- Лекция 11. Пыльцевая трубка. Основные механизмы полярного роста

Содержание
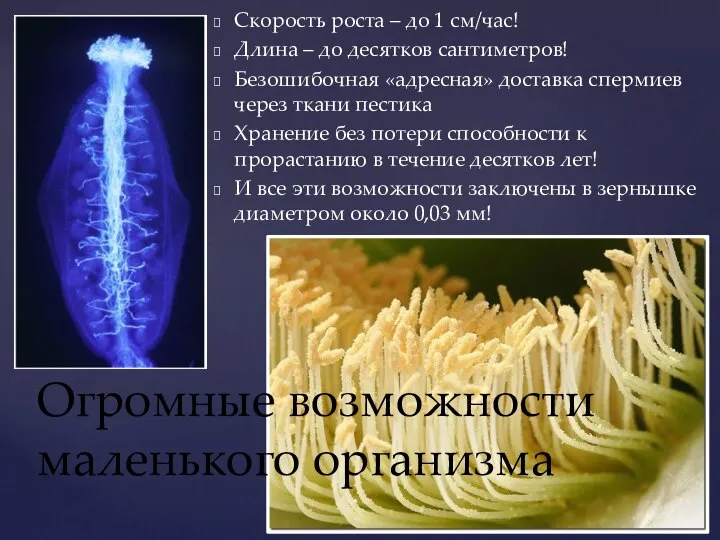
- 2. Скорость роста – до 1 см/час! Длина – до десятков сантиметров! Безошибочная «адресная» доставка спермиев через
- 3. 1824 1847 Ассирийские священники и египетские боги изображались в процессе ритуального опыления пальмы (5000 лет до
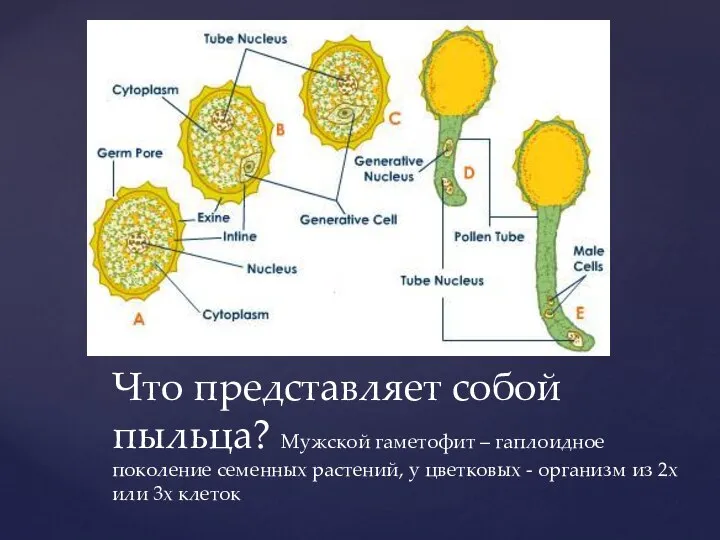
- 4. Что представляет собой пыльца? Мужской гаметофит – гаплоидное поколение семенных растений, у цветковых - организм из
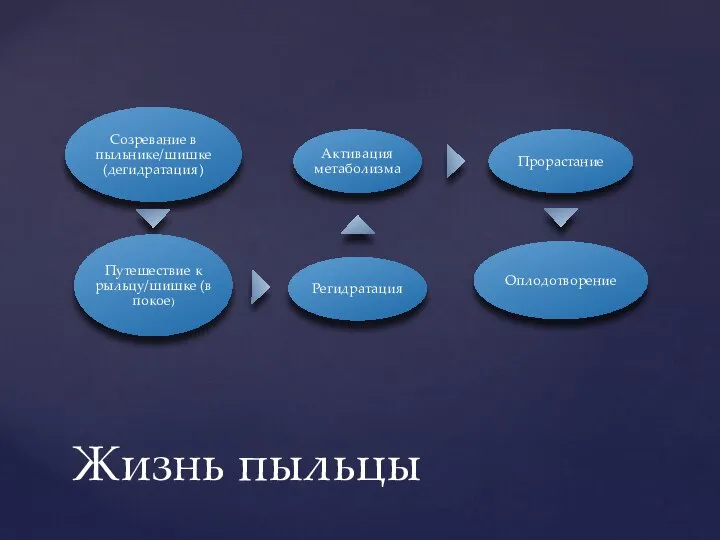
- 5. Жизнь пыльцы
- 6. Пыльцевая трубка как модельный объект Гаплоидный геном – легко выявить важные гены Отсутствие межклеточных контактов –
- 7. Ключевые механизмы поддержания полярного роста: Зональная организация цитоплазмы позволяет каждой органелле быть «в нужное время в
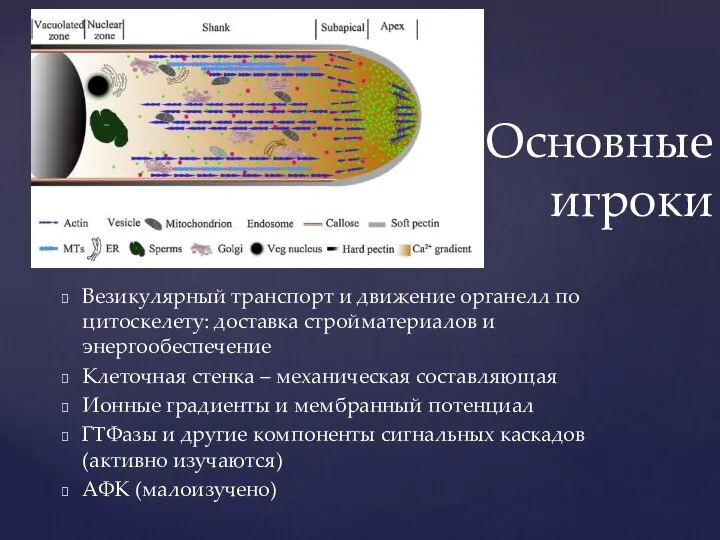
- 8. Везикулярный транспорт и движение органелл по цитоскелету: доставка стройматериалов и энергообеспечение Клеточная стенка – механическая составляющая
- 9. Апикальный (или «растущий») компартмент Базальный компартмент Апикальный рост пыльцевой трубки поддерживается за счет полярной организации цитоплазмы.
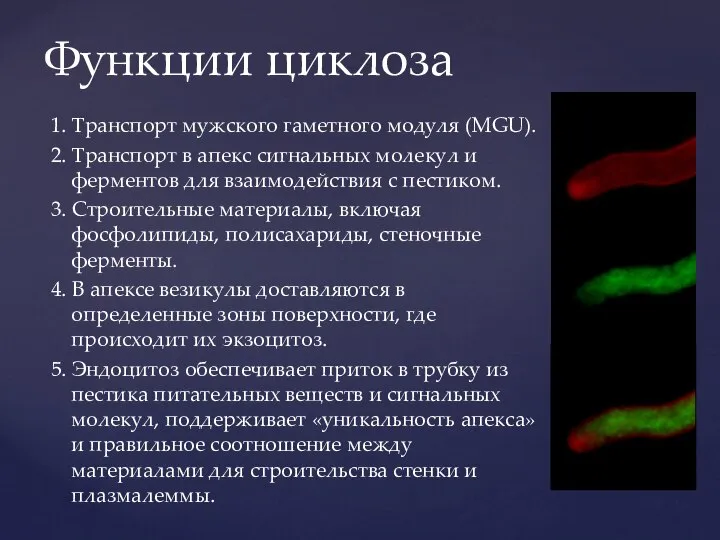
- 10. Функции циклоза 1. Транспорт мужского гаметного модуля (MGU). 2. Транспорт в апекс сигнальных молекул и ферментов
- 11. Растущий компартмент Lancelle, Hepler, 1992 Брейгина и др., 2009
- 12. Цитоскелет
- 13. Актиновый цитоскелет играет ключевую роль в поддержании полярного роста Продольные актиновые тяжи обеспечивают ток цитоплазмы (фонтан)
- 14. Микротрубочки Отвечает за транспорт спермиев к зародышевому мешку Остальные функции неизвестны Ингибиторы не останавливают рост ПТ
- 15. Рециклирование мембран необходимо для поддержания полярности – «уникальность апекса». В пыльцевой трубке образование окаймленных пузырьков с
- 16. Поляризованная секреция поддерживается благодаря актиновому цитоскелету, а также аннексину и белкам экзоцистного комплекса Экзоцисты предсталяют из
- 17. Клеточная стенка: градиент жесткости Движущей силой для роста является тургорное давление. Однако, оно не является векторной
