- Конъюгация. F-плазмида

Содержание
- 2. F-плазмида F-плазмида кодирует все белки, необходимые для конъюгации. Также имеются: ориджин для тета-репликации, второй ориджин (слабый)
- 3. Гены tra F-плазмиды Dtr-гены, или Dtr-компоненты (DNA Transfer and Replication). Необходимы для процессинга плазмиды при подготовке
- 4. Аппарат переноса F-плазмиды из клетки в клетку Основной компонент аппарата – пилус, структура, скрепляющая друг с
- 5. Сцепляющие белки (coupling proteins) Белки – компоненты Mpf-аппарата, осуществляющие его связь с Dtr-аппаратом и сигнализирующие ему
- 6. Dtr-аппарат Состоит из нескольких белков, осуществляющих разные функции. Релаксаза (TraI в случае F-плазмиды) является частью большого
- 7. Работа Dtr-аппарата Клетка-донор и клетка-реципиент находятся неподалеку друг от друга. Собирается Mpf-структура, образуется пара «брачующихся» клеток.
- 8. Работа Dtr-аппарата Релаксаза и хеликаза в комплексе с одной из цепочек плазмиды проникают в клетку-реципиент. Релаксаза
- 9. Регуляция конъюгации Процесс конъюгации обычно эффективен только в первое время после того, как плазмида попала в
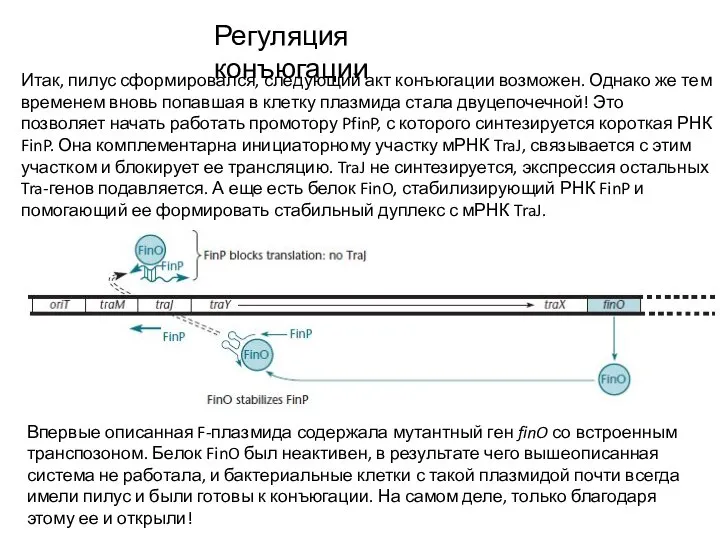
- 10. Регуляция конъюгации Итак, пилус сформировался, следующий акт конъюгации возможен. Однако же тем временем вновь попавшая в
- 11. Бактериальный промискуитет Перенос плазмиды из бактерию в бактерию другого вида – рядовое событие. Существует класс плазмид,
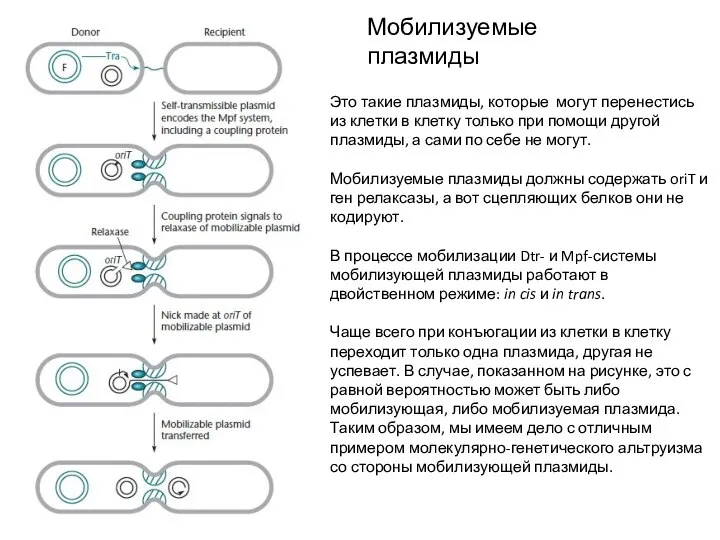
- 12. Мобилизуемые плазмиды Это такие плазмиды, которые могут перенестись из клетки в клетку только при помощи другой
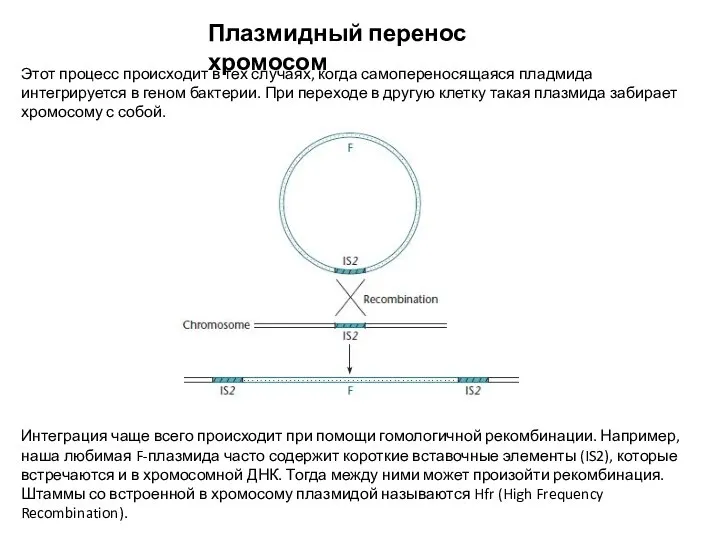
- 13. Плазмидный перенос хромосом Этот процесс происходит в тех случаях, когда самопереносящаяся пладмида интегрируется в геном бактерии.
