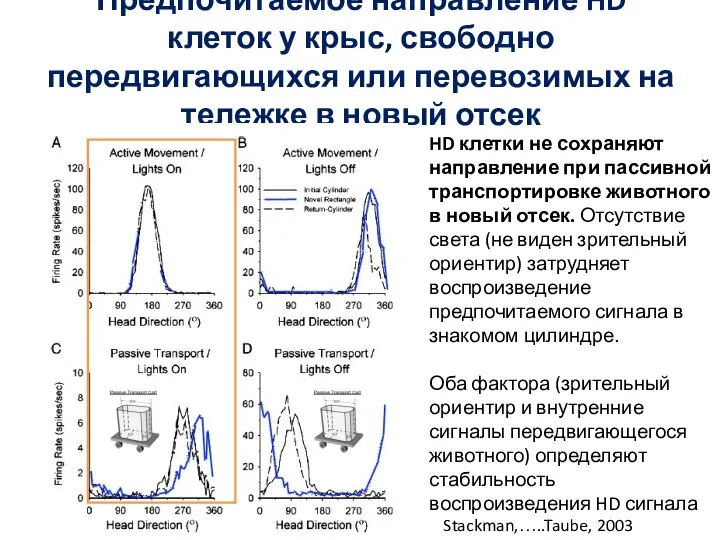
Слайд 80Предпочитаемое направление HD клеток у крыс, свободно передвигающихся или перевозимых на тележке
Предпочитаемое направление HD клеток у крыс, свободно передвигающихся или перевозимых на тележке
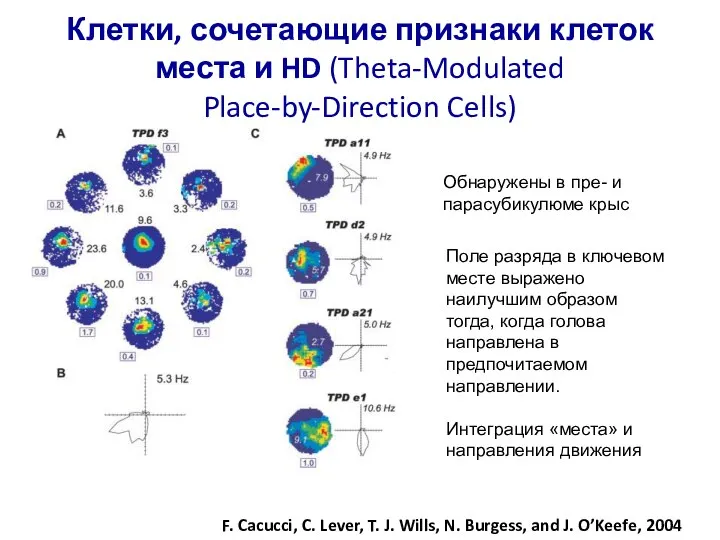
Слайд 81Клетки, сочетающие признаки клеток места и HD (Theta-Modulated Place-by-Direction Cells)
F. Cacucci, C.
Клетки, сочетающие признаки клеток места и HD (Theta-Modulated Place-by-Direction Cells)
F. Cacucci, C.
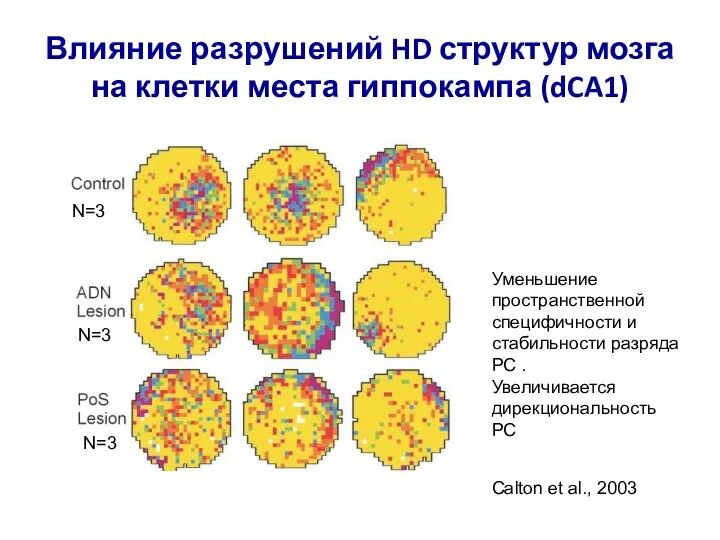
Слайд 82Влияние разрушений HD структур мозга на клетки места гиппокампа (dCA1)
Уменьшение пространственной специфичности
Влияние разрушений HD структур мозга на клетки места гиппокампа (dCA1)
Уменьшение пространственной специфичности
Слайд 83Влияние разрушений HD структур мозга на клетки места гиппокампа (dCA1)
Удаление postsubiculum (PoS)
Влияние разрушений HD структур мозга на клетки места гиппокампа (dCA1)
Удаление postsubiculum (PoS)
Слайд 84Трехмерное HD кодирование в мозге летучей мыши (Rousettus aegyptiacus)
Направление на горизонтальной плоскости
Трехмерное HD кодирование в мозге летучей мыши (Rousettus aegyptiacus)
Направление на горизонтальной плоскости
Слайд 85Типы HD клеток у летучих мышей
Активность зависит только от наклона над горизонтальной
Типы HD клеток у летучих мышей
Активность зависит только от наклона над горизонтальной

