- Коэволюция

Содержание
- 2. КОЭВОЛЮЦИЯ 1. Эволюционные взаимоотношения видов, не связанных генетически (Иорданский) 2. Взаимные эволюционные изменения взаимодействующих видов (Thompson)
- 3. Коэволюция между близкими видами приводит к диверсификации Трёхиглые колюшки, живущие в одном и том же озере,
- 4. Антагонизм Мутуализм Хотя бы один вид вредит другому (когда они обитают вместе, то «пострадавший» хуже размножается)
- 5. Lithophragma parviflorum и моль Greya politella Географическая мозаика коэволюции, или от Антагонизма к мутуализму
- 6. Коэволюционная гонка вооружений В основе лежит частотно-зависимый отбор. Отбор благоприятствует паразитам, способным атаковать наиболее распространенный генотип
- 7. Брюхоногий моллюск Potamopyrgos antipodarum и сосальщик Microphallus sp. Пятилетние наблюдения подтвердили существование «флуктуирующего полиморфизма»
- 8. В эксперименте показано, что наиболее заразны фаги из будущего, а наиболее чувствительны к современным фагам бактерии
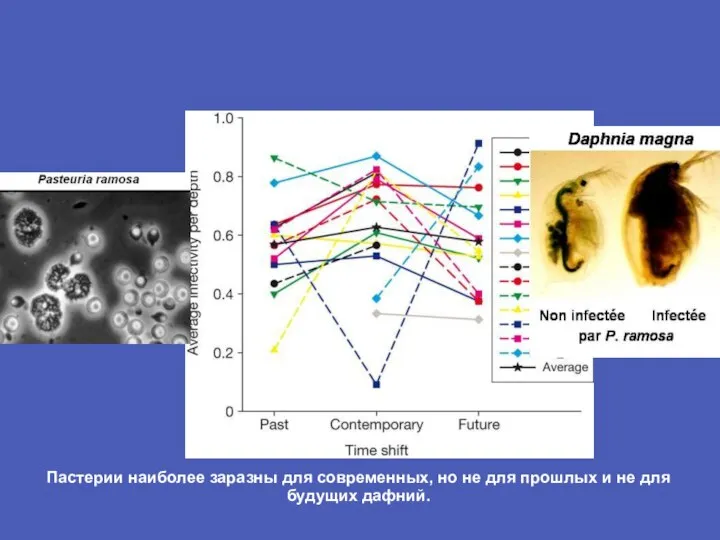
- 9. Пастерии наиболее заразны для современных, но не для прошлых и не для будущих дафний.
- 10. Обыкновенная кукушка Луговой конёк Тростниковая камышёвка Британская трясогузка Лесная завирушка В Исландии принимают яйца кукушек (не
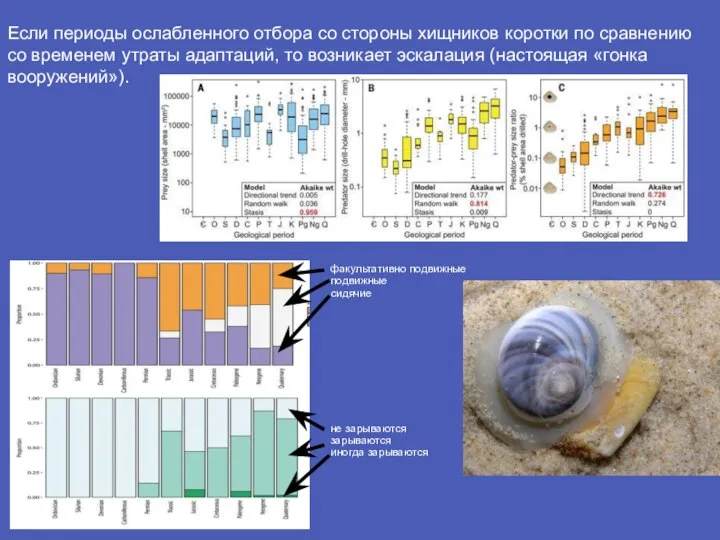
- 11. Если периоды ослабленного отбора со стороны хищников коротки по сравнению со временем утраты адаптаций, то возникает
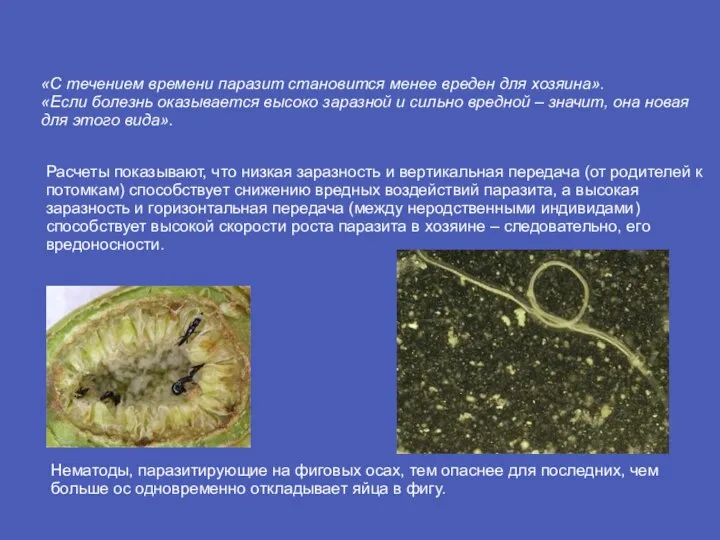
- 12. «С течением времени паразит становится менее вреден для хозяина». «Если болезнь оказывается высоко заразной и сильно
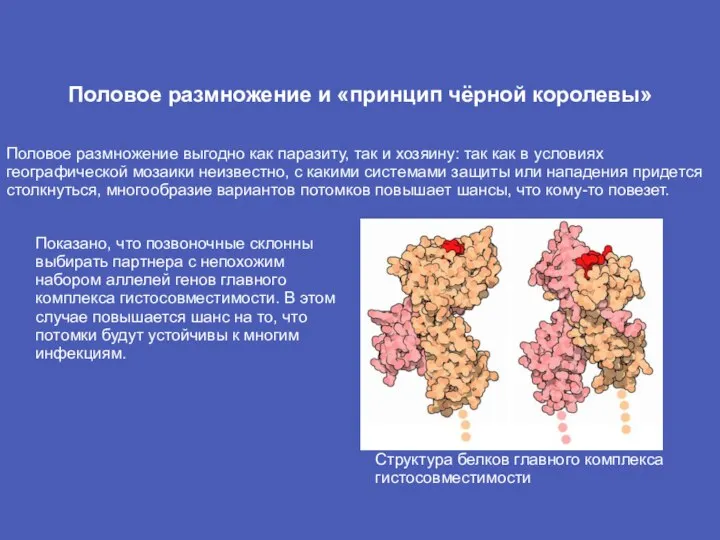