- 18. Этерифицированные пектины откладываются в кончике трубки в процессе экзоцитоза По мере удаления от кончика они деэтерифицируются
- 19. Каллоза Breygina et al., 2012 Xie et al., 2012 Отсутствует в кончике Появляется на расстоянии 10-30
- 20. Жесткий компонент, но его не очень много Отложение также неравномерное: в кончике отсутствует, далее постепенно накапливается
- 21. Регуляторные механизмы: кальций Hepler, 2012 Кальций входит в кончике трубки через каналы , а в субапикальной
- 22. Кальциевый градиент определяет место слияния везикул с ПМ и, таким образом, задает направление роста Кальций регулирует
- 23. В кончике рН кислый, протоны входят в цитоплазму, предположительно, через неспецифичные катионные каналы. Кислый кончик присутствует
- 24. контроль ортованадат фузикокцин Брейгина и др., 2010 Градиент мембранного потенциала контроль ортованадат NPPB Брейгина и др.,
- 25. Таким образом, пыльцевая трубка – прекрасная модель для изучения ИОННОЙ РЕГУЛЯЦИИ РОСТА, которая включает в себя
- 26. Регуляторные механизмы: ГТФазы Qin, Yang, 2011 GFP-NtRab2 Маленькие ГТФ-связывающие белки - универсальные переключатели сигнальных путей. Rab2
- 27. Фосфатидилинозитолдифосфат (PIP2) есть только в апикальной мембране растущих ПТ Его узкая локализация обеспечивается белком phosphotidylinositol transfer
- 28. Осцилляции Chebli, Geitmann, 2007
- 29. Никита Максимов, к.б.н. Саша Подолян, аспирант 1 года Группа репродуктивной физиологии растений МГУ Группа в контакте:
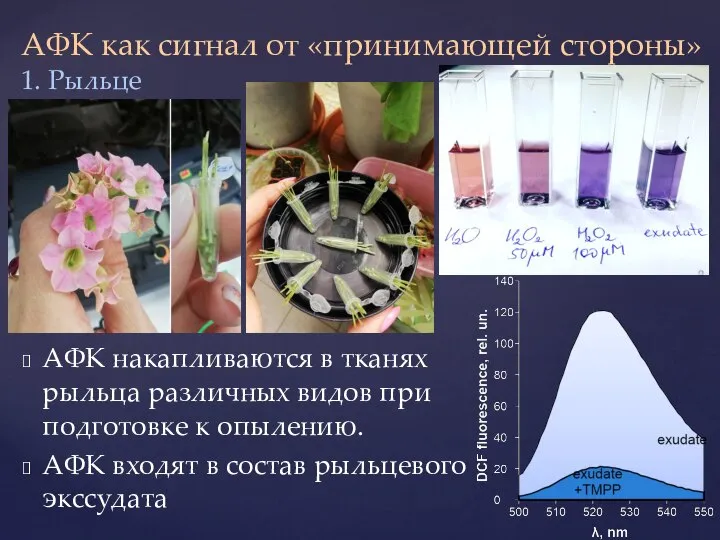
- 30. АФК как сигнал от «принимающей стороны» АФК накапливаются в тканях рыльца различных видов при подготовке к
- 31. Проточная цитометрия: как? a b c Что такое облако протопластов?! Данные получены в сотрудничестве с Е.Шиловым
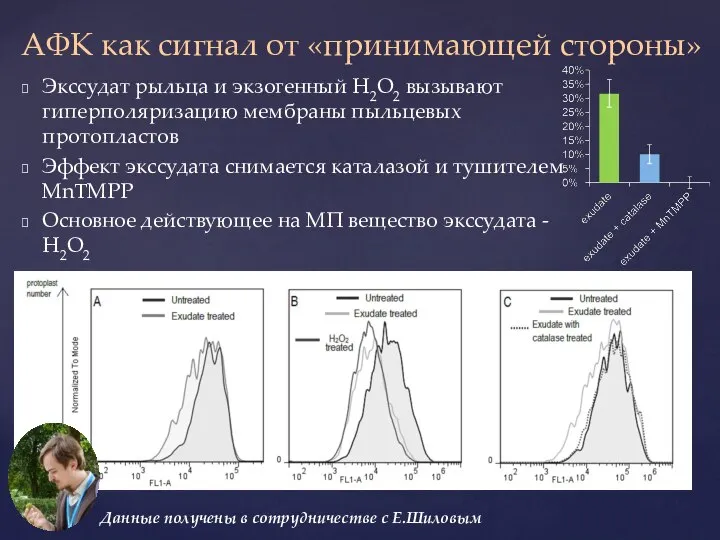
- 32. Экссудат рыльца и экзогенный Н2О2 вызывают гиперполяризацию мембраны пыльцевых протопластов Эффект экссудата снимается каталазой и тушителем
- 33. АФК как сигнал: H2O2 активирует Ca2+ ток и вход Ca2+ в цитоплазму Breygina et al., 2016.
- 34. АФК как сигнал: H2O2 активирует ток K+ Сравнение средней плотности тока при (в % от контроля).
- 35. АФК как сигнал: H2O2 убирает в трубках «щелочной поясок» В растущих трубках лилии вслед за массированным
