Магнитные изотопные эффекты (миэ) в металл-зависимом ферментативном катализе и их роль в фармакологии. Нанокатиониты
- Магнитные изотопные эффекты (миэ) в металл-зависимом ферментативном катализе и их роль в фармакологии. Нанокатиониты

Содержание
- 2. Магнитный изотопный эффект выражается в зависимости скорости химической реакции (или вероятности рождения молекулы) от ядерного спина,
- 3. Распространенность изотопов биологически активных двухвалентных металлов в природе
- 4. Нанотопология активного сайта креатинкиназы Комбинированное решение уравнений Шредингера и Пуассона Влияние нанотопологии каталитического сайта КК на
- 5. Скорость образования АТФ митохондрией (А) и креатинкиназой (В) как функция изотопа Mg ИНТАКТНАЯ МИТОХОНДРИЯ МИТОХОНДРИЯ, ПОДВЕРГНУТАЯ
- 6. Формирование ион-радикальных пар (Синглет-триплетный каналы фосфорилирования) Магнитный изотопный эффект 25Mg2+ в регуляции митохондриального синтеза АТФ, осуществляемого
- 7. Ион-радикальный механизм глицерофосфаткиназной реакции
- 8. Схема реакции фосфорилирования с помощью АТФ синтазы
- 9. НАНОКАТИОНИТЫ Способны переносить катионы металлов и «отдавать» их в условиях избытка положительных зарядов в окружающей среде
- 10. Структура PMC-16 (Пат. ЕР 1992627А1, 2007)
- 11. ПЕЧЕНЬ МИТОХОНДРИЯ ЦИТОПЛАЗМА ПЛАЗМОЛЕММА ЯДРО Время, ч (имп/мин[59Fe]PMC16 ) / мг белка) х 10-3
- 12. Время, ч МИТОХОНДРИЯ ЦИТОПЛАЗМА ПЛАЗМОЛЕММА ЯДРО МИОКАРД (имп/мин[59Fe]PMC16 ) / мг белка) х 10-3
- 13. ФАРМАКОКИНЕТИКА [Mg]PMC16 (КРЫСЫ) Однократная внутривенная инъекция 20мг/кг (M ± SEM, n = 6) мониторинг в течение
- 14. КАТАЛИТИЧЕСКИЙ САЙТ ФЕРМЕНТА ДОКИНГ РМС16
- 15. МИКРОФОТОГРАФИИ ПЕРИНУКЛЕАРНОЙ ОБЛАСТИ МИОКАРДИОЦИТОВ КРЫСЫ (ПРОСВЕЧИВАЮЩАЯ ЭЛЕКТРОННАЯ МИКРОСКОПИЯ)
- 16. ДИСПЛАЗИЯ МИТОХОНДРИЙ МИОКАРДИОЦИТОВ КРОЛИКА (ИНДУЦИРОВАНА ДОКСОРУБИЦИНОМ) (B) Митохондрия (M): 0.2 DL50 PMC16, 6 часов → 0.5
- 17. A B 25Mg2+ ДИСПЛАЗИЯ ЯДРА МИОКАРДИОЦИТОВ КРОЛИКА (ИНДУЦИРОВАНА ДОКСОРУБИЦИНОМ) 25Mg2+ Гранулярная деструкция матрикса Ядро (N): 0.5
- 18. Перспективы применения в нейробиологии «умных» нанокатионитов на основе порфириновых аддуктов фуллерена С60 Магнитные изотопные эффекты в
- 19. ВОЗМОЖНЫЕ БИОЛОГИЧЕСКИЕ ЭФФЕКТЫ [Mg]PMC16, ПРЕДСКАЗАННЫЕ НА ОСНОВЕ ИХ СТРУКТУРЫ
- 20. Магнитный изотопный эффект 43Ca Y, [(нмоль АТФ/мин)/мг КК]х10-3 A, [имп/мин γ-[32P]АТФ/мг КК]х10-3
- 21. Zn-индуцированный синтез АТФ креатинкиназой
- 22. Zn-индуцированный синтез АТФ пируваткиназой
- 23. Синтез ATP креатинкиназой зависит от изотопии Ca и Zn • с немагнитными изотопами 40Ca (1) и
- 24. Уровень предельных величин замещения магния экзогенными ионами Ca2+ и Zn2+ в молекулах креатинкиназы и ДНК-полимеразы β
- 25. Предпосылки применения МИЭ МИЭ в управлении металл – зависимым ферментативным катализом (А.Л. Бучаченко и соавт., 2005-2013;
- 26. 1 – маркер, кислый гликопротеин плазматической мембраны клеток HeLa, 2 – маркер, гистон Н1А клеток HeLa,
- 27. Очистка и характеристика ДНК-полимеразы β из частично фракционированного хроматина клеток HL-60 А и Б - ИЭФ
- 28. Идентификационные критерии ДНК-полимераз семейства β Молекулярная масса Низкая процессивность фермента Низкая продуктивность (n Mg-зависимый фермент Мономер
- 29. Каталитическая активность бета-подобной ДНК-полимеразы, выделенной из хроматина клеток HL-60: воздействие ингибиторов и KCl
- 30. Зависимость скорости синтеза ДНК ДНК-полимеразой β из клеток HL-60 от концентраций ионов 40Ca2+ и 43Ca2+ A
- 31. Механизм нарушения синтеза ДНК путь
- 32. В отличии от киназ, в случае ДНК-полимераз донором электрона выступает кислород рибозы, а не фосфатной группы.