- 13. Половое размножение и «принцип чёрной королевы» Половое размножение выгодно как паразиту, так и хозяину: так как
- 14. Принцип Черной королевы (Red Queen principle) На самом деле, их два: макро- и микроэволюционный. Но в
- 15. светлые точки – клетки вольбахии вольбахии внутри клетки насекомого Drosophila simulans W W R R W
- 16. Гипотеза коэволюции к монокультуре и комплементарным симбионтам Симбиотический мутуализм эволюционирует из подмножества симбиотических взаимоотношений, в которых
- 17. Возникновению и поддержанию мутуализма способствует низкое генетическое разнообразие симбионтов Самки клопов передают своих симбионтов потомству через
- 18. Escovopsis sp. Acromyrmex octospinosus Streptomyces на кутикуле муравья Как и любая монокультура, мутуалистические взаимодействия подвержены опасности
- 19. Со временем симбионт и хозяин становятся всё более и более взаимозависимыми Митохондрии (органеллы, отвечающие за обеспечение
- 20. Мутуализм свободно живущих видов Основные модельные системы: растения и опылители, растения и плодоядные животные
- 21. Узкая взаимная специализация встречается среди свободно живущих мутуалистов, но является скорее исключением.
- 22. Сети свободно живущих мутуалистов
- 23. Сети свободно живущих мутуалистов Реальные мутуалистические сети обладают некими инвариантными свойствами: вложенностью, асимметрией взаимодействий и модульностью.
- 25. Скачать презентацию
Слайд 2КОЭВОЛЮЦИЯ
1. Эволюционные взаимоотношения видов, не связанных генетически (Иорданский)
2. Взаимные эволюционные изменения взаимодействующих
КОЭВОЛЮЦИЯ
1. Эволюционные взаимоотношения видов, не связанных генетически (Иорданский)
2. Взаимные эволюционные изменения взаимодействующих