- 36. Два оптических метода оценки Em Ратиометрический быстрый краситель Di-4-ANEPPS: Съемка в двух каналах при возбуждении синим
- 37. Количественная флуоресцентная микроскопия: динамика эффекта Проточная цитометрия с красителем DiBAC4(3): исследование большой популяции протопластов АФК как
- 38. Гиперполяризация плазмалеммы при действии H2O2 (10 μМ) - распределение После 10 минут инкубации происходит достоверное смещение
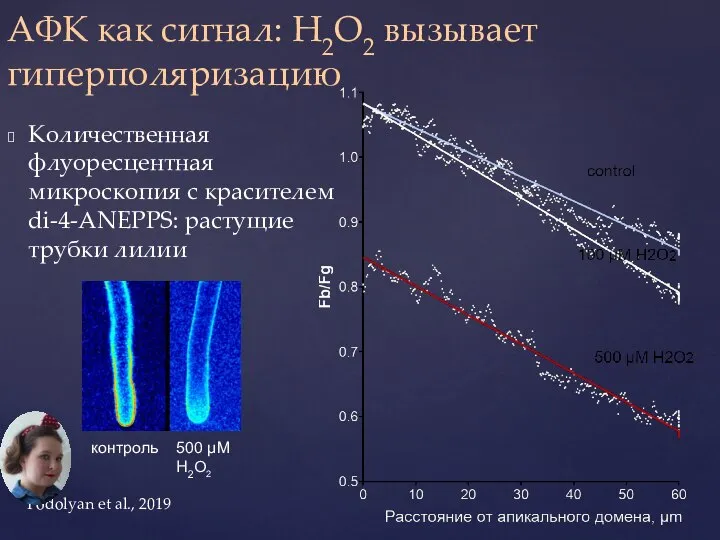
- 39. АФК как сигнал: H2O2 вызывает гиперполяризацию Количественная флуоресцентная микроскопия с красителем di-4-ANEPPS: растущие трубки лилии контроль
- 40. Ca2+ K+ АДФ + Фн АТФ H+ Cl- H2O2 + + ? ? Em
- 41. В трубке ион-транспортные системы расположены полярно Они формируют в цитоплазме динамичные градиенты концентраций, а на мембране
- 42. Пероксид водорода вызывает изменения протеома пыльцы. Появляются более 50 белков, которых не было в контроле. Основные
- 43. Цветковые VS Хвойные Изученный модельный объект Быстрый рост Униполярный рост Обращенный фонтан Крутой градиент Са2+ Крутой
- 44. Градиент мембранного потенциала табак ель
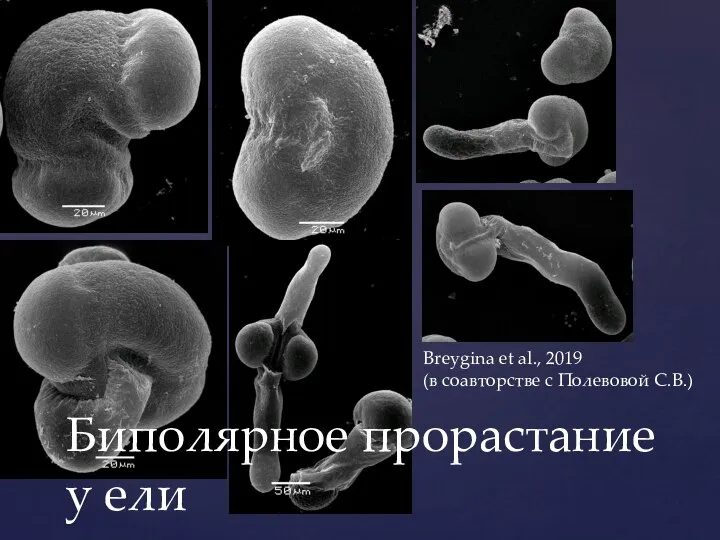
- 45. Биполярное прорастание у ели Breygina et al., 2019 (в соавторстве с Полевовой С.В.)
- 46. Две трубки, а МГМ один. Куда же он пойдет? Он тоже сомневается… b a c d
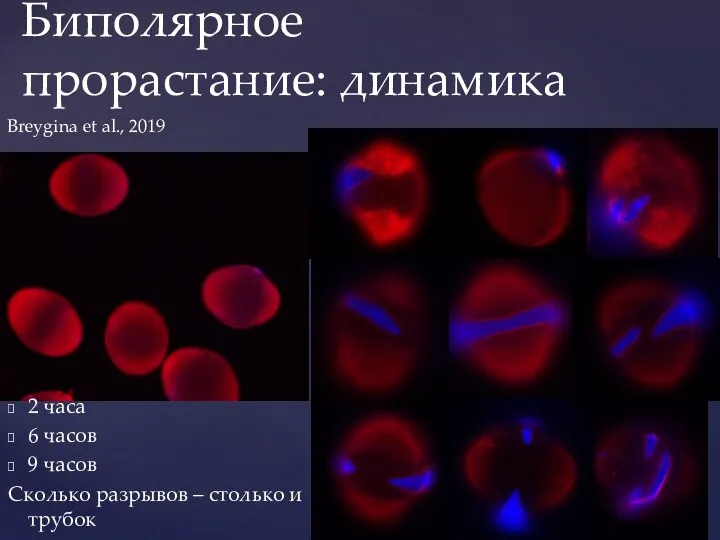
- 47. Биполярное прорастание: динамика 2 часа 6 часов 9 часов Сколько разрывов – столько и трубок Breygina
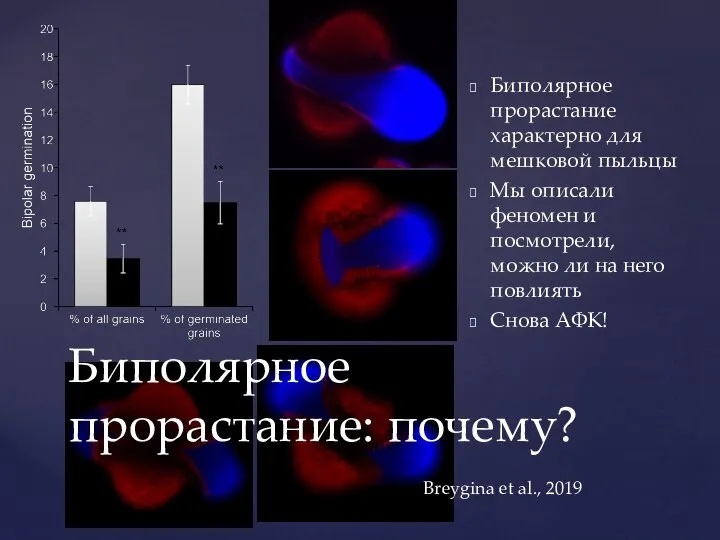
- 48. Биполярное прорастание характерно для мешковой пыльцы Мы описали феномен и посмотрели, можно ли на него повлиять
- 49. Никита Максимов, к.б.н. Саша Подолян, аспирант 1 года Группа репродуктивной физиологии растений МГУ Группа в контакте:
- 51. Скачать презентацию
Слайд 2Скорость роста – до 1 см/час!
Длина – до десятков сантиметров!
Безошибочная «адресная» доставка
Скорость роста – до 1 см/час!
Длина – до десятков сантиметров!
Безошибочная «адресная» доставка
Слайд 31824
1847
Ассирийские священники и египетские боги изображались в процессе ритуального опыления пальмы (5000
1824
1847
Ассирийские священники и египетские боги изображались в процессе ритуального опыления пальмы (5000