Слайд 86«HD cells are similar to a compass in that their discharge is
«HD cells are similar to a compass in that their discharge is

Слайд 87Спасибо за внимание
Спасибо за внимание

Слайд 88Основные анатомические связи гиппокампа и парагиппокампальной области
Witter, 2010
PER и POR – периринальная
Основные анатомические связи гиппокампа и парагиппокампальной области
Witter, 2010
PER и POR – периринальная
Слайд 89Изменение предпочитаемого направления HD постсубикулума на 90 градусов при повороте зрительного ориентира
Изменение предпочитаемого направления HD постсубикулума на 90 градусов при повороте зрительного ориентира
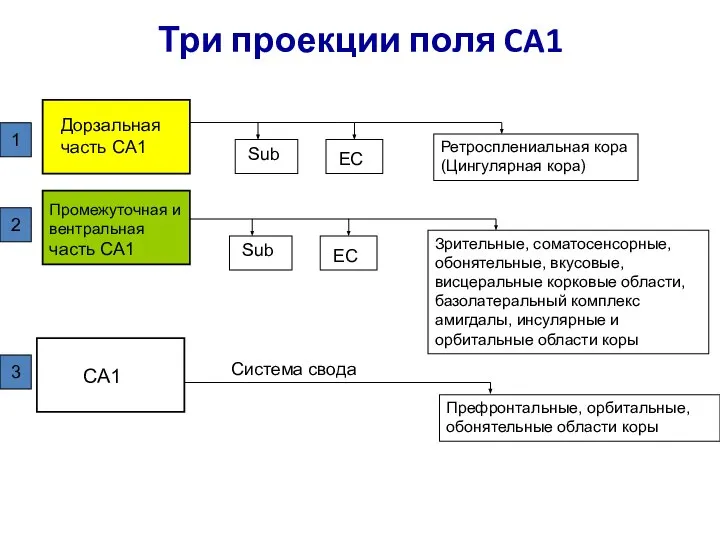
Слайд 90Три проекции поля CA1
Дорзальная часть CA1
Промежуточная и вентральная часть CA1
CA1
Sub
EC
Ретросплениальная кора
Три проекции поля CA1
Дорзальная часть CA1
Промежуточная и вентральная часть CA1
CA1
Sub
EC
Ретросплениальная кора
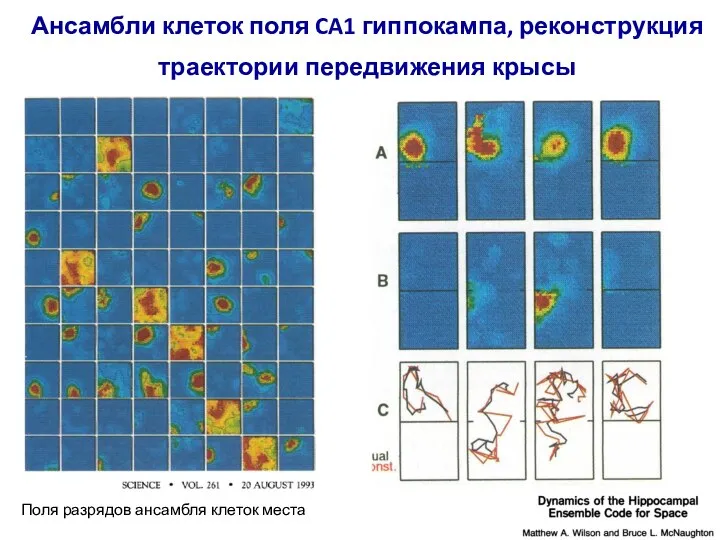
Слайд 91Ансамбли клеток поля CA1 гиппокампа, реконструкция траектории передвижения крысы
Поля разрядов ансамбля
Ансамбли клеток поля CA1 гиппокампа, реконструкция траектории передвижения крысы
Поля разрядов ансамбля
Слайд 92Краткое содержание предыдущей серии…. Ч.1
1. Эдвард Толмен: идея когнитивной карты
1.1 Ментальное
Краткое содержание предыдущей серии…. Ч.1
1. Эдвард Толмен: идея когнитивной карты
1.1 Ментальное