Слайд 3Коэволюция между близкими видами приводит к диверсификации
Трёхиглые колюшки, живущие в одном и
Коэволюция между близкими видами приводит к диверсификации
Трёхиглые колюшки, живущие в одном и

Слайд 4Антагонизм
Мутуализм
Хотя бы один вид вредит другому (когда они обитают вместе, то «пострадавший»
Антагонизм
Мутуализм
Хотя бы один вид вредит другому (когда они обитают вместе, то «пострадавший»

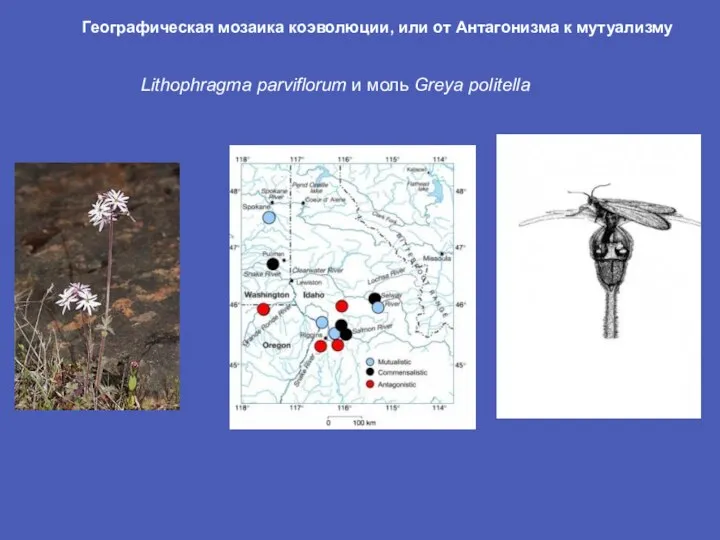
Слайд 5Lithophragma parviflorum и моль Greya politella
Географическая мозаика коэволюции, или от Антагонизма
Lithophragma parviflorum и моль Greya politella
Географическая мозаика коэволюции, или от Антагонизма
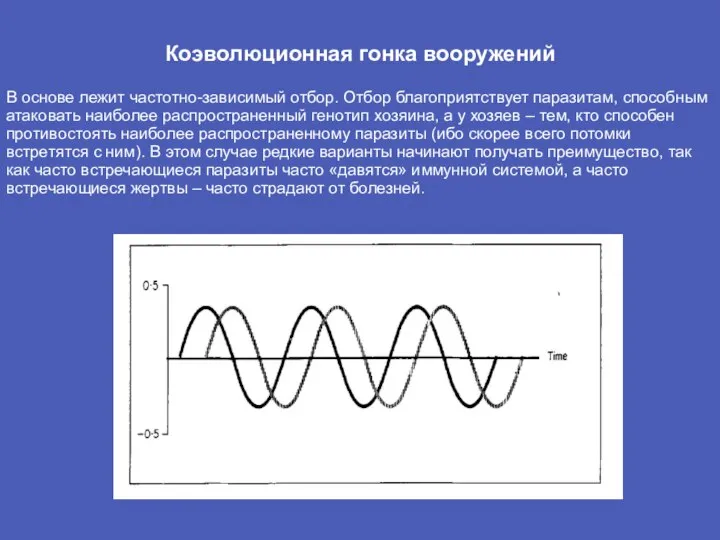
Слайд 6Коэволюционная гонка вооружений
В основе лежит частотно-зависимый отбор. Отбор благоприятствует паразитам, способным атаковать
Коэволюционная гонка вооружений
В основе лежит частотно-зависимый отбор. Отбор благоприятствует паразитам, способным атаковать
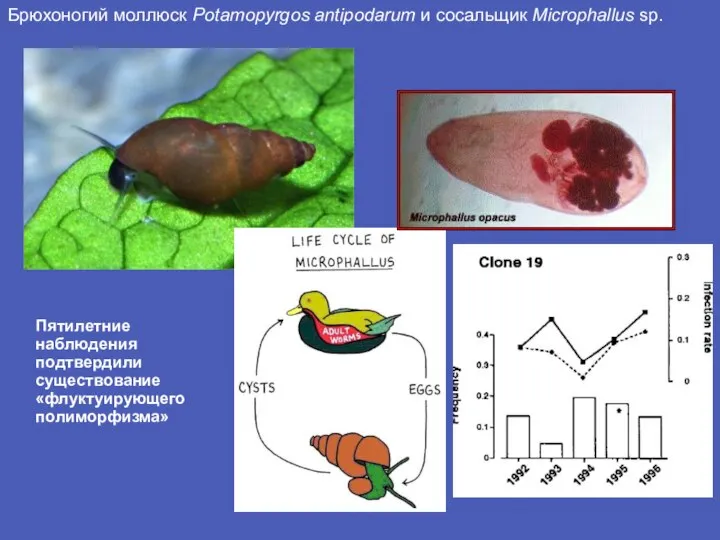
Слайд 7Брюхоногий моллюск Potamopyrgos antipodarum и сосальщик Microphallus sp.
Пятилетние наблюдения подтвердили существование «флуктуирующего
Брюхоногий моллюск Potamopyrgos antipodarum и сосальщик Microphallus sp.
Пятилетние наблюдения подтвердили существование «флуктуирующего
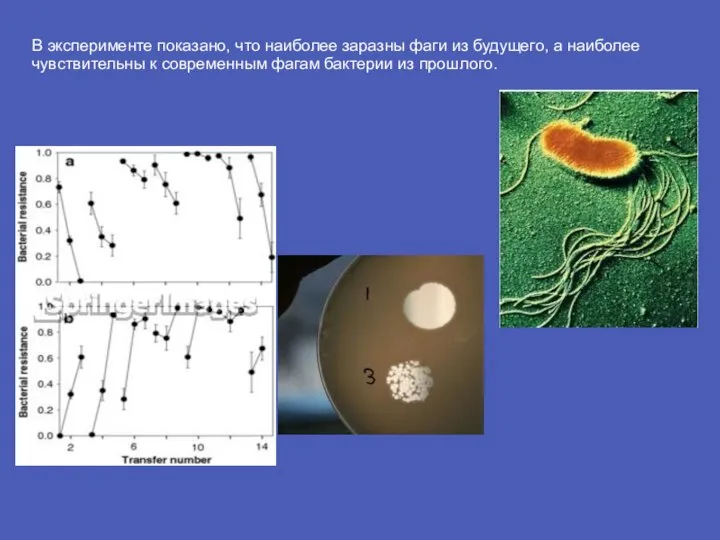
Слайд 8В эксперименте показано, что наиболее заразны фаги из будущего, а наиболее чувствительны
В эксперименте показано, что наиболее заразны фаги из будущего, а наиболее чувствительны
Слайд 9Пастерии наиболее заразны для современных, но не для прошлых и не для
Пастерии наиболее заразны для современных, но не для прошлых и не для
Слайд 10Обыкновенная кукушка
Луговой конёк
Тростниковая камышёвка
Британская трясогузка
Лесная завирушка
В Исландии
принимают яйца кукушек
(не умеет противодействовать кукушкам)
В
Обыкновенная кукушка
Луговой конёк
Тростниковая камышёвка
Британская трясогузка
Лесная завирушка
В Исландии
принимают яйца кукушек
(не умеет противодействовать кукушкам)
В