Слайд 4Что представляет собой пыльца? Мужской гаметофит – гаплоидное поколение семенных растений, у
Что представляет собой пыльца? Мужской гаметофит – гаплоидное поколение семенных растений, у
Слайд 5Жизнь пыльцы
Жизнь пыльцы
Слайд 6Пыльцевая трубка как модельный объект
Гаплоидный геном – легко выявить важные гены
Отсутствие межклеточных
Пыльцевая трубка как модельный объект
Гаплоидный геном – легко выявить важные гены
Отсутствие межклеточных

Слайд 7Ключевые механизмы поддержания полярного роста:
Зональная организация цитоплазмы позволяет каждой органелле быть «в
Ключевые механизмы поддержания полярного роста:
Зональная организация цитоплазмы позволяет каждой органелле быть «в

Слайд 8Везикулярный транспорт и движение органелл по цитоскелету: доставка стройматериалов и энергообеспечение
Клеточная стенка
Везикулярный транспорт и движение органелл по цитоскелету: доставка стройматериалов и энергообеспечение
Клеточная стенка
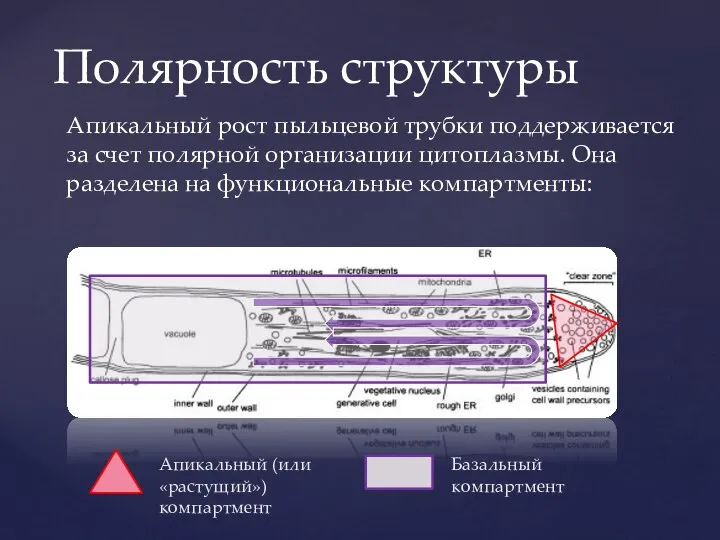
Слайд 9Апикальный (или «растущий») компартмент
Базальный компартмент
Апикальный рост пыльцевой трубки поддерживается за счет полярной
Апикальный (или «растущий») компартмент
Базальный компартмент
Апикальный рост пыльцевой трубки поддерживается за счет полярной
Слайд 10Функции циклоза
1. Транспорт мужского гаметного модуля (MGU).
2. Транспорт в апекс сигнальных молекул
Функции циклоза
1. Транспорт мужского гаметного модуля (MGU).
2. Транспорт в апекс сигнальных молекул
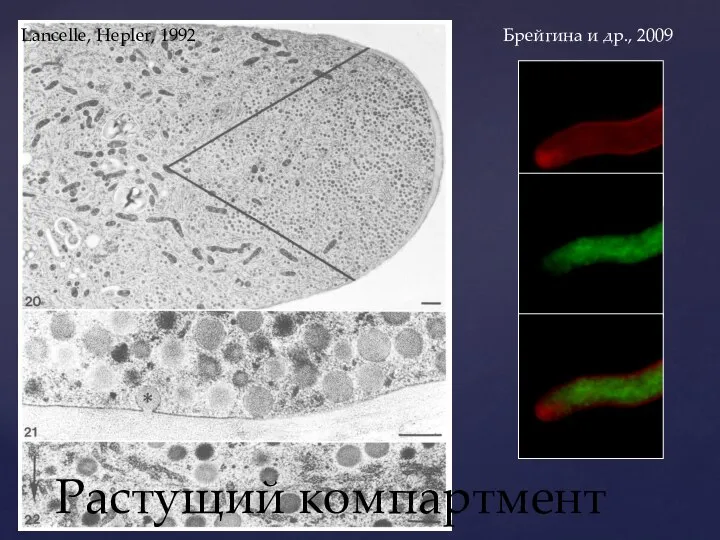
Слайд 11Растущий компартмент
Lancelle, Hepler, 1992
Брейгина и др., 2009
Растущий компартмент
Lancelle, Hepler, 1992
Брейгина и др., 2009
Слайд 12Цитоскелет
Цитоскелет
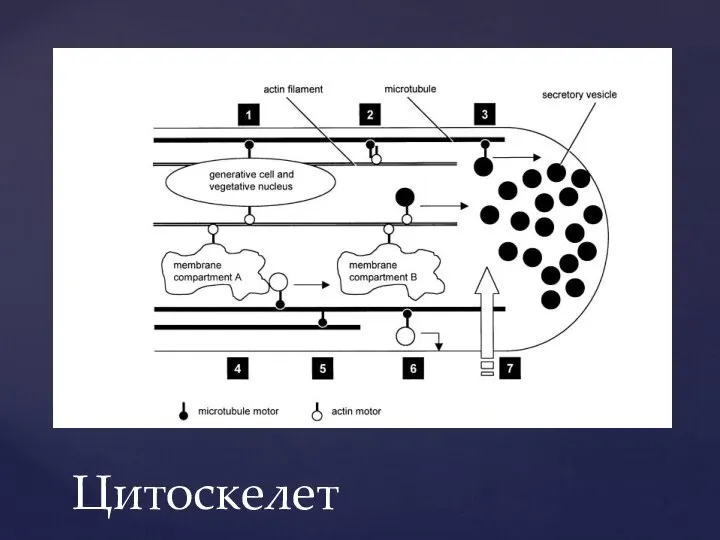
Слайд 13Актиновый цитоскелет играет ключевую роль в поддержании полярного роста
Продольные актиновые тяжи обеспечивают
Актиновый цитоскелет играет ключевую роль в поддержании полярного роста
Продольные актиновые тяжи обеспечивают

Слайд 14Микротрубочки
Отвечает за транспорт спермиев к зародышевому мешку
Остальные функции неизвестны
Ингибиторы не останавливают рост
Микротрубочки
Отвечает за транспорт спермиев к зародышевому мешку
Остальные функции неизвестны
Ингибиторы не останавливают рост

Слайд 15Рециклирование мембран необходимо для поддержания полярности – «уникальность апекса».
В пыльцевой трубке образование
Рециклирование мембран необходимо для поддержания полярности – «уникальность апекса».
В пыльцевой трубке образование

Слайд 16Поляризованная секреция поддерживается благодаря актиновому цитоскелету, а также аннексину и белкам экзоцистного
Поляризованная секреция поддерживается благодаря актиновому цитоскелету, а также аннексину и белкам экзоцистного

Слайд 17Клеточная стенка:
градиент жесткости
Движущей силой для роста является тургорное давление. Однако, оно
Клеточная стенка:
градиент жесткости
Движущей силой для роста является тургорное давление. Однако, оно

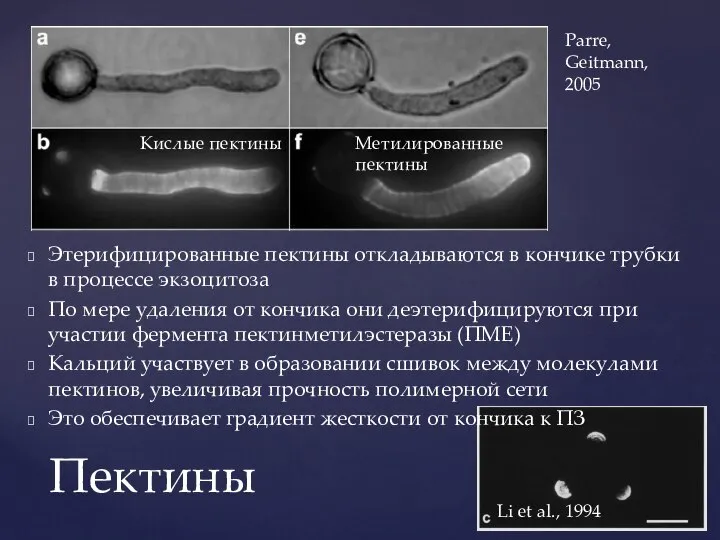
Слайд 18Этерифицированные пектины откладываются в кончике трубки в процессе экзоцитоза
По мере удаления от
Этерифицированные пектины откладываются в кончике трубки в процессе экзоцитоза
По мере удаления от
Слайд 19Каллоза
Breygina et al., 2012
Xie et al., 2012
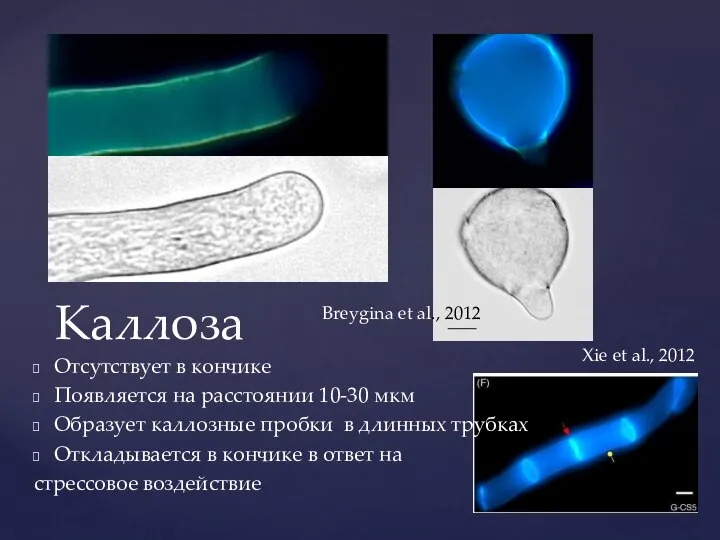
Отсутствует в кончике
Появляется на расстоянии 10-30
Каллоза
Breygina et al., 2012
Xie et al., 2012
Отсутствует в кончике
Появляется на расстоянии 10-30
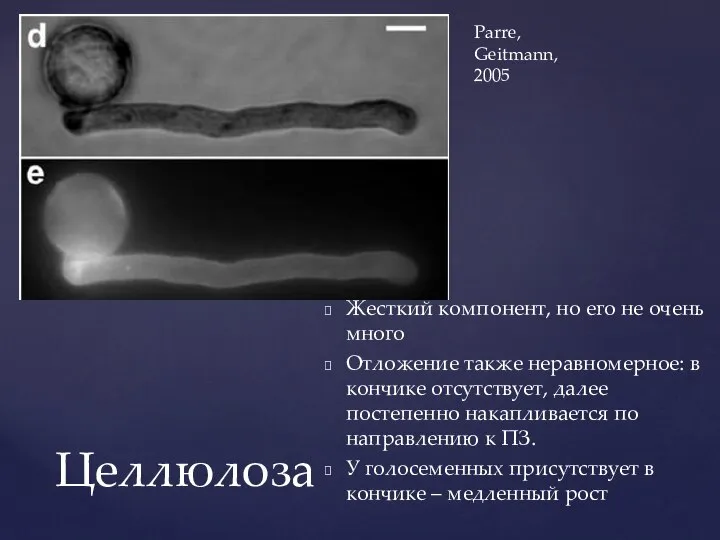
Слайд 20Жесткий компонент, но его не очень много
Отложение также неравномерное: в кончике отсутствует,
Жесткий компонент, но его не очень много
Отложение также неравномерное: в кончике отсутствует,
Слайд 21Регуляторные механизмы: кальций
Hepler, 2012
Кальций входит в кончике трубки через каналы , а
Регуляторные механизмы: кальций
Hepler, 2012
Кальций входит в кончике трубки через каналы , а