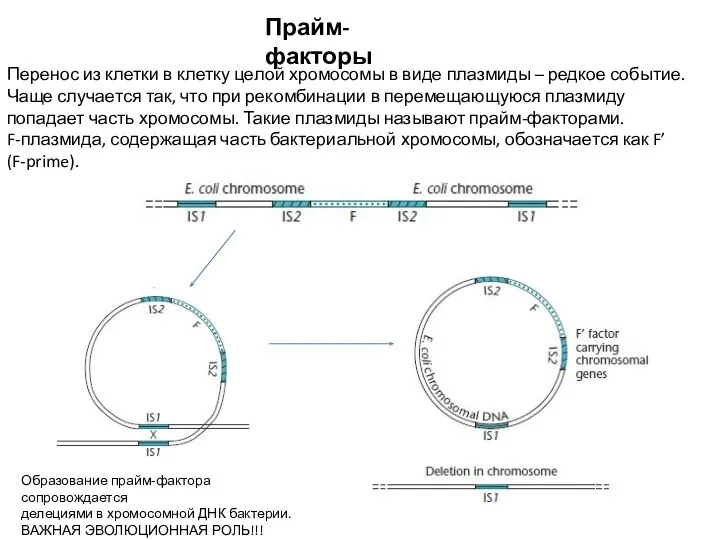
- 14. Прайм-факторы Перенос из клетки в клетку целой хромосомы в виде плазмиды – редкое событие. Чаще случается
- 15. Конъюгация у грам-положительных бактерий В целом организована так же, как и у грам-отрицательных, однако имеются интересные
- 16. Конъюгация у грам-положительных бактерий Клетка-донор содержит плазмиду pAD1, с которой синтезируется белок TraA – транскрипционный репрессор,
- 17. Конъюгация у грам-положительных бактерий Феромон cAD1 связывается с TraC и проникает внутрь клетки-донора через специальную транспортную
- 18. Конъюгация у грам-положительных бактерий Собственно конъюгация происходит практически так же, как и у грам-отрицательных бактерий. Плазмида
- 19. Конъюгация у грам-положительных бактерий Выключение конъюгации у трансконъюгантов происходит по нескольким механизмам. Олигопептид iAD1, закодированный в
- 20. Конъюгация у грам-положительных бактерий: интегративные конъюгирующие элементы (ICE) ICE не способны к автономной репликации, то есть
- 21. Самовырезание Tn916. Белки Int и Xis вносят разрывы в хромосому неподалеку от границ ICE. Образуются липкие
- 22. Самовстраивание Tn916. Перед конъюгацией кольцевой ICE никируется, и одна цепочка ICE проникает в реципиента. Там на
- 23. Трансформация Трансформацией называется процесс поглощения бактериальной клеткой ДНК из внешней среды. Способность поглощать ДНК из внешней
- 24. Природная компетентность грам-положительных бактерий (B. subtilis) Com-аппарат состоит из белков ComG (формирует пилус), ComEA (рецептор для
- 25. Природная компетентность грам-отрицательных бактерий (N. gonorrhoeae) Все примерно то же самое, только для транспорта через внешнюю
- 26. Регуляция компетентности у B. subtilis Компетентность у этой бактерии проявляется в момент истощения ресурсов. Истощение сопровождается
- 28. Скачать презентацию
Слайд 2F-плазмида
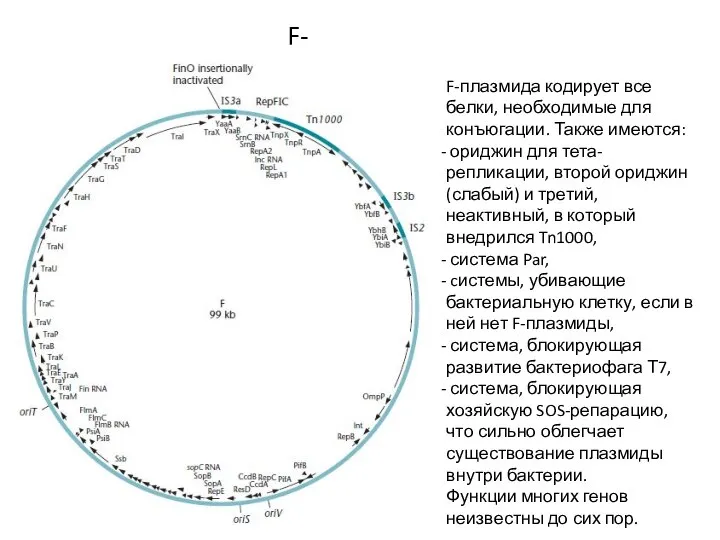
F-плазмида кодирует все белки, необходимые для конъюгации. Также имеются:
ориджин для тета-репликации,
F-плазмида
F-плазмида кодирует все белки, необходимые для конъюгации. Также имеются:
ориджин для тета-репликации,
Слайд 3Гены tra F-плазмиды
Dtr-гены, или Dtr-компоненты
(DNA Transfer and Replication).