- 33. Влияние МИЭ - Ме2+ на кинетику катализа, обеспечиваемого ДНК-полимеразой β из клеток HL-60 ([Ме2+]opt=20 mM) *Mg
- 34. Условия инкубации фермента оптимизированы: рН 8.0, + 37 °С, 20 мМ MeCl2, 60 мин ВОЗДЕЙСТВИЕ ИЗОТОПИИ
- 35. n n n Относительная активность ДНК-полимеразы β Изменение каталитической активности ДНК-полимеразы β как проявление МИЭ 43Ca2+
- 36. ЗАВИСИМОСТЬ ОТНОСИТЕЛЬНОЙ КАТАЛИТИЧЕСКОЙ АКТИВНОСТИ И ПРОДУКТИВНОСТИ ДНК-ПОЛИМЕРАЗЫ β ОТ ВЕЛИЧИНЫ ЗАМЕЩЕНИЯ МАГНИЯ ИЗОТОПАМИ КАЛЬЦИЯ ВО ВНУТРИФЕРМЕНТНОМ
- 37. Влияние наночастиц PMC16, транспортирующих изотопы Mg и Ca, на выживаемость клеток (LC50) HL-60 и миелобластов здоровых
- 38. МАКСИМАЛЬНО ДОСТИЖИМЫЙ УРОВЕНЬ ЗАМЕЩЕНИЯ ЭНДОГЕННОГО Mg2+ КАЛЬЦИЕМ В ОЧИЩЕННОЙ ДНК-ПОЛИМЕРАЗЕ БЕТА ИЗ КЛЕТОК HL-60 A –
- 39. Кривые доза-эффект препаратов порфирин-фуллеренов (Ме4[PMC16]) для клеток линии HL60 (миелобластный лейкоз) Цитотоксический эффект магнитных изотопов проявляется
- 40. Распределение опухолевых клеток, окрашенных Аннексином-V/FITC и PI по флуоресценции после инкубации без PMC16(25Mg) (а) и с
- 41. Результаты оценки индукции апоптоза Распределение опухолевых клеток, окрашенных Annexin V/FITC/PI по флуоресценции после инкубации без препарата
- 42. Сравнение LС50 препаратов PMC16(Co), PMC16(24Mg), PMC16(25Mg), p LC50, мг/мл