Слайд 22Кальциевый градиент определяет место слияния везикул с ПМ и, таким образом, задает
Кальциевый градиент определяет место слияния везикул с ПМ и, таким образом, задает

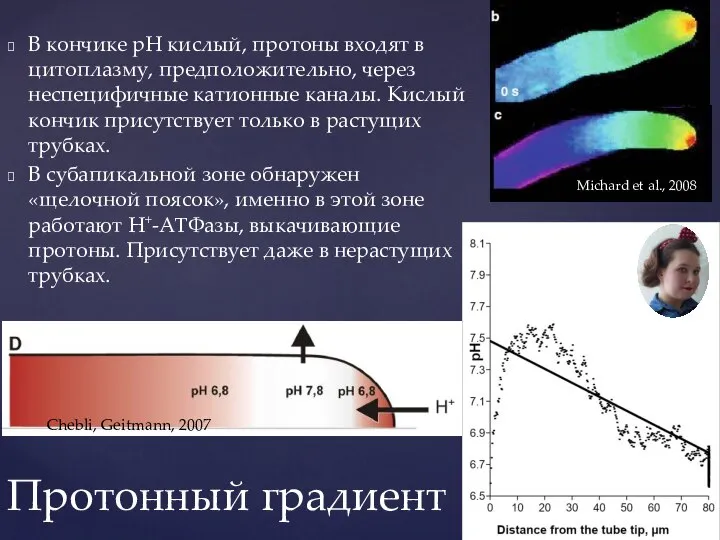
Слайд 23В кончике рН кислый, протоны входят в цитоплазму, предположительно, через неспецифичные катионные
В кончике рН кислый, протоны входят в цитоплазму, предположительно, через неспецифичные катионные
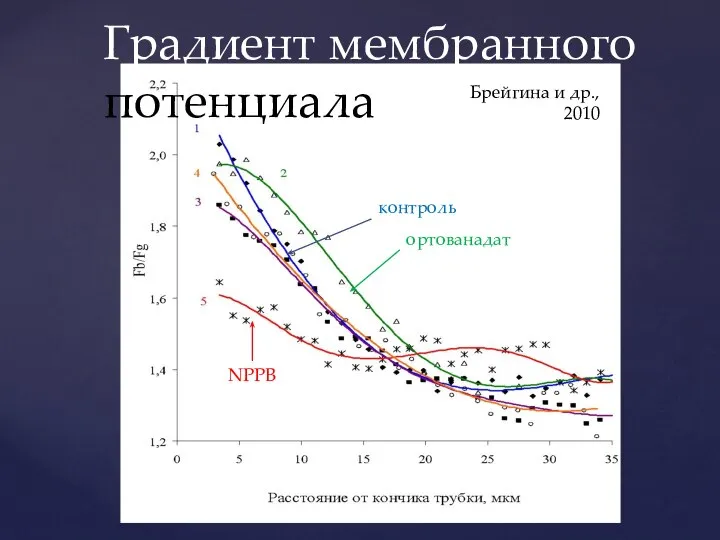
Слайд 24контроль
ортованадат
фузикокцин
Брейгина и др., 2010
Градиент мембранного потенциала
контроль
ортованадат
NPPB
Брейгина и др., 2010
контроль
ортованадат
фузикокцин
Брейгина и др., 2010
Градиент мембранного потенциала
контроль
ортованадат
NPPB
Брейгина и др., 2010
Слайд 25Таким образом, пыльцевая трубка – прекрасная модель для изучения ИОННОЙ РЕГУЛЯЦИИ РОСТА,
Таким образом, пыльцевая трубка – прекрасная модель для изучения ИОННОЙ РЕГУЛЯЦИИ РОСТА,