Необходимы для процессинга плазмиды
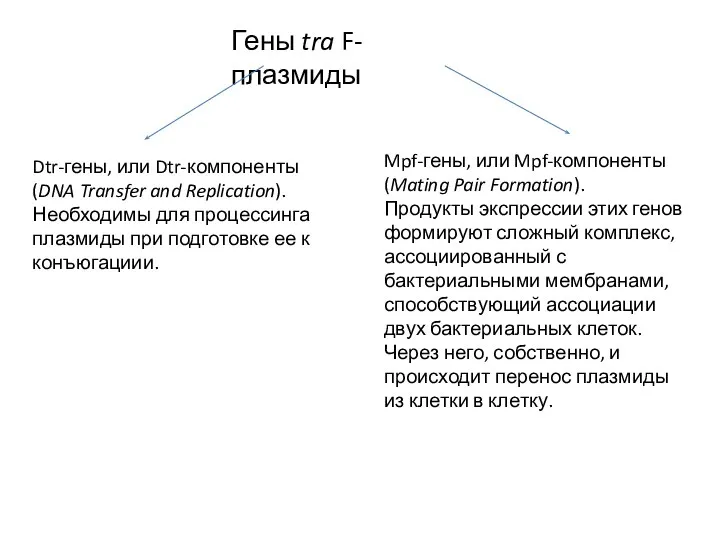
Гены tra F-плазмиды
Dtr-гены, или Dtr-компоненты
(DNA Transfer and Replication).
Необходимы для процессинга плазмиды
Слайд 4Аппарат переноса F-плазмиды из клетки в клетку
Основной компонент аппарата – пилус, структура,
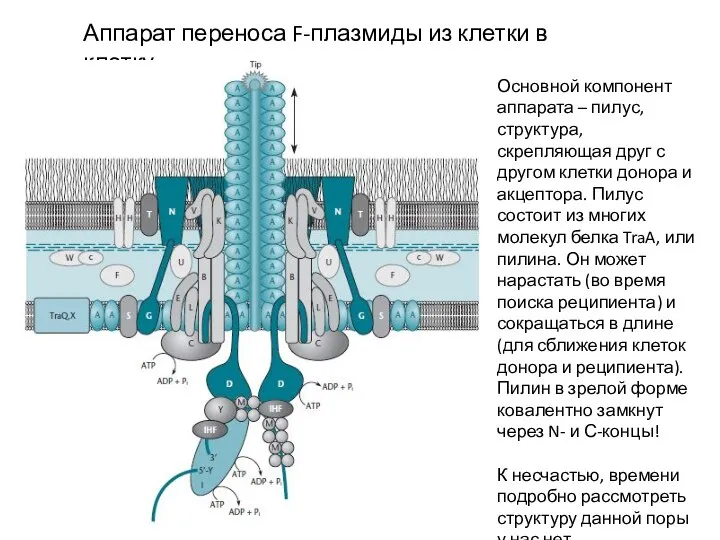
Аппарат переноса F-плазмиды из клетки в клетку
Основной компонент аппарата – пилус, структура,
Слайд 5Сцепляющие белки (coupling proteins)
Белки – компоненты Mpf-аппарата, осуществляющие его связь с Dtr-аппаратом
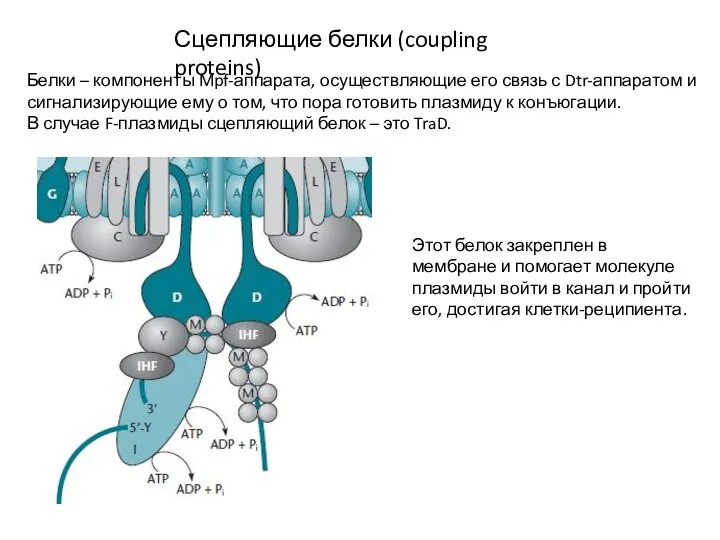
Сцепляющие белки (coupling proteins)
Белки – компоненты Mpf-аппарата, осуществляющие его связь с Dtr-аппаратом
Слайд 6Dtr-аппарат
Состоит из нескольких белков, осуществляющих разные функции.
Релаксаза (TraI в случае F-плазмиды) является
Dtr-аппарат
Состоит из нескольких белков, осуществляющих разные функции.
Релаксаза (TraI в случае F-плазмиды) является
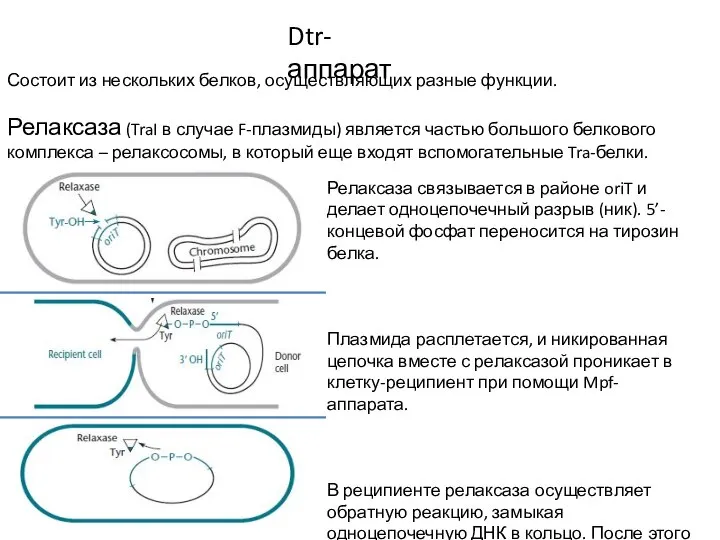
Слайд 7Работа Dtr-аппарата
Клетка-донор и клетка-реципиент находятся неподалеку друг от друга.