- 43. Медианы LС50 препаратов LC50, мг/мл LC50, мг/мл LC50, мг/мл LC50 LC50
- 44. PMC16 25Mg2+,43Ca2+ PMC16 PMC16 25Mg2+,43Ca2+ 25Mg2+, 43Ca2+ КК, αФГК, ПК, АТФ-синтаза ∆[дНТФ]↑ Поддержка Анаболизма Перинуклеальные Пермеазы
- 46. Скачать презентацию
Слайд 2Магнитный изотопный эффект
выражается в зависимости скорости химической реакции (или вероятности рождения молекулы)
от
Магнитный изотопный эффект
выражается в зависимости скорости химической реакции (или вероятности рождения молекулы)
от

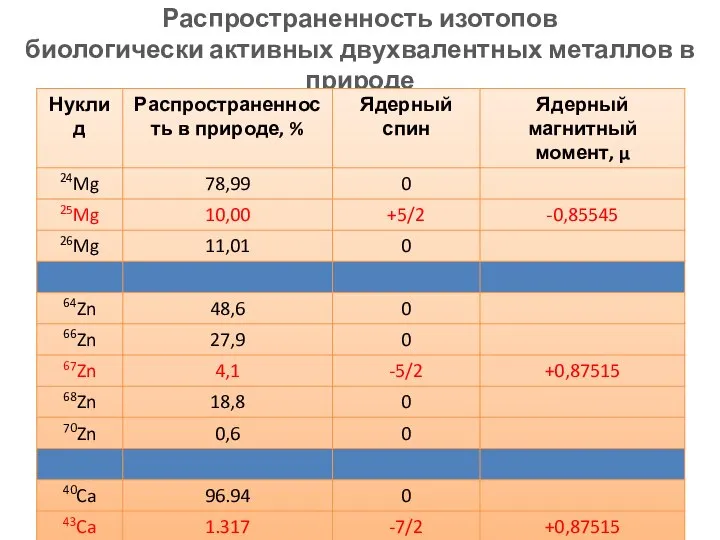
Слайд 3Распространенность изотопов
биологически активных двухвалентных металлов в природе
Распространенность изотопов
биологически активных двухвалентных металлов в природе
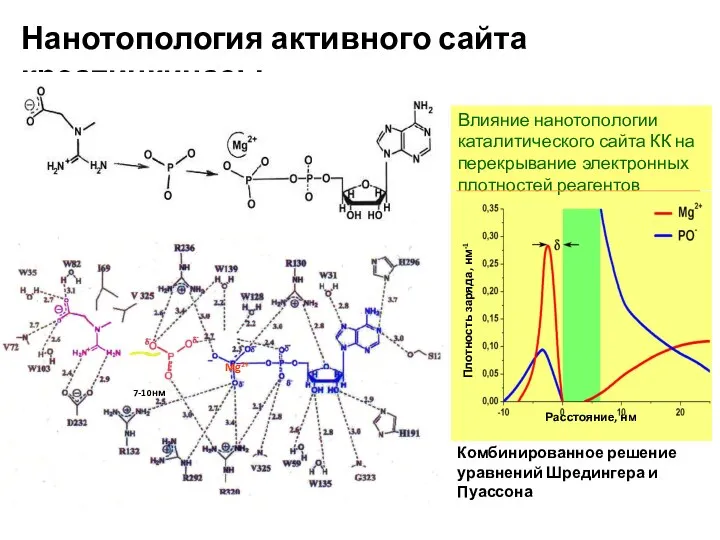
Слайд 4Нанотопология активного сайта креатинкиназы
Комбинированное решение уравнений Шредингера и Пуассона
Влияние нанотопологии каталитического сайта
Нанотопология активного сайта креатинкиназы
Комбинированное решение уравнений Шредингера и Пуассона
Влияние нанотопологии каталитического сайта
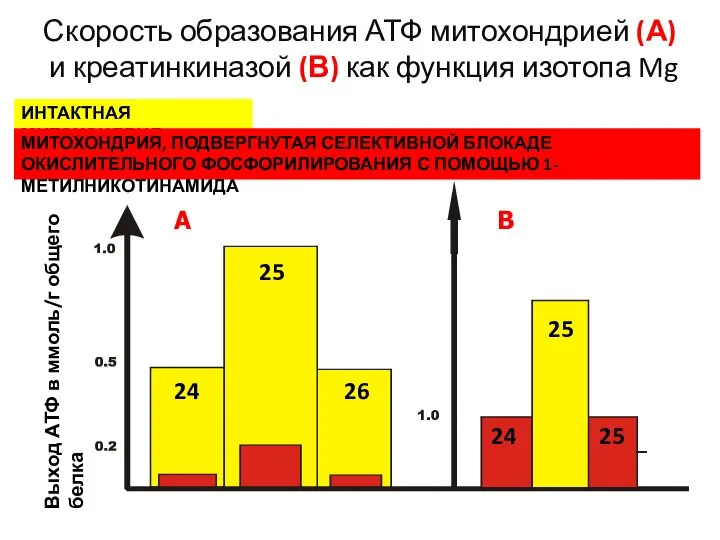
Слайд 5Скорость образования АТФ митохондрией (А)
и креатинкиназой (В) как функция изотопа Mg
Скорость образования АТФ митохондрией (А)
и креатинкиназой (В) как функция изотопа Mg
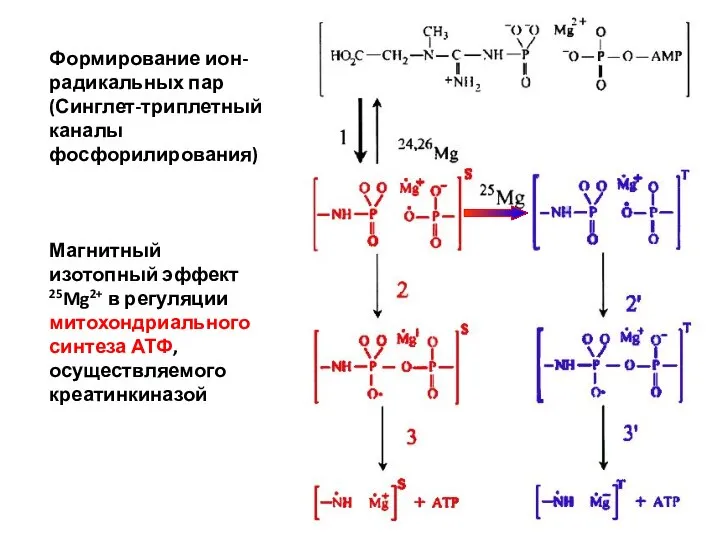