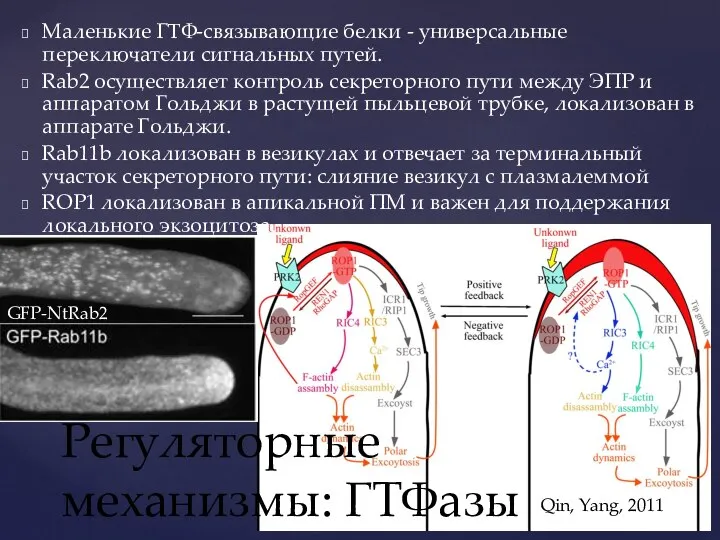
Слайд 26Регуляторные механизмы: ГТФазы
Qin, Yang, 2011
GFP-NtRab2
Маленькие ГТФ-связывающие белки - универсальные переключатели сигнальных путей.
Регуляторные механизмы: ГТФазы
Qin, Yang, 2011
GFP-NtRab2
Маленькие ГТФ-связывающие белки - универсальные переключатели сигнальных путей.
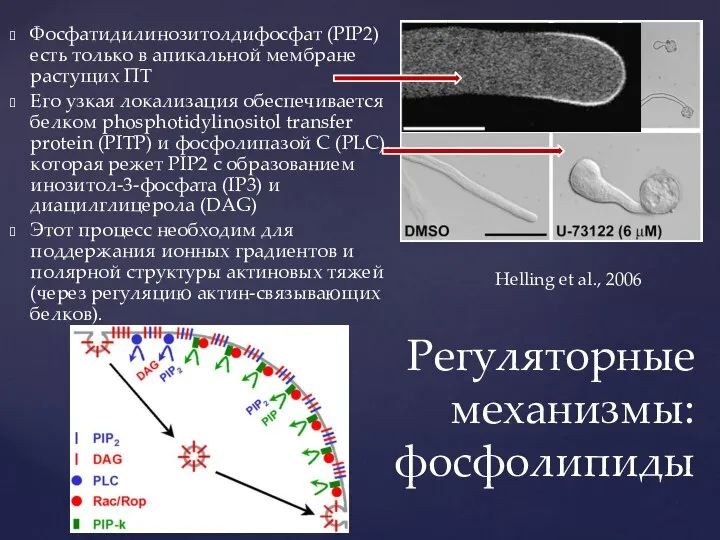
Слайд 27Фосфатидилинозитолдифосфат (PIP2) есть только в апикальной мембране растущих ПТ
Его узкая локализация обеспечивается
Фосфатидилинозитолдифосфат (PIP2) есть только в апикальной мембране растущих ПТ
Его узкая локализация обеспечивается
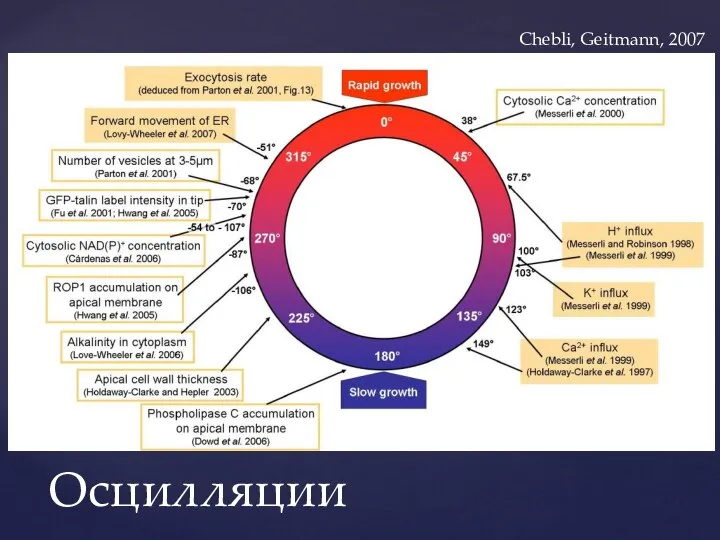
Слайд 28Осцилляции
Chebli, Geitmann, 2007
Осцилляции
Chebli, Geitmann, 2007
Слайд 29Никита Максимов, к.б.н.
Саша Подолян, аспирант 1 года
Группа репродуктивной физиологии растений МГУ
Группа в
Никита Максимов, к.б.н.
Саша Подолян, аспирант 1 года
Группа репродуктивной физиологии растений МГУ
Группа в

Слайд 30АФК как сигнал от «принимающей стороны»
АФК накапливаются в тканях рыльца различных видов
АФК как сигнал от «принимающей стороны»
АФК накапливаются в тканях рыльца различных видов
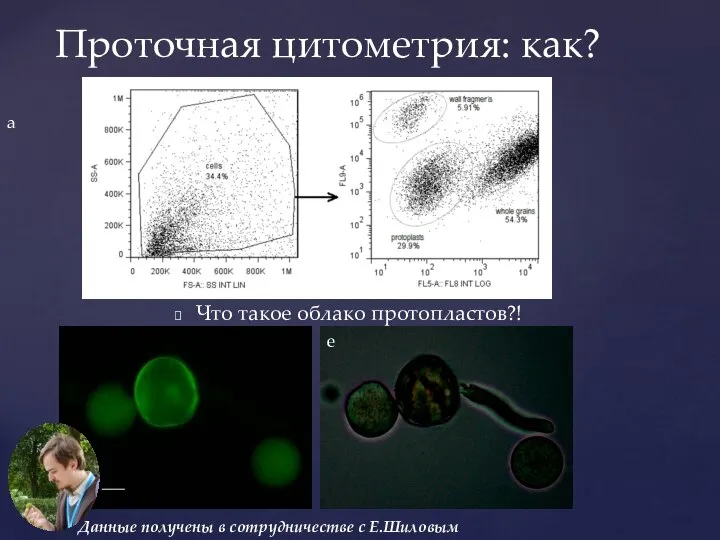
Слайд 31Проточная цитометрия: как?
a
b
c
Что такое облако протопластов?!
Данные получены в сотрудничестве с Е.Шиловым
Проточная цитометрия: как?
a
b
c
Что такое облако протопластов?!
Данные получены в сотрудничестве с Е.Шиловым
Слайд 32Экссудат рыльца и экзогенный Н2О2 вызывают гиперполяризацию мембраны пыльцевых протопластов
Эффект экссудата снимается
Экссудат рыльца и экзогенный Н2О2 вызывают гиперполяризацию мембраны пыльцевых протопластов
Эффект экссудата снимается
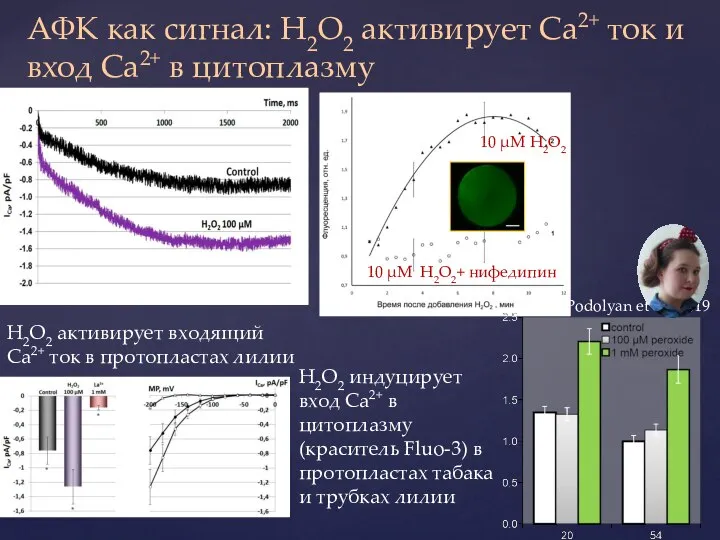
Слайд 33АФК как сигнал: H2O2 активирует Ca2+ ток и вход Ca2+ в цитоплазму
АФК как сигнал: H2O2 активирует Ca2+ ток и вход Ca2+ в цитоплазму
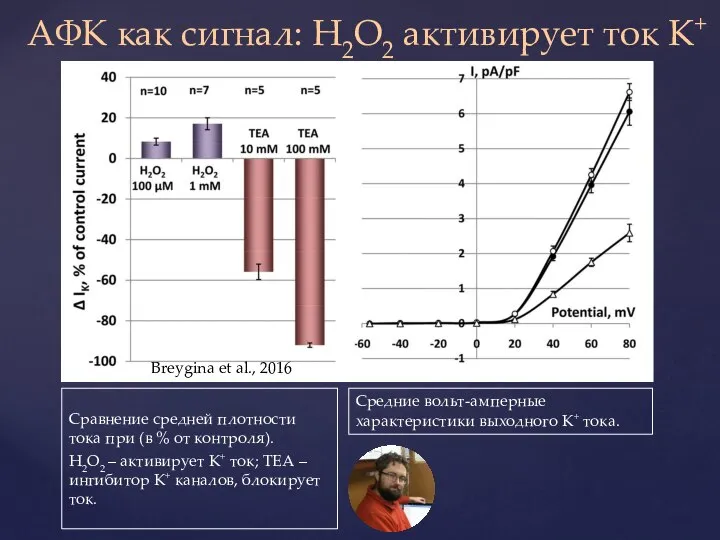
Слайд 34АФК как сигнал: H2O2 активирует ток K+
Сравнение средней плотности тока при (в
АФК как сигнал: H2O2 активирует ток K+
Сравнение средней плотности тока при (в
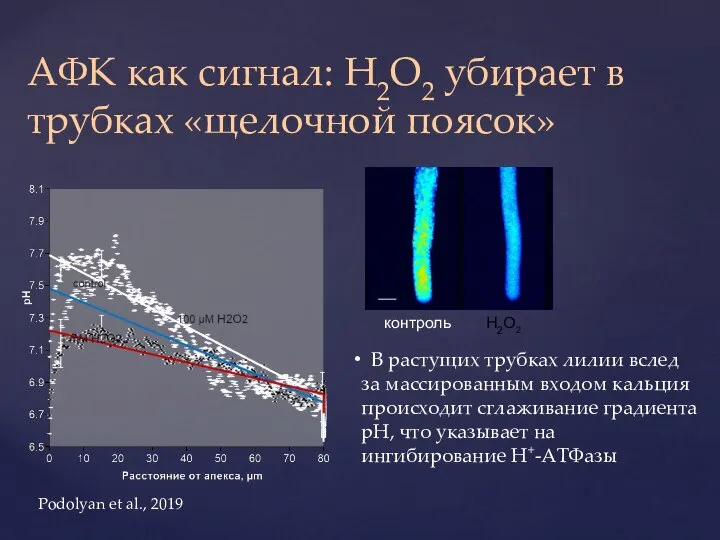
Слайд 35АФК как сигнал: H2O2 убирает в трубках «щелочной поясок»
В растущих трубках
АФК как сигнал: H2O2 убирает в трубках «щелочной поясок»
В растущих трубках
Слайд 36Два оптических метода оценки Em
Ратиометрический быстрый краситель Di-4-ANEPPS:
Съемка в двух каналах при
Два оптических метода оценки Em
Ратиометрический быстрый краситель Di-4-ANEPPS:
Съемка в двух каналах при