Собирается Mpf-структура, образуется пара
Работа Dtr-аппарата
Клетка-донор и клетка-реципиент находятся неподалеку друг от друга.
Собирается Mpf-структура, образуется пара
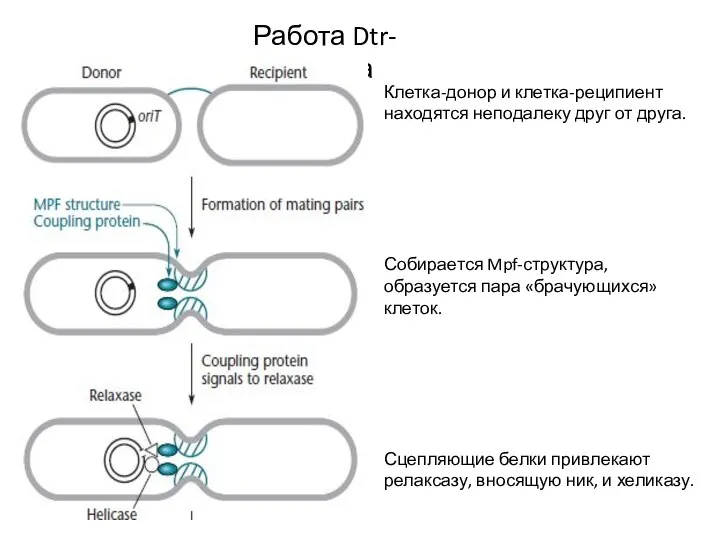
Слайд 8Работа Dtr-аппарата
Релаксаза и хеликаза в комплексе с одной из цепочек плазмиды проникают
Работа Dtr-аппарата
Релаксаза и хеликаза в комплексе с одной из цепочек плазмиды проникают
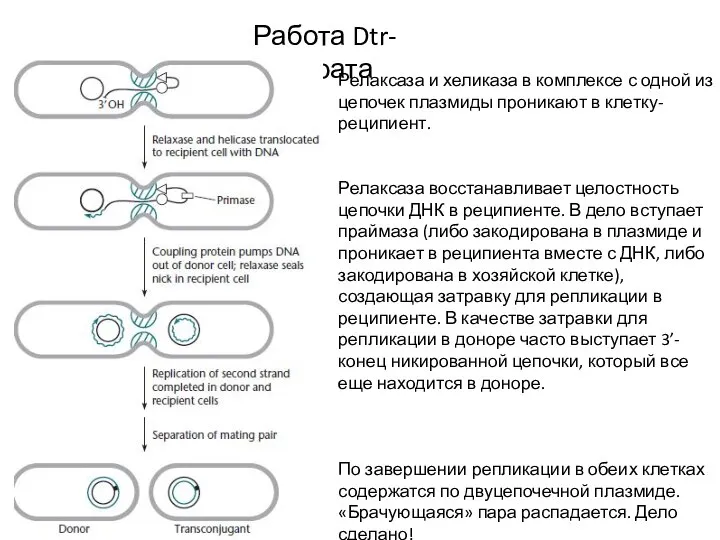
Слайд 9Регуляция конъюгации
Процесс конъюгации обычно эффективен только в первое время после того, как
Регуляция конъюгации
Процесс конъюгации обычно эффективен только в первое время после того, как
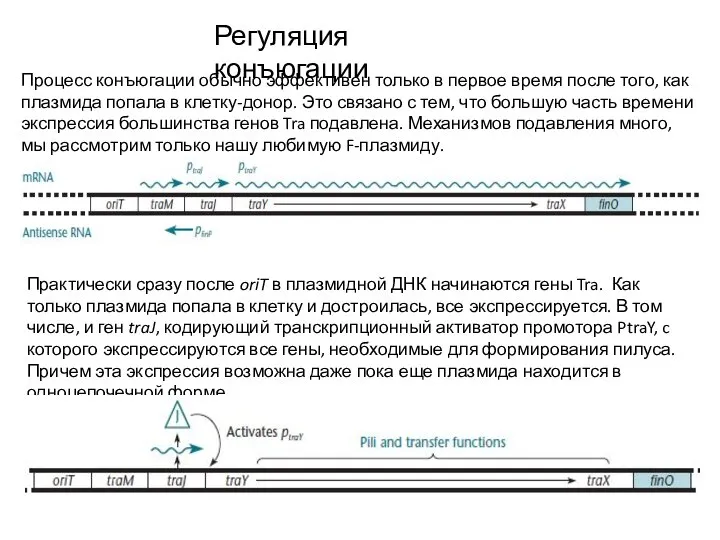
Слайд 10Регуляция конъюгации
Итак, пилус сформировался, следующий акт конъюгации возможен. Однако же тем временем
Регуляция конъюгации
Итак, пилус сформировался, следующий акт конъюгации возможен. Однако же тем временем
Слайд 11Бактериальный промискуитет
Перенос плазмиды из бактерию в бактерию другого вида – рядовое событие.
Существует
Бактериальный промискуитет
Перенос плазмиды из бактерию в бактерию другого вида – рядовое событие.