Слайд 11Если периоды ослабленного отбора со стороны хищников коротки по сравнению со временем
Если периоды ослабленного отбора со стороны хищников коротки по сравнению со временем
Слайд 12«С течением времени паразит становится менее вреден для хозяина».
«Если болезнь оказывается высоко
«С течением времени паразит становится менее вреден для хозяина». «Если болезнь оказывается высоко
Слайд 13Половое размножение и «принцип чёрной королевы»
Половое размножение выгодно как паразиту, так и
Половое размножение и «принцип чёрной королевы»
Половое размножение выгодно как паразиту, так и
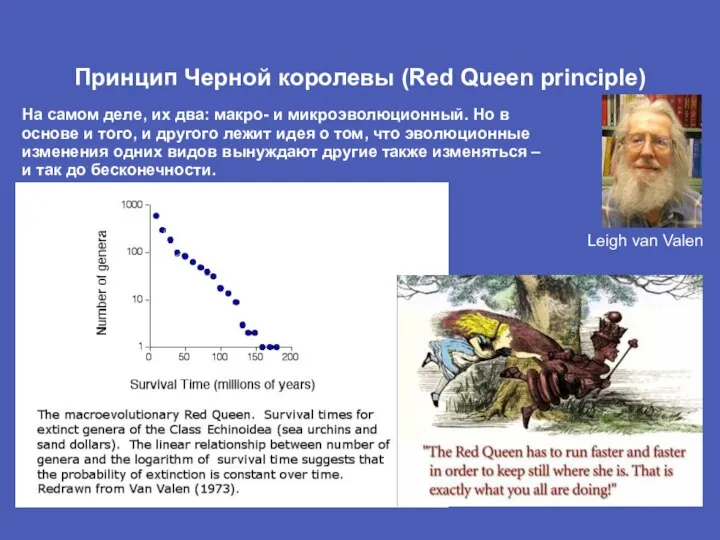
Слайд 14Принцип Черной королевы (Red Queen principle)
На самом деле, их два: макро- и
Принцип Черной королевы (Red Queen principle)
На самом деле, их два: макро- и
Слайд 15светлые точки – клетки вольбахии
вольбахии внутри клетки насекомого
Drosophila simulans
W
W
R
R
W – Watsonville (инфицированная
светлые точки – клетки вольбахии
вольбахии внутри клетки насекомого
Drosophila simulans
W
W
R
R
W – Watsonville (инфицированная