Слайд 93Когнитивные карты. Представления Толмена
В процессе обучения в мозге крысы образуется нечто подобное
Когнитивные карты. Представления Толмена
В процессе обучения в мозге крысы образуется нечто подобное

Слайд 94Краткое содержание предыдущей серии…. Ч.2
2.3 Исследовательская активность – необходимый компонент построения
Краткое содержание предыдущей серии…. Ч.2
2.3 Исследовательская активность – необходимый компонент построения

Слайд 95Place representation как часть когнитивной карты
Строятся с использованием двух типов стимулов :
На
Place representation как часть когнитивной карты
Строятся с использованием двух типов стимулов :
На

Слайд 97Определение понятия «карты» по O’Keefe, Nadel,1978
The simplest definition of a map is
Определение понятия «карты» по O’Keefe, Nadel,1978
The simplest definition of a map is

Слайд 98Когнитивные карты
(O’Keefe, Nadel, 1978)
«Когда животное двигается, «внутренняя навигационная» система будет переключать
Когнитивные карты
(O’Keefe, Nadel, 1978)
«Когда животное двигается, «внутренняя навигационная» система будет переключать

Слайд 99История экспериментального изучения функций мозга
Jean Pierre Flourens (1794-1867)
Франция
Обнаружил анестезирующий эффект хлороформа
В 1825
История экспериментального изучения функций мозга
Jean Pierre Flourens (1794-1867)
Франция
Обнаружил анестезирующий эффект хлороформа
В 1825