Существует

Слайд 12Мобилизуемые плазмиды
Это такие плазмиды, которые могут перенестись из клетки в клетку только
Мобилизуемые плазмиды
Это такие плазмиды, которые могут перенестись из клетки в клетку только
Слайд 13Плазмидный перенос хромосом
Этот процесс происходит в тех случаях, когда самопереносящаяся пладмида интегрируется
Плазмидный перенос хромосом
Этот процесс происходит в тех случаях, когда самопереносящаяся пладмида интегрируется
Слайд 14Прайм-факторы
Перенос из клетки в клетку целой хромосомы в виде плазмиды – редкое
Прайм-факторы
Перенос из клетки в клетку целой хромосомы в виде плазмиды – редкое
Слайд 15Конъюгация у грам-положительных бактерий
В целом организована так же, как и у грам-отрицательных,
Конъюгация у грам-положительных бактерий
В целом организована так же, как и у грам-отрицательных,
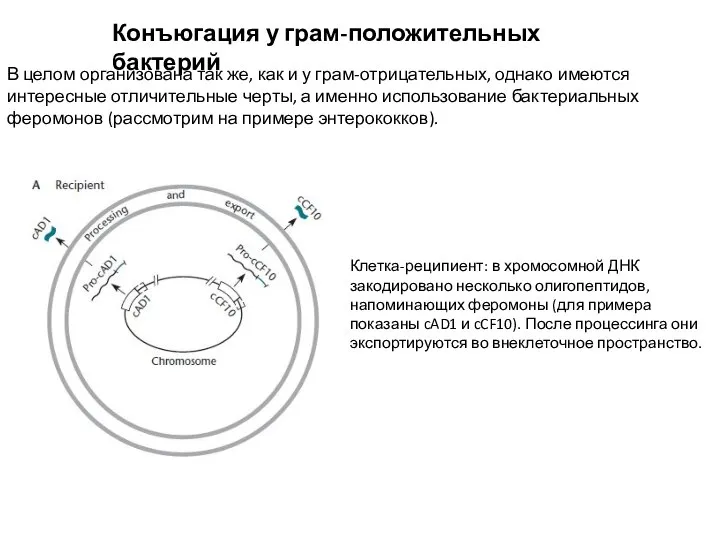
Слайд 16Конъюгация у грам-положительных бактерий
Клетка-донор содержит плазмиду pAD1, с которой синтезируется белок TraA
Конъюгация у грам-положительных бактерий
Клетка-донор содержит плазмиду pAD1, с которой синтезируется белок TraA
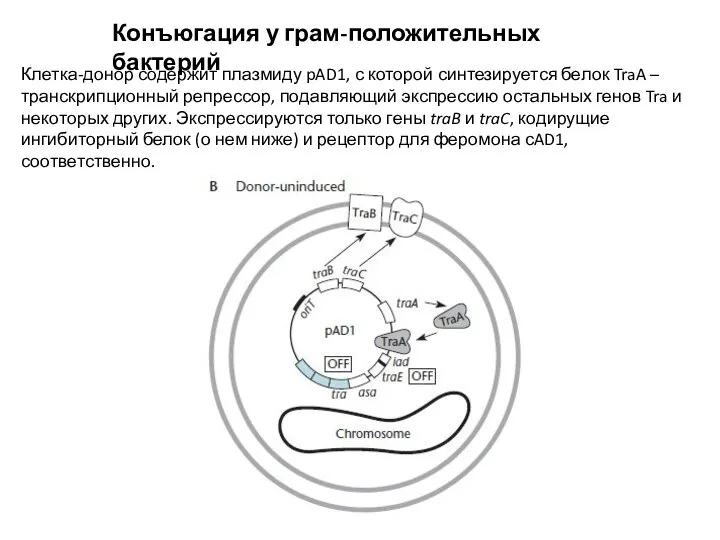
Слайд 17Конъюгация у грам-положительных бактерий
Феромон cAD1 связывается с TraC и проникает внутрь клетки-донора
Конъюгация у грам-положительных бактерий
Феромон cAD1 связывается с TraC и проникает внутрь клетки-донора
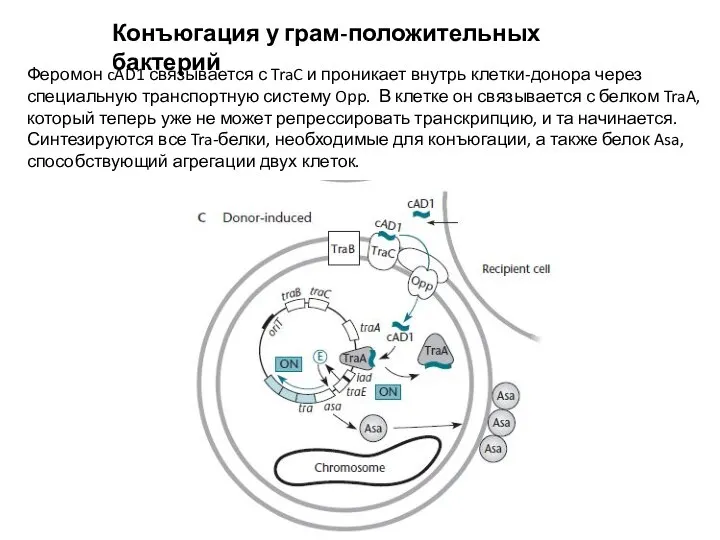
Слайд 18Конъюгация у грам-положительных бактерий
Собственно конъюгация происходит практически так же, как и у
Конъюгация у грам-положительных бактерий
Собственно конъюгация происходит практически так же, как и у
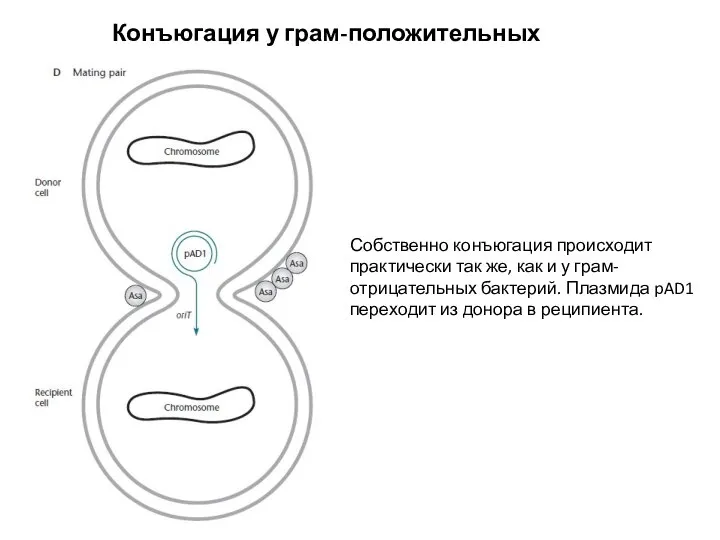
Слайд 19Конъюгация у грам-положительных бактерий
Выключение конъюгации у трансконъюгантов происходит по нескольким механизмам.