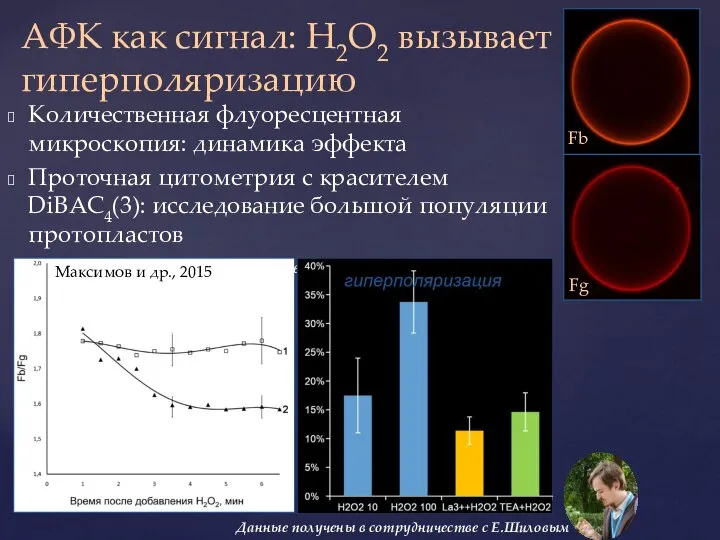
Слайд 37Количественная флуоресцентная микроскопия: динамика эффекта
Проточная цитометрия с красителем DiBAC4(3): исследование большой
Количественная флуоресцентная микроскопия: динамика эффекта
Проточная цитометрия с красителем DiBAC4(3): исследование большой
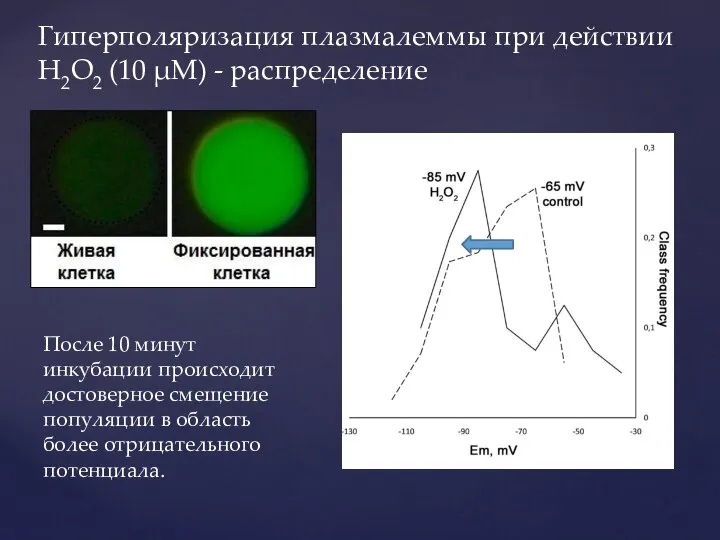
Слайд 38Гиперполяризация плазмалеммы при действии H2O2 (10 μМ) - распределение
После 10 минут инкубации
Гиперполяризация плазмалеммы при действии H2O2 (10 μМ) - распределение
После 10 минут инкубации
Слайд 39АФК как сигнал: H2O2 вызывает гиперполяризацию
Количественная флуоресцентная микроскопия с красителем di-4-ANEPPS: растущие
АФК как сигнал: H2O2 вызывает гиперполяризацию
Количественная флуоресцентная микроскопия с красителем di-4-ANEPPS: растущие
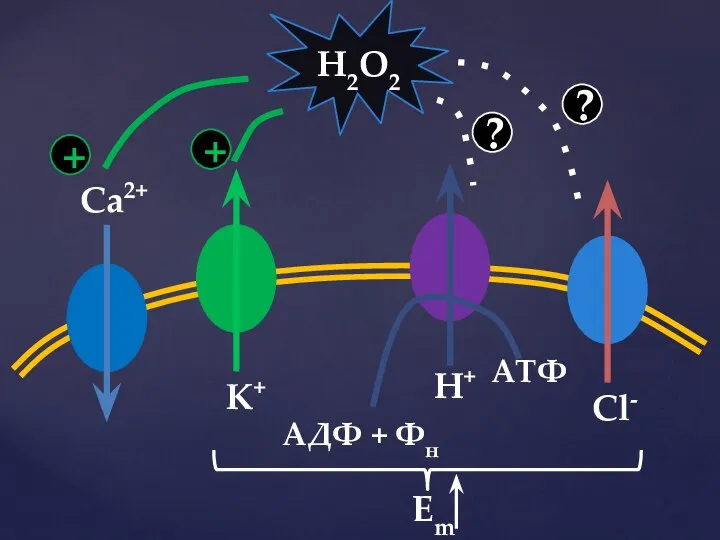
Слайд 40Ca2+
K+
АДФ + Фн
АТФ
H+
Cl-
H2O2
+
+
?
?
Em
Ca2+
K+
АДФ + Фн
АТФ
H+
Cl-
H2O2
+
+
?
?
Em
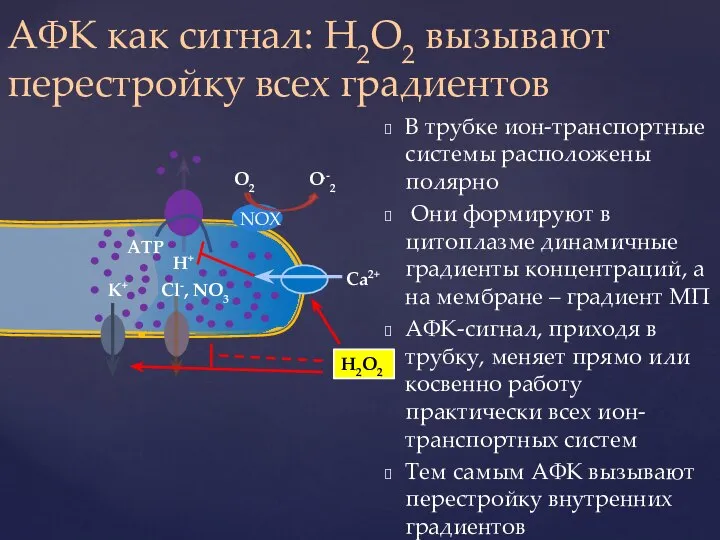
Слайд 41В трубке ион-транспортные системы расположены полярно
Они формируют в цитоплазме динамичные градиенты
В трубке ион-транспортные системы расположены полярно
Они формируют в цитоплазме динамичные градиенты
Слайд 42Пероксид водорода вызывает изменения протеома пыльцы. Появляются более 50 белков, которых не
Пероксид водорода вызывает изменения протеома пыльцы. Появляются более 50 белков, которых не