Слайд 6Формирование ион-радикальных пар
(Синглет-триплетный каналы фосфорилирования)
Магнитный изотопный эффект 25Mg2+ в регуляции митохондриального синтеза
Формирование ион-радикальных пар
(Синглет-триплетный каналы фосфорилирования)
Магнитный изотопный эффект 25Mg2+ в регуляции митохондриального синтеза
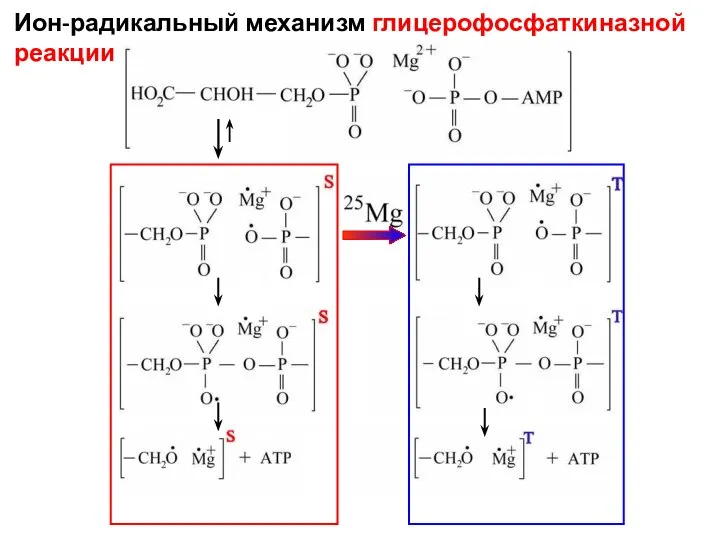
Слайд 7Ион-радикальный механизм глицерофосфаткиназной реакции
Ион-радикальный механизм глицерофосфаткиназной реакции
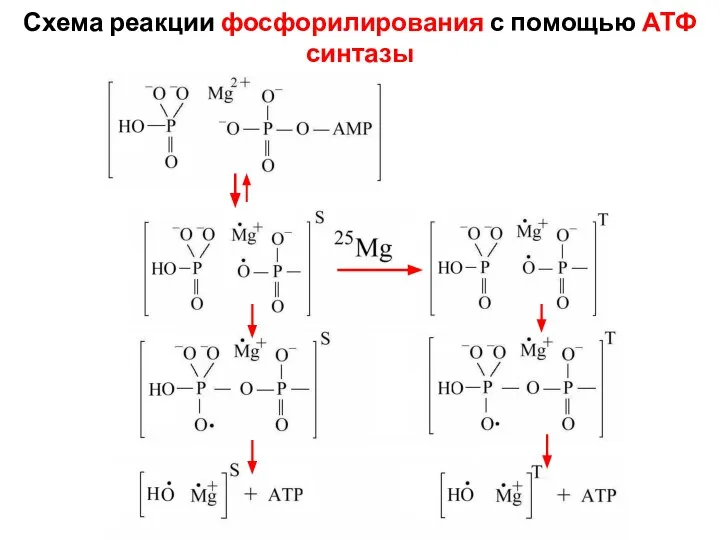
Слайд 8Схема реакции фосфорилирования с помощью АТФ синтазы
Схема реакции фосфорилирования с помощью АТФ синтазы
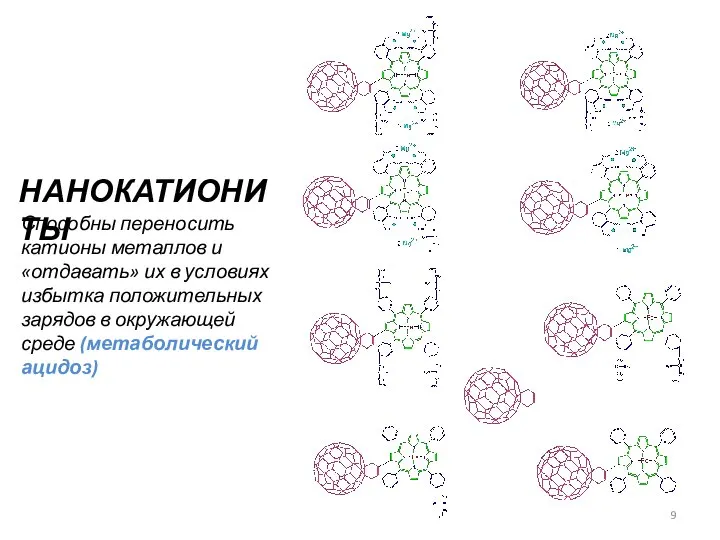
Слайд 9НАНОКАТИОНИТЫ
Способны переносить катионы металлов и «отдавать» их в условиях избытка положительных зарядов
НАНОКАТИОНИТЫ
Способны переносить катионы металлов и «отдавать» их в условиях избытка положительных зарядов
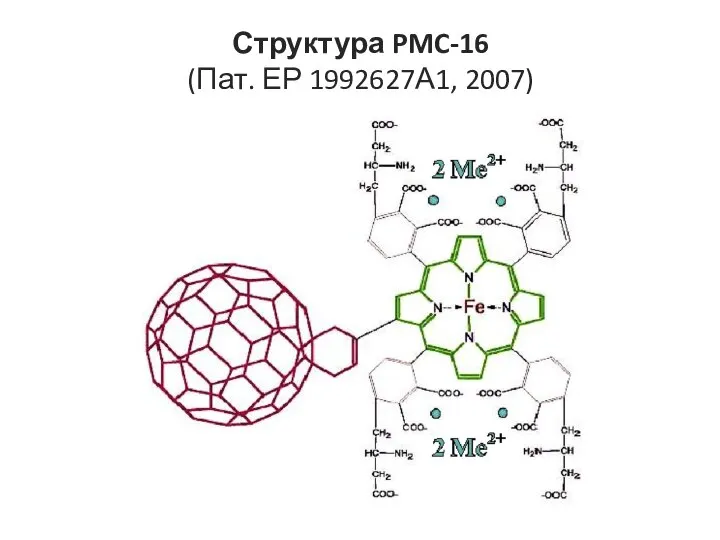
Слайд 10Структура PMC-16
(Пат. ЕР 1992627А1, 2007)
Структура PMC-16
(Пат. ЕР 1992627А1, 2007)
Слайд 11ПЕЧЕНЬ
МИТОХОНДРИЯ
ЦИТОПЛАЗМА
ПЛАЗМОЛЕММА
ЯДРО
Время, ч
(имп/мин[59Fe]PMC16 ) / мг белка) х 10-3
ПЕЧЕНЬ
МИТОХОНДРИЯ
ЦИТОПЛАЗМА
ПЛАЗМОЛЕММА
ЯДРО
Время, ч
(имп/мин[59Fe]PMC16 ) / мг белка) х 10-3