Олигопептид iAD1,
Конъюгация у грам-положительных бактерий
Выключение конъюгации у трансконъюгантов происходит по нескольким механизмам.
Олигопептид iAD1,
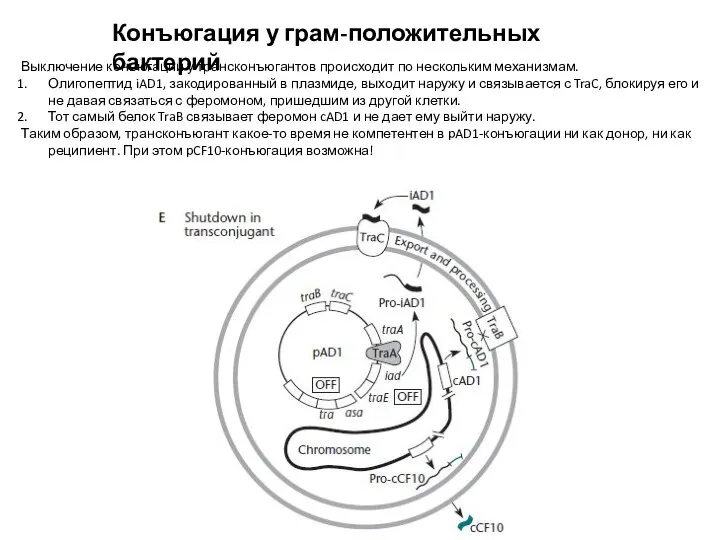
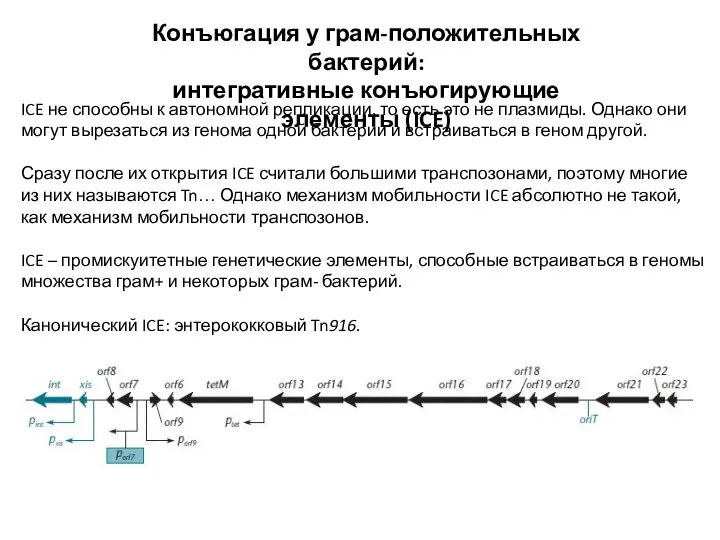
Слайд 20Конъюгация у грам-положительных бактерий:
интегративные конъюгирующие элементы (ICE)
ICE не способны к автономной репликации,
Конъюгация у грам-положительных бактерий:
интегративные конъюгирующие элементы (ICE)
ICE не способны к автономной репликации,
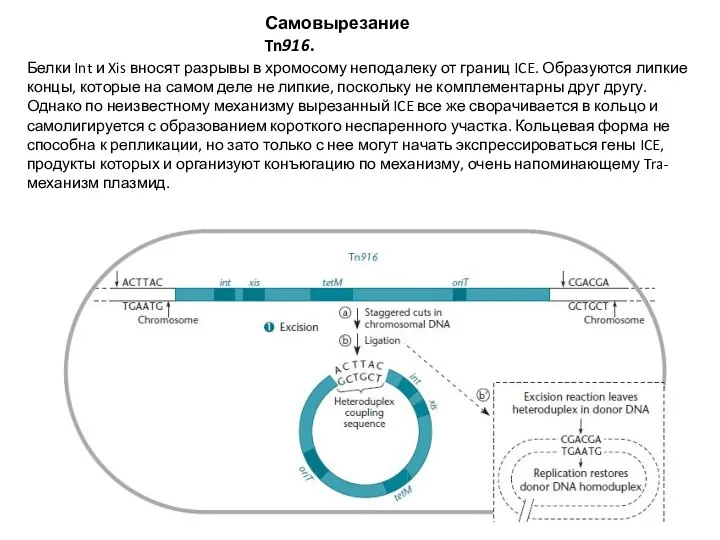
Слайд 21Самовырезание Tn916.
Белки Int и Xis вносят разрывы в хромосому неподалеку от границ
Самовырезание Tn916.