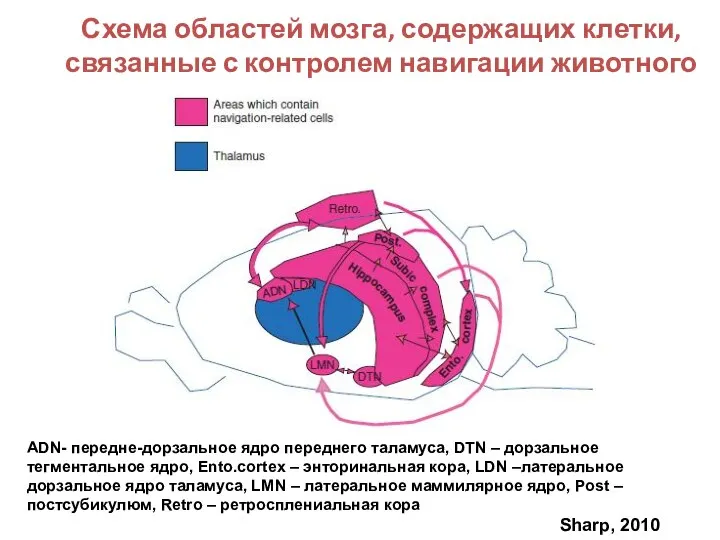
Слайд 100Схема областей мозга, содержащих клетки, связанные с контролем навигации животного
ADN- передне-дорзальное ядро
Схема областей мозга, содержащих клетки, связанные с контролем навигации животного
ADN- передне-дорзальное ядро
Слайд 101Пути, связывающие структуры, где обнаружены HD клетки
Taube, Basset, 2003
Вестибулярные сигналы из
Пути, связывающие структуры, где обнаружены HD клетки
Taube, Basset, 2003
Вестибулярные сигналы из
Слайд 102Навигация и вестибулярная система
Yoder, Taube, 2014
Навигация и вестибулярная система
Yoder, Taube, 2014

Слайд 105Отличия hd в разных структурах
Постсубикулуюм – есть реакция на место
Отличия hd в разных структурах
Постсубикулуюм – есть реакция на место

Слайд 106When the shape of the animal's environment is
changed, for example, from a
When the shape of the animal's environment is
changed, for example, from a

Слайд 107the hippocampus acts as a cognitive mapping system, which we shall call
the hippocampus acts as a cognitive mapping system, which we shall call