![ПЕЧЕНЬ МИТОХОНДРИЯ ЦИТОПЛАЗМА ПЛАЗМОЛЕММА ЯДРО Время, ч (имп/мин[59Fe]PMC16 ) / мг белка) х 10-3](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1064917/slide-10.jpg)
Слайд 12Время, ч
МИТОХОНДРИЯ
ЦИТОПЛАЗМА
ПЛАЗМОЛЕММА
ЯДРО
МИОКАРД
(имп/мин[59Fe]PMC16 ) / мг белка) х 10-3
Время, ч
МИТОХОНДРИЯ
ЦИТОПЛАЗМА
ПЛАЗМОЛЕММА
ЯДРО
МИОКАРД
(имп/мин[59Fe]PMC16 ) / мг белка) х 10-3
![Время, ч МИТОХОНДРИЯ ЦИТОПЛАЗМА ПЛАЗМОЛЕММА ЯДРО МИОКАРД (имп/мин[59Fe]PMC16 ) / мг белка) х 10-3](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1064917/slide-11.jpg)
Слайд 13ФАРМАКОКИНЕТИКА [Mg]PMC16 (КРЫСЫ)
Однократная внутривенная инъекция 20мг/кг
(M ± SEM, n = 6)
мониторинг
ФАРМАКОКИНЕТИКА [Mg]PMC16 (КРЫСЫ)
Однократная внутривенная инъекция 20мг/кг
(M ± SEM, n = 6)
мониторинг
![ФАРМАКОКИНЕТИКА [Mg]PMC16 (КРЫСЫ) Однократная внутривенная инъекция 20мг/кг (M ± SEM, n =](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1064917/slide-12.jpg)
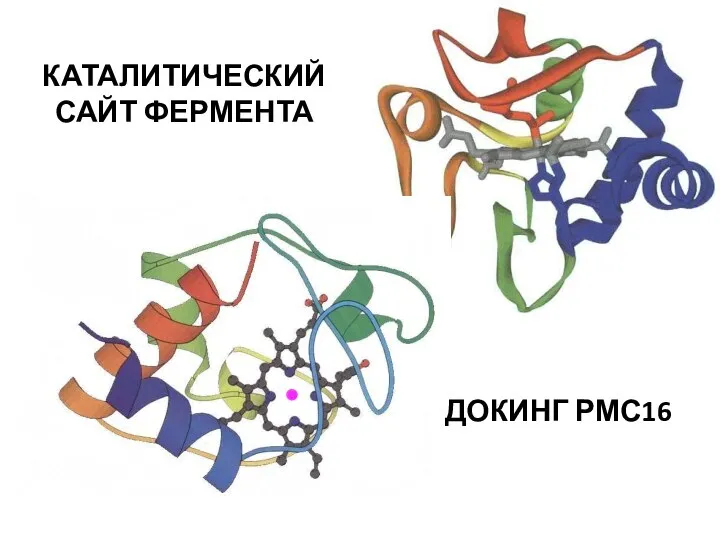
Слайд 14КАТАЛИТИЧЕСКИЙ САЙТ ФЕРМЕНТА
ДОКИНГ РМС16
КАТАЛИТИЧЕСКИЙ САЙТ ФЕРМЕНТА
ДОКИНГ РМС16
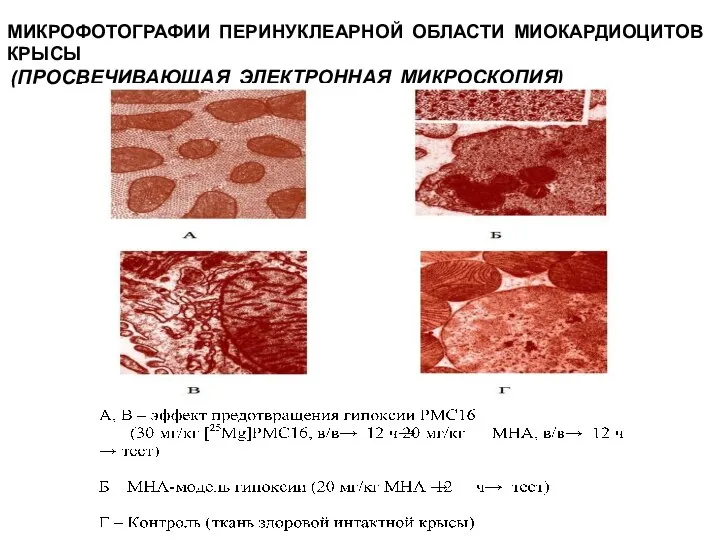
Слайд 15МИКРОФОТОГРАФИИ ПЕРИНУКЛЕАРНОЙ ОБЛАСТИ МИОКАРДИОЦИТОВ КРЫСЫ
(ПРОСВЕЧИВАЮЩАЯ ЭЛЕКТРОННАЯ МИКРОСКОПИЯ)
МИКРОФОТОГРАФИИ ПЕРИНУКЛЕАРНОЙ ОБЛАСТИ МИОКАРДИОЦИТОВ КРЫСЫ
(ПРОСВЕЧИВАЮЩАЯ ЭЛЕКТРОННАЯ МИКРОСКОПИЯ)
Слайд 16ДИСПЛАЗИЯ МИТОХОНДРИЙ МИОКАРДИОЦИТОВ КРОЛИКА