Слайд 43Цветковые VS Хвойные
Изученный модельный объект
Быстрый рост
Униполярный рост
Обращенный фонтан
Крутой градиент Са2+
Крутой градиент МП
Зависимость
Цветковые VS Хвойные
Изученный модельный объект
Быстрый рост
Униполярный рост
Обращенный фонтан
Крутой градиент Са2+
Крутой градиент МП
Зависимость

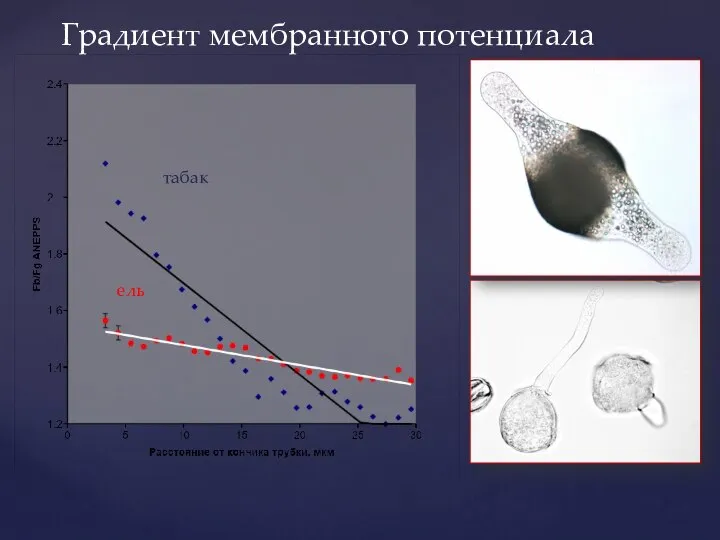
Слайд 44Градиент мембранного потенциала
табак
ель
Градиент мембранного потенциала
табак
ель
Слайд 45Биполярное прорастание у ели
Breygina et al., 2019
(в соавторстве с Полевовой С.В.)
Биполярное прорастание у ели
Breygina et al., 2019
(в соавторстве с Полевовой С.В.)
Слайд 46Две трубки, а МГМ один. Куда же он пойдет?
Он тоже сомневается…
b
a
c
d
Биполярное
Две трубки, а МГМ один. Куда же он пойдет?
Он тоже сомневается…
b
a
c
d
Биполярное

Слайд 47Биполярное прорастание: динамика
2 часа
6 часов
9 часов
Сколько разрывов – столько и трубок
Breygina et
Биполярное прорастание: динамика
2 часа
6 часов
9 часов
Сколько разрывов – столько и трубок
Breygina et
Слайд 48Биполярное прорастание характерно для мешковой пыльцы
Мы описали феномен и посмотрели, можно ли
Биполярное прорастание характерно для мешковой пыльцы
Мы описали феномен и посмотрели, можно ли
Слайд 49Никита Максимов, к.б.н.
Саша Подолян, аспирант 1 года
Группа репродуктивной физиологии растений МГУ
Группа в
Никита Максимов, к.б.н.
Саша Подолян, аспирант 1 года
Группа репродуктивной физиологии растений МГУ
Группа в

 Организм как единая саморазвивающаяся биологическая система
Организм как единая саморазвивающаяся биологическая система Покрытосеменные растения
Покрытосеменные растения Микроскопия. Лекция–визуализация
Микроскопия. Лекция–визуализация Обмен липидов
Обмен липидов Klass_dvustvorchatye
Klass_dvustvorchatye Механические и проводящие ткани
Механические и проводящие ткани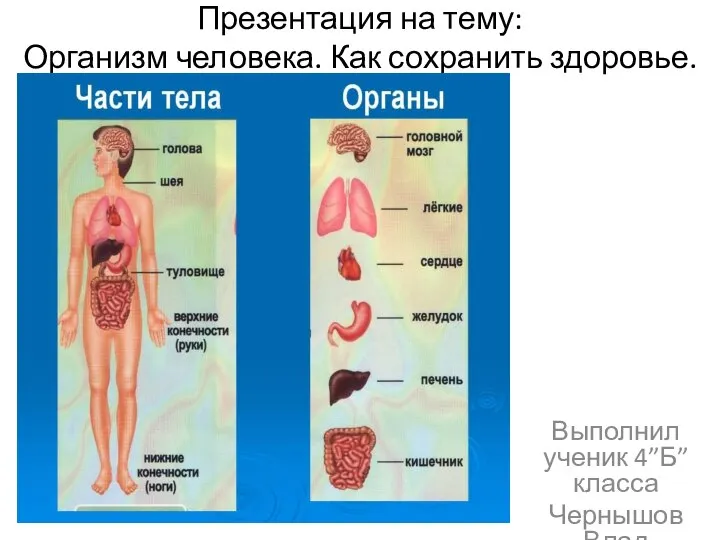 Организм человека. Как сохранить здоровье
Организм человека. Как сохранить здоровье Покрытосеменные Класс двудольные Семейство Паслёновые
Покрытосеменные Класс двудольные Семейство Паслёновые Презентация на тему Физиология клетки
Презентация на тему Физиология клетки  Кабашный. Углеводы.ppt
Кабашный. Углеводы.ppt Тематическое оценивание. Кожа. Выделительная система
Тематическое оценивание. Кожа. Выделительная система Мышцы. Строение мышц
Мышцы. Строение мышц Семейство Ивовые (ива)
Семейство Ивовые (ива) Физиология мотиваций
Физиология мотиваций Строение клетки. Аппарат Гольджи
Строение клетки. Аппарат Гольджи эндокринная система
эндокринная система Съедобные и несъедобные грибы
Съедобные и несъедобные грибы Реакция на раздражители у живых организмов
Реакция на раздражители у живых организмов Ржавчинные и головневые грибы
Ржавчинные и головневые грибы Лесные жители Пермского края
Лесные жители Пермского края Витамин F
Витамин F Зрительный анализатор
Зрительный анализатор Строение и функции Мочевыделительной системы
Строение и функции Мочевыделительной системы Природа и её значение в жизни человека
Природа и её значение в жизни человека ЦНС - центральная нервная система
ЦНС - центральная нервная система Вечнозеленые растения
Вечнозеленые растения Строение зрительного анализатора
Строение зрительного анализатора Сравнение свойств организма человека и животных
Сравнение свойств организма человека и животных