Белки Int и Xis вносят разрывы в хромосому неподалеку от границ
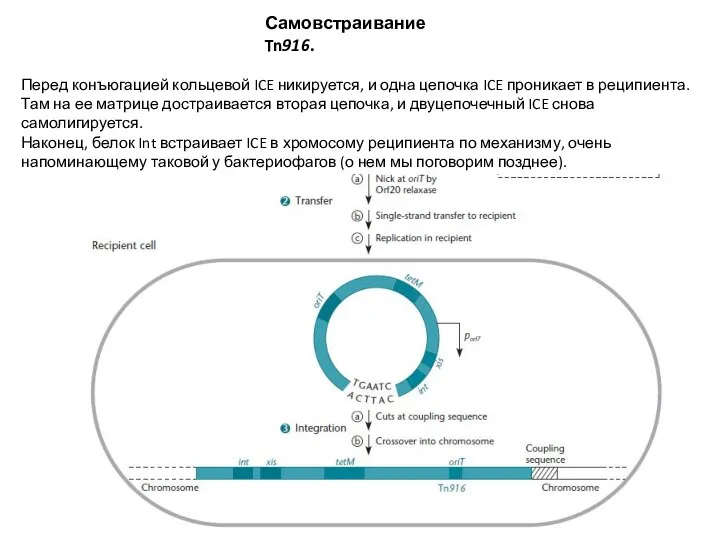
Слайд 22Самовстраивание Tn916.
Перед конъюгацией кольцевой ICE никируется, и одна цепочка ICE проникает в
Самовстраивание Tn916.
Перед конъюгацией кольцевой ICE никируется, и одна цепочка ICE проникает в
Слайд 23Трансформация
Трансформацией называется процесс поглощения бактериальной клеткой ДНК из внешней среды.
Способность поглощать ДНК
Трансформация
Трансформацией называется процесс поглощения бактериальной клеткой ДНК из внешней среды.
Способность поглощать ДНК
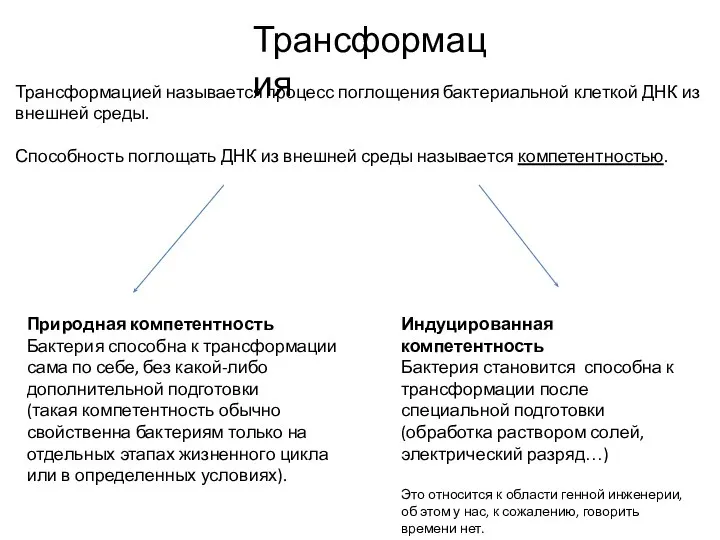
Слайд 24Природная компетентность грам-положительных бактерий (B. subtilis)
Com-аппарат состоит из белков ComG (формирует пилус),
Природная компетентность грам-положительных бактерий (B. subtilis)
Com-аппарат состоит из белков ComG (формирует пилус),
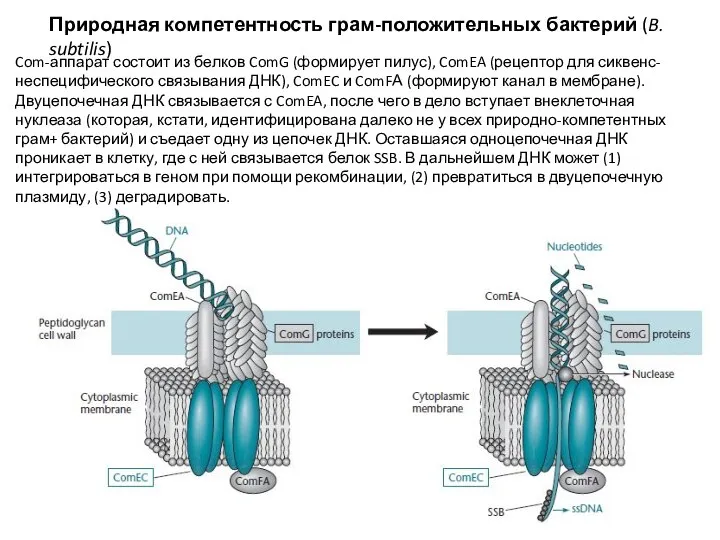
Слайд 25Природная компетентность грам-отрицательных бактерий (N. gonorrhoeae)
Все примерно то же самое, только для
Природная компетентность грам-отрицательных бактерий (N. gonorrhoeae)
Все примерно то же самое, только для
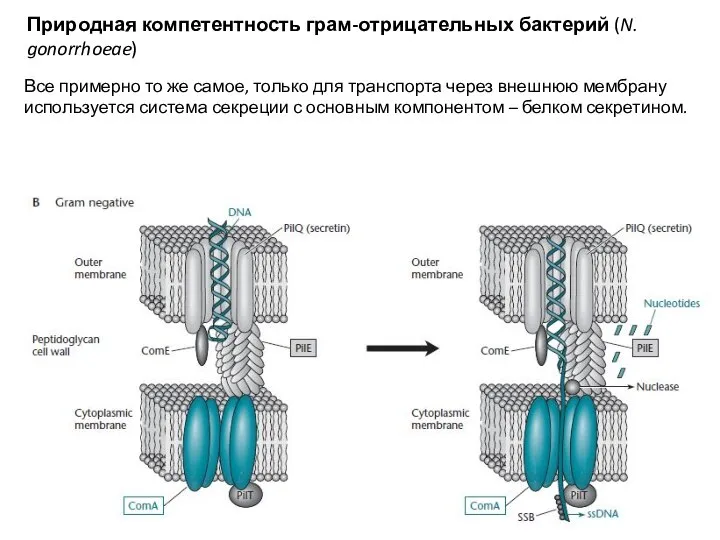
Слайд 26Регуляция компетентности у B. subtilis
Компетентность у этой бактерии проявляется в момент истощения
Регуляция компетентности у B. subtilis
Компетентность у этой бактерии проявляется в момент истощения
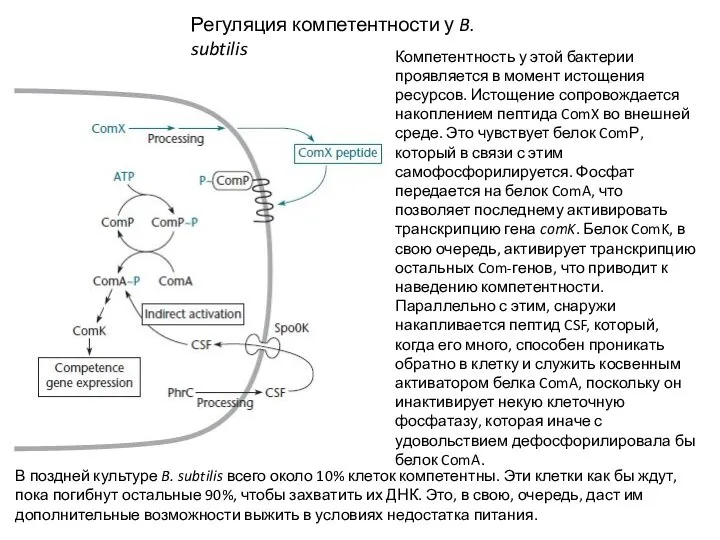
 خمسة أنواع من الماشية
خمسة أنواع من الماشية Физиология микробов
Физиология микробов Структура популяций
Структура популяций Общие принципы работы организма