(ИНДУЦИРОВАНА ДОКСОРУБИЦИНОМ)
(B) Митохондрия (M):
0.2 DL50 PMC16, 6
ДИСПЛАЗИЯ МИТОХОНДРИЙ МИОКАРДИОЦИТОВ КРОЛИКА
(ИНДУЦИРОВАНА ДОКСОРУБИЦИНОМ)
(B) Митохондрия (M):
0.2 DL50 PMC16, 6

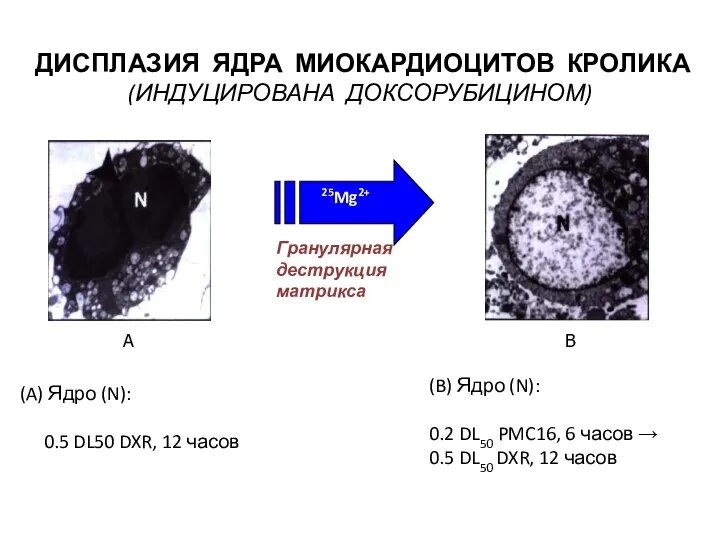
Слайд 17A
B
25Mg2+
ДИСПЛАЗИЯ ЯДРА МИОКАРДИОЦИТОВ КРОЛИКА
(ИНДУЦИРОВАНА ДОКСОРУБИЦИНОМ)
25Mg2+
Гранулярная
деструкция
матрикса
Ядро (N):
0.5 DL50 DXR,
A
B
25Mg2+
ДИСПЛАЗИЯ ЯДРА МИОКАРДИОЦИТОВ КРОЛИКА
(ИНДУЦИРОВАНА ДОКСОРУБИЦИНОМ)
25Mg2+
Гранулярная
деструкция
матрикса
Ядро (N):
0.5 DL50 DXR,
Слайд 18Перспективы применения в нейробиологии «умных» нанокатионитов на основе
порфириновых аддуктов фуллерена С60
Магнитные
Перспективы применения в нейробиологии «умных» нанокатионитов на основе
порфириновых аддуктов фуллерена С60
Магнитные

Слайд 19ВОЗМОЖНЫЕ БИОЛОГИЧЕСКИЕ ЭФФЕКТЫ [Mg]PMC16, ПРЕДСКАЗАННЫЕ НА ОСНОВЕ ИХ СТРУКТУРЫ
ВОЗМОЖНЫЕ БИОЛОГИЧЕСКИЕ ЭФФЕКТЫ [Mg]PMC16, ПРЕДСКАЗАННЫЕ НА ОСНОВЕ ИХ СТРУКТУРЫ
![ВОЗМОЖНЫЕ БИОЛОГИЧЕСКИЕ ЭФФЕКТЫ [Mg]PMC16, ПРЕДСКАЗАННЫЕ НА ОСНОВЕ ИХ СТРУКТУРЫ](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1064917/slide-18.jpg)
Слайд 20Магнитный изотопный эффект 43Ca
Y, [(нмоль АТФ/мин)/мг КК]х10-3
A, [имп/мин γ-[32P]АТФ/мг КК]х10-3
Магнитный изотопный эффект 43Ca
Y, [(нмоль АТФ/мин)/мг КК]х10-3
A, [имп/мин γ-[32P]АТФ/мг КК]х10-3