Слайд 10875 cm
220 cm
Лебедев, Плескачева, Анохин, ЖВНД, 2012
Ø 35см
Ø 75см
Ø 150см
Ø 220см
Размер арены
75 cm
220 cm
Лебедев, Плескачева, Анохин, ЖВНД, 2012
Ø 35см
Ø 75см
Ø 150см
Ø 220см
Размер арены
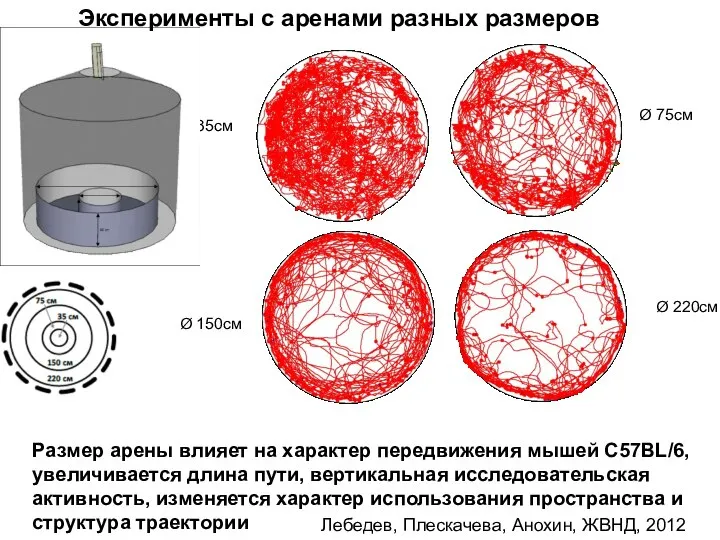
Слайд 109Размер арены влиял на интенсивность экспрессии с-Fos в каудальной области гиппокампа
Лебедев, 2012,
Размер арены влиял на интенсивность экспрессии с-Fos в каудальной области гиппокампа
Лебедев, 2012,
Слайд 1101st trial,
20 min
2nd trial,
20 min (ITI=24 h)
Small
Large
SS
SL
LL
LS
Apparatus: mice were tested in two
1st trial,
20 min
2nd trial,
20 min (ITI=24 h)
Small
Large
SS
SL
LL
LS
Apparatus: mice were tested in two
Слайд 111Частичное цитотоксическое билатеральное повреждение гиппокампа мышей с помощью введения NMDA было проведено
Частичное цитотоксическое билатеральное повреждение гиппокампа мышей с помощью введения NMDA было проведено
Слайд 112Частичное повреждение гиппокампа повлияло на интенсивность вертикальной активности при слабом воздействии на
Частичное повреждение гиппокампа повлияло на интенсивность вертикальной активности при слабом воздействии на
Слайд 113В большой арене частичное повреждение гиппокампа вызвало нарушение динамики угашения горизонтальной активности
1st
В большой арене частичное повреждение гиппокампа вызвало нарушение динамики угашения горизонтальной активности
1st
Слайд 114Повреждение гиппокампа изменило характеристики передвижения, особенно в большой арене
Мыши с поврежденным гиппокампом
Повреждение гиппокампа изменило характеристики передвижения, особенно в большой арене
Мыши с поврежденным гиппокампом
Слайд 115Удаление гиппокампа увеличило число скоростных протяженных сегментов у животных, передвигающихся в большой
Удаление гиппокампа увеличило число скоростных протяженных сегментов у животных, передвигающихся в большой
Слайд 116Mice
Voles
Samples of animal tracks in large arena (diam. 220 cm), Segment Analyzer
Mice
Voles
Samples of animal tracks in large arena (diam. 220 cm), Segment Analyzer
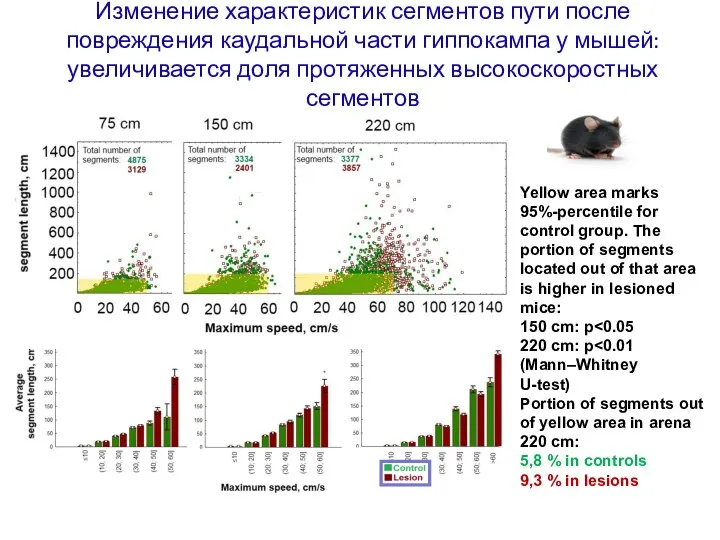
Слайд 117Изменение характеристик сегментов пути после повреждения каудальной части гиппокампа у мышей: увеличивается
Изменение характеристик сегментов пути после повреждения каудальной части гиппокампа у мышей: увеличивается