Общие принципы работы организма Анатомия насекомых
Анатомия насекомых Кодирование звука
Кодирование звука Презентация на тему Геометрия и "живые" молекулы
Презентация на тему Геометрия и "живые" молекулы  Борьба за существование и естественный отбор – движущая сила эволюции
Борьба за существование и естественный отбор – движущая сила эволюции Птицы школьного двора
Птицы школьного двора Китообразные животные
Китообразные животные Хвостатые и бесхвостые земноводные
Хвостатые и бесхвостые земноводные Нуклеиновые кислоты. Системы репликации и репарации. Лекция 2
Нуклеиновые кислоты. Системы репликации и репарации. Лекция 2 Кожа
Кожа Строение клетки
Строение клетки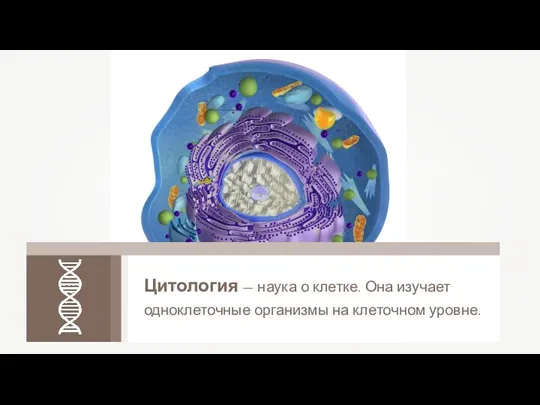 Цитология – наука о клетке
Цитология – наука о клетке Взаимодействие аллельных генов
Взаимодействие аллельных генов Опорно-двигательная система
Опорно-двигательная система zrenie
zrenie Муравьи - социальные животные
Муравьи - социальные животные Презентация на тему Витамины и их роль в жизни людей
Презентация на тему Витамины и их роль в жизни людей  История собаководства
История собаководства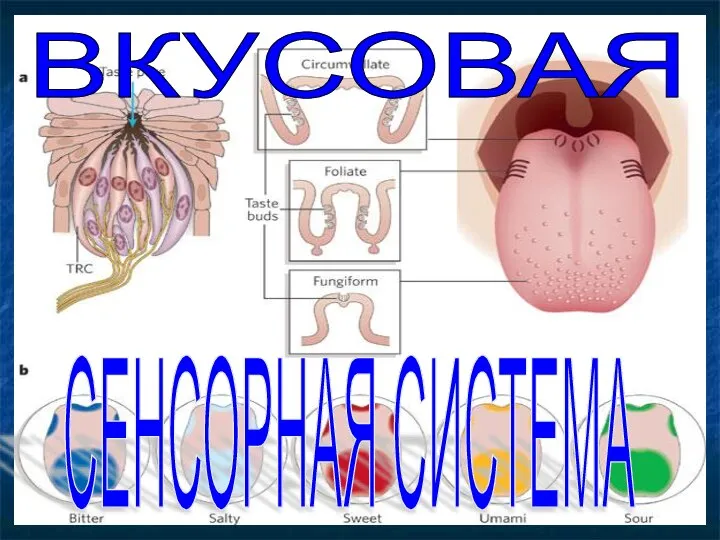 Вкусовая сенсорная система
Вкусовая сенсорная система Садовые цветы
Садовые цветы Обменные процессы в организме
Обменные процессы в организме Кроветворение
Кроветворение Животные. Разнообразие
Животные. Разнообразие Бабочки, многообразие
Бабочки, многообразие Презентация на тему Вспомогательные органы кровеносной системы
Презентация на тему Вспомогательные органы кровеносной системы