![Магнитный изотопный эффект 43Ca Y, [(нмоль АТФ/мин)/мг КК]х10-3 A, [имп/мин γ-[32P]АТФ/мг КК]х10-3](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1064917/slide-19.jpg)
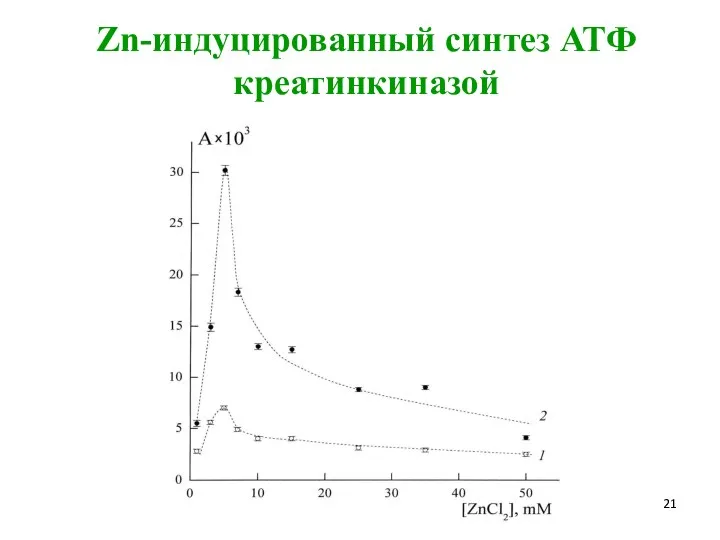
Слайд 21Zn-индуцированный синтез АТФ креатинкиназой
Zn-индуцированный синтез АТФ креатинкиназой
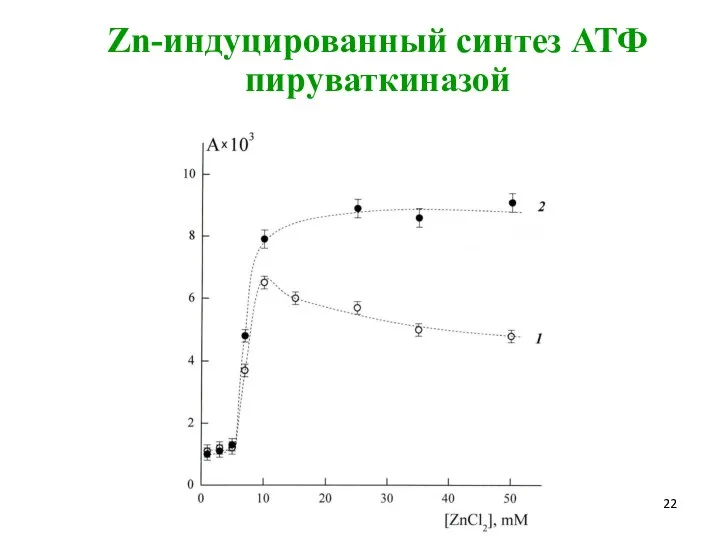
Слайд 22Zn-индуцированный синтез АТФ пируваткиназой
Zn-индуцированный синтез АТФ пируваткиназой
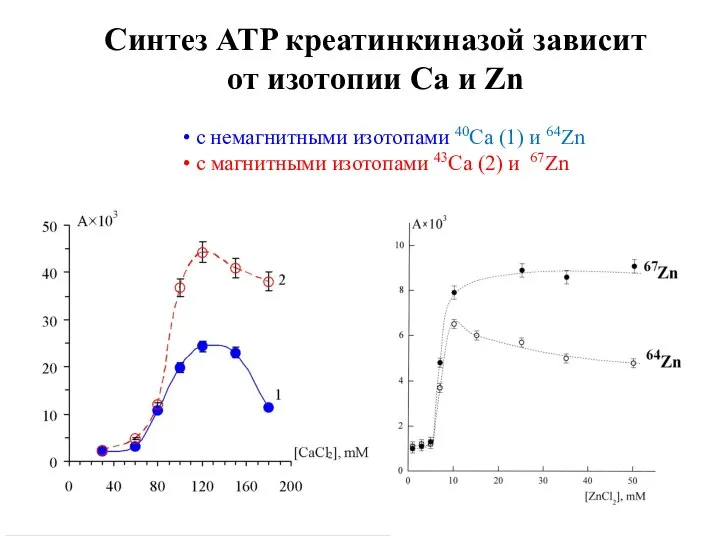
Слайд 23Синтез ATP креатинкиназой зависит
от изотопии Ca и Zn • с немагнитными
Синтез ATP креатинкиназой зависит от изотопии Ca и Zn • с немагнитными
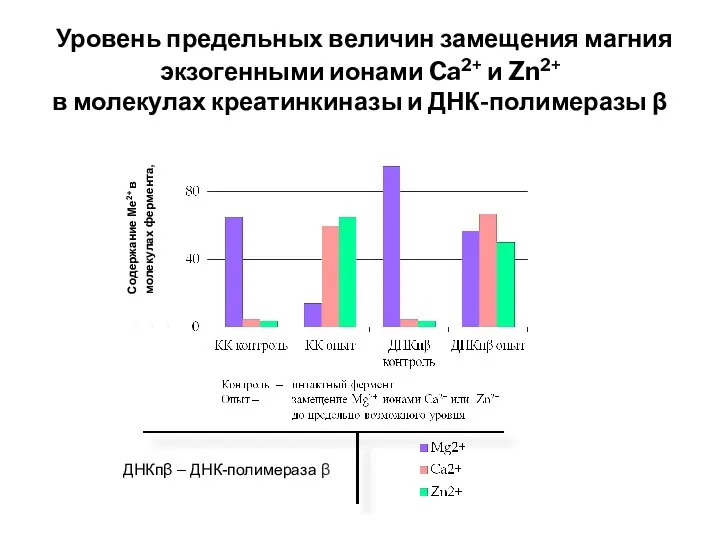
Слайд 24 Уровень предельных величин замещения магния экзогенными ионами Ca2+ и Zn2+
в
Уровень предельных величин замещения магния экзогенными ионами Ca2+ и Zn2+ в
Слайд 25Предпосылки применения МИЭ
МИЭ в управлении
металл – зависимым
ферментативным катализом
(А.Л. Бучаченко и
Предпосылки применения МИЭ
МИЭ в управлении
металл – зависимым
ферментативным катализом
(А.Л. Бучаченко и
Слайд 261 – маркер, кислый гликопротеин плазматической мембраны клеток HeLa,
2 – маркер, гистон
1 – маркер, кислый гликопротеин плазматической мембраны клеток HeLa,
2 – маркер, гистон
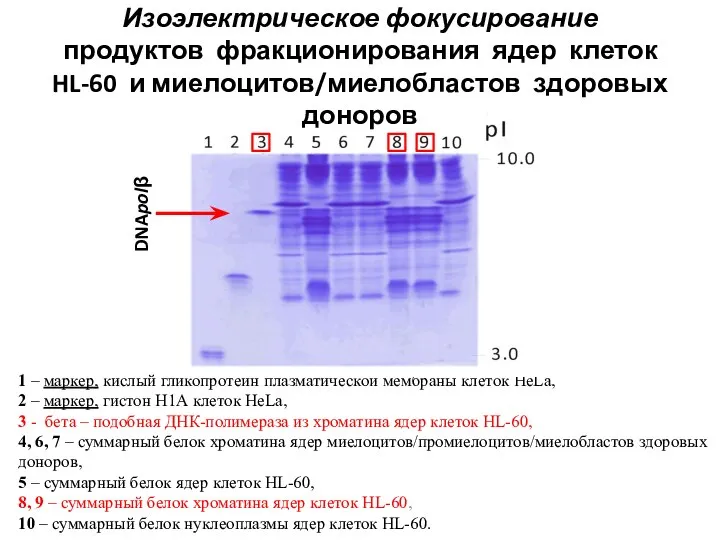
Слайд 27Очистка и характеристика