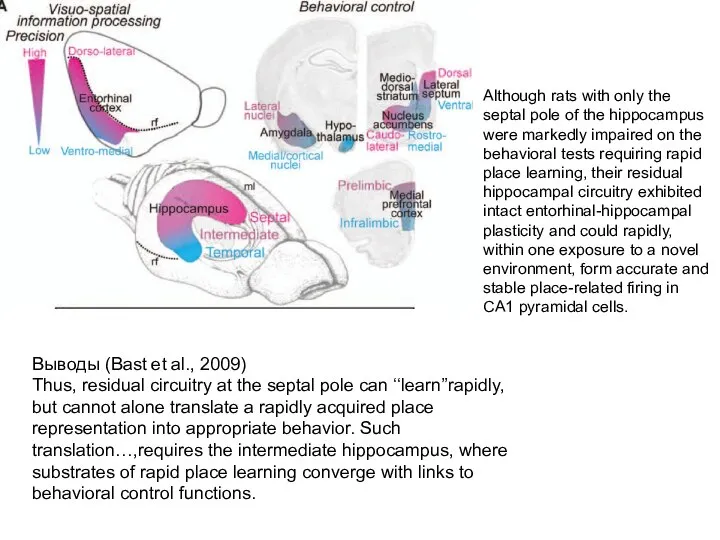
Слайд 118Выводы (Bast et al., 2009)
Thus, residual circuitry at the septal pole can
Выводы (Bast et al., 2009)
Thus, residual circuitry at the septal pole can
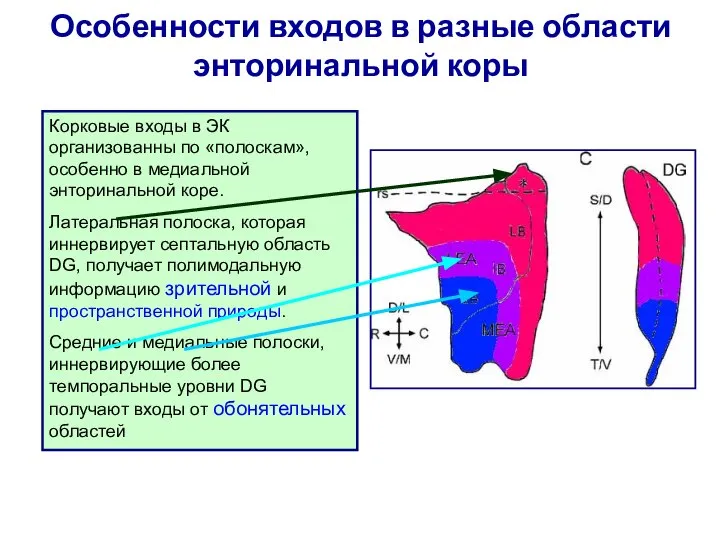
Слайд 119Особенности входов в разные области энторинальной коры
Корковые входы в ЭК организованны по
Особенности входов в разные области энторинальной коры
Корковые входы в ЭК организованны по
 Презентация на тему БАБОЧКИ
Презентация на тему БАБОЧКИ  Презентация на тему Одноклеточные и многоклеточные организмы
Презентация на тему Одноклеточные и многоклеточные организмы  Интересные факты о белках
Интересные факты о белках Черви. Коловратки
Черви. Коловратки Презентация на тему Царства живой природы
Презентация на тему Царства живой природы  Размножение. Бесполое размножение
Размножение. Бесполое размножение Верхние челюстные кости
Верхние челюстные кости Роль растений в жизни человека
Роль растений в жизни человека Опорно-двигательная система человека
Опорно-двигательная система человека Широколиственные леса
Широколиственные леса Презентация на тему Тканини рослин
Презентация на тему Тканини рослин  Медицинская энтомология
Медицинская энтомология Презентация на тему Сульфатредуцирующие бактерии
Презентация на тему Сульфатредуцирующие бактерии  Мутации. Мутанты
Мутации. Мутанты Презентация на тему Кровь
Презентация на тему Кровь  Болезни лошадей
Болезни лошадей Высев семян Ленкоранской акации
Высев семян Ленкоранской акации Внешнее и внутреннее строение стебля растения
Внешнее и внутреннее строение стебля растения Расселение бактерий. Биология. 6 класс
Расселение бактерий. Биология. 6 класс Строение инфузории и амебы
Строение инфузории и амебы 20141013_organicheskiy_mir_okeana
20141013_organicheskiy_mir_okeana Рабочая тетрадь. Биология 9 класс
Рабочая тетрадь. Биология 9 класс Эпителиальные ткани: особенности, функции, классификация
Эпителиальные ткани: особенности, функции, классификация Размножение и индивидуальное развитие организмов
Размножение и индивидуальное развитие организмов Лекарственные растения семейства буранчиковые
Лекарственные растения семейства буранчиковые Презентация на тему ЖАБЫ
Презентация на тему ЖАБЫ  Клетки ядерных организмов
Клетки ядерных организмов Приёмы рефлексии
Приёмы рефлексии