Слайд 16Гипотеза коэволюции к монокультуре и комплементарным симбионтам
Симбиотический мутуализм эволюционирует из подмножества симбиотических
Гипотеза коэволюции к монокультуре и комплементарным симбионтам
Симбиотический мутуализм эволюционирует из подмножества симбиотических

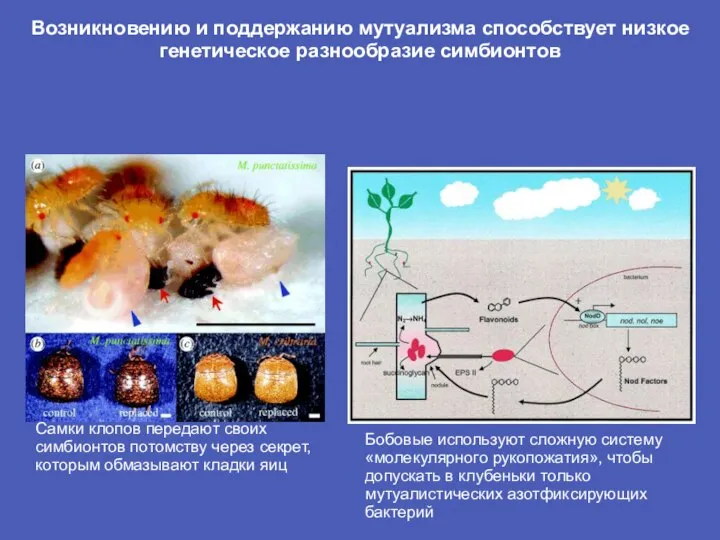
Слайд 17Возникновению и поддержанию мутуализма способствует низкое генетическое разнообразие симбионтов
Самки клопов передают своих
Возникновению и поддержанию мутуализма способствует низкое генетическое разнообразие симбионтов
Самки клопов передают своих
Слайд 18Escovopsis sp.
Acromyrmex octospinosus
Streptomyces на кутикуле муравья
Как и любая монокультура, мутуалистические взаимодействия
Escovopsis sp.
Acromyrmex octospinosus
Streptomyces на кутикуле муравья
Как и любая монокультура, мутуалистические взаимодействия

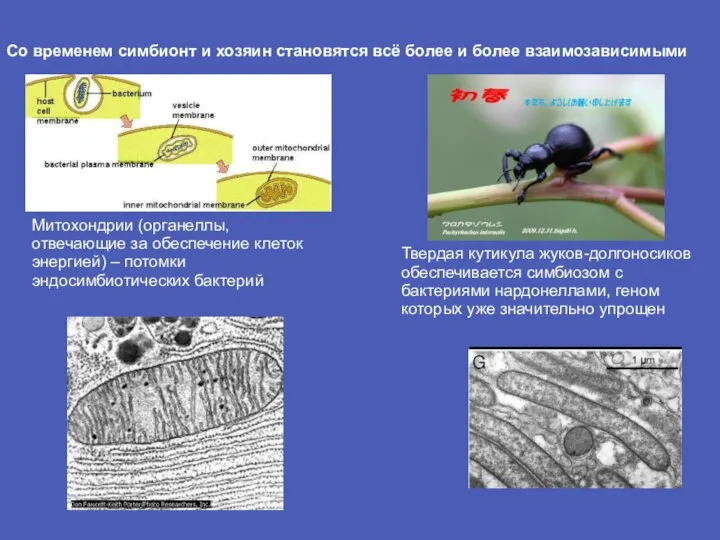
Слайд 19Со временем симбионт и хозяин становятся всё более и более взаимозависимыми
Митохондрии (органеллы,
Со временем симбионт и хозяин становятся всё более и более взаимозависимыми
Митохондрии (органеллы,
Слайд 20Мутуализм свободно живущих видов
Основные модельные системы: растения и опылители, растения и плодоядные
Мутуализм свободно живущих видов
Основные модельные системы: растения и опылители, растения и плодоядные

Слайд 21Узкая взаимная специализация встречается среди свободно живущих мутуалистов, но является скорее исключением.
Узкая взаимная специализация встречается среди свободно живущих мутуалистов, но является скорее исключением.