ДНК-полимеразы β
из частично фракционированного хроматина клеток HL-60
А
Очистка и характеристика
ДНК-полимеразы β
из частично фракционированного хроматина клеток HL-60
А
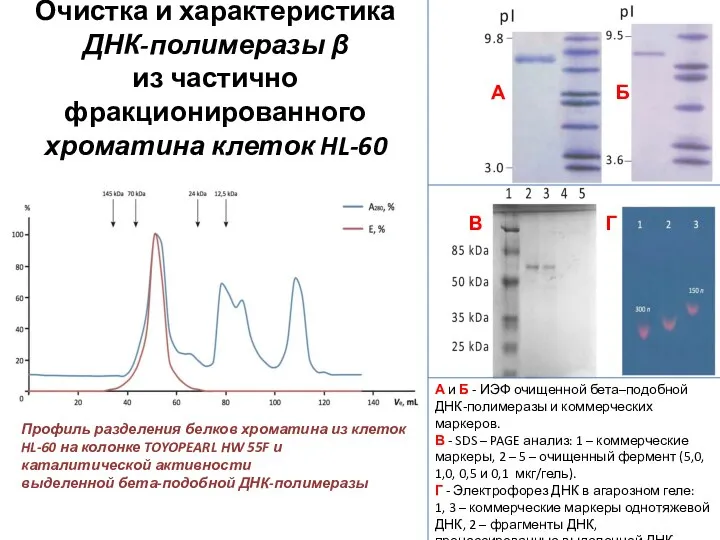
Слайд 28Идентификационные критерии
ДНК-полимераз семейства β
Молекулярная масса <110 кДа
Низкая процессивность фермента
Низкая продуктивность
Идентификационные критерии
ДНК-полимераз семейства β
Молекулярная масса <110 кДа
Низкая процессивность фермента
Низкая продуктивность

Слайд 29Каталитическая активность бета-подобной ДНК-полимеразы,
выделенной из хроматина клеток HL-60:
воздействие ингибиторов и
Каталитическая активность бета-подобной ДНК-полимеразы, выделенной из хроматина клеток HL-60: воздействие ингибиторов и
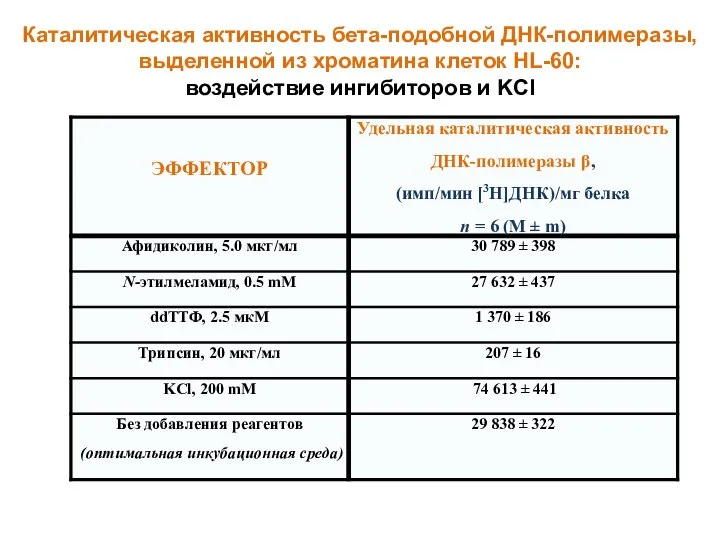
Слайд 30Зависимость скорости синтеза ДНК ДНК-полимеразой β из клеток HL-60
от концентраций ионов
Зависимость скорости синтеза ДНК ДНК-полимеразой β из клеток HL-60
от концентраций ионов
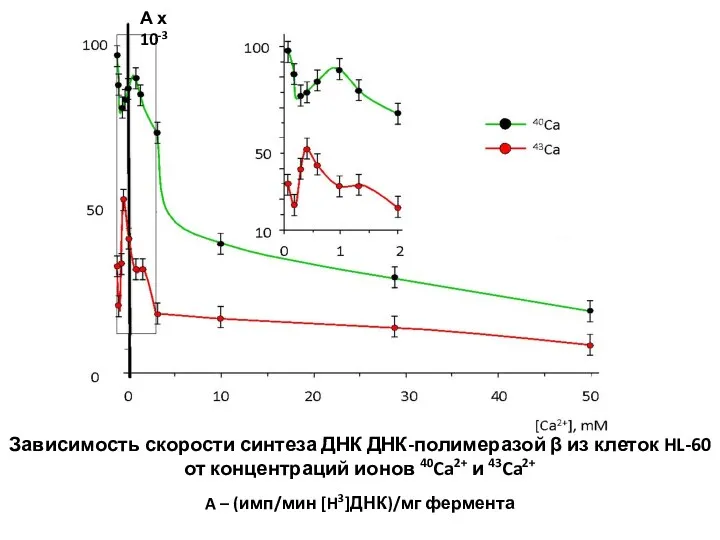
Слайд 31Механизм
нарушения
синтеза ДНК
путь
Механизм
нарушения
синтеза ДНК
путь
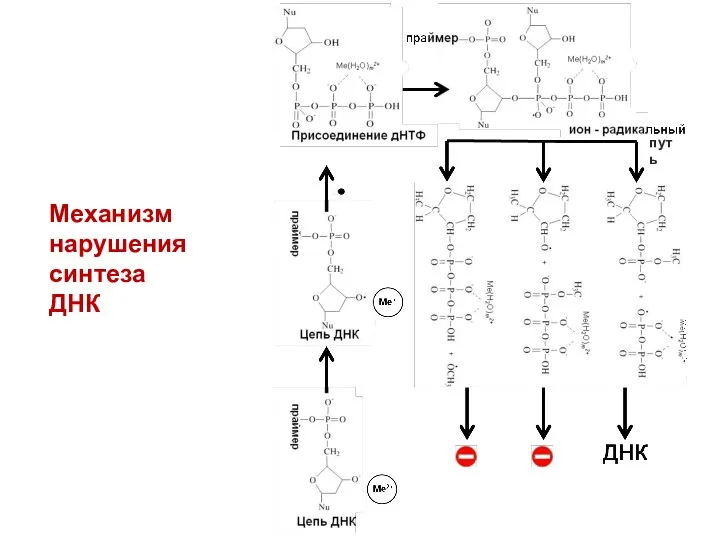
Слайд 32В отличии от киназ, в случае ДНК-полимераз донором электрона выступает кислород рибозы,
В отличии от киназ, в случае ДНК-полимераз донором электрона выступает кислород рибозы,