Слайд 22Сети свободно живущих мутуалистов
Сети свободно живущих мутуалистов
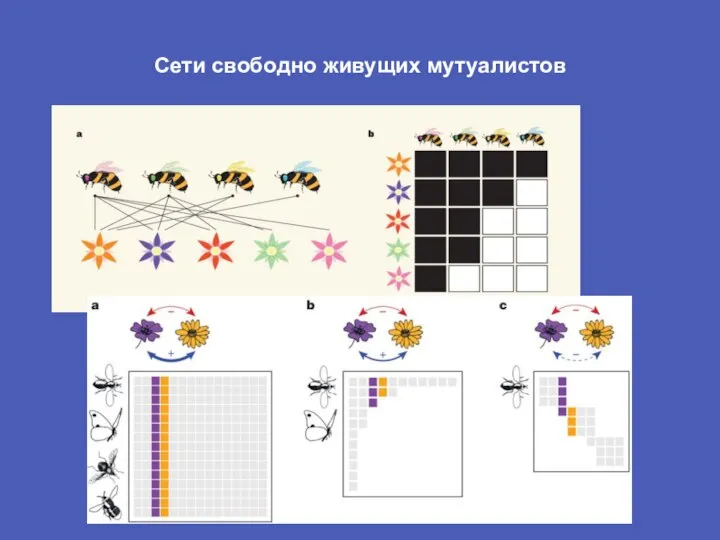
Слайд 23Сети свободно живущих мутуалистов
Реальные мутуалистические сети обладают некими инвариантными свойствами: вложенностью, асимметрией
Сети свободно живущих мутуалистов
Реальные мутуалистические сети обладают некими инвариантными свойствами: вложенностью, асимметрией

 Птахи, які виводять потомство взимку
Птахи, які виводять потомство взимку Старение. Теории о процессе старения
Старение. Теории о процессе старения Биохимия. Базовый уровень
Биохимия. Базовый уровень Нутрициология. Лекция 1
Нутрициология. Лекция 1 Транспортные системы организма. Круги кровообращения
Транспортные системы организма. Круги кровообращения Плод – орган семенного размножения
Плод – орган семенного размножения Типы отношений между организмами
Типы отношений между организмами Микробиология
Микробиология Компактизация хроматина
Компактизация хроматина Zheludochno-kishechny_trakt_Prezentatsia_1
Zheludochno-kishechny_trakt_Prezentatsia_1 Закономерности наследования признаков
Закономерности наследования признаков Семейство кувшинковые или нимфейные
Семейство кувшинковые или нимфейные Пищеварение в кишечнике. Всасывание. Роль печени. Аппендицит
Пищеварение в кишечнике. Всасывание. Роль печени. Аппендицит Таблицы по зоологии (часть 2)
Таблицы по зоологии (часть 2) Анатомо-физиологические особенности и методы исследования эндокринной системы, половое развитие ребёнка
Анатомо-физиологические особенности и методы исследования эндокринной системы, половое развитие ребёнка Эволюционное учение Ч. Дарвина
Эволюционное учение Ч. Дарвина Родительский лекторий. Качество питания и здоровье
Родительский лекторий. Качество питания и здоровье Тип Членистоногие
Тип Членистоногие Спинной мозг. Продолговатый мозг, мост, мозжечок. IV желудочек. Ромбовидная ямка
Спинной мозг. Продолговатый мозг, мост, мозжечок. IV желудочек. Ромбовидная ямка Египетская гладкошерстная кошка
Египетская гладкошерстная кошка Анатомия кишечника
Анатомия кишечника Берёза. Что такое берёза
Берёза. Что такое берёза Основы рационального питания
Основы рационального питания Презентация по биологии_ _Движение живых организмов_
Презентация по биологии_ _Движение живых организмов_ Ядовитые грибы и ягоды
Ядовитые грибы и ягоды Типы вегетативного размножения комнатных растений
Типы вегетативного размножения комнатных растений Дыхательные движения
Дыхательные движения Презентация на тему Орган зрения
Презентация на тему Орган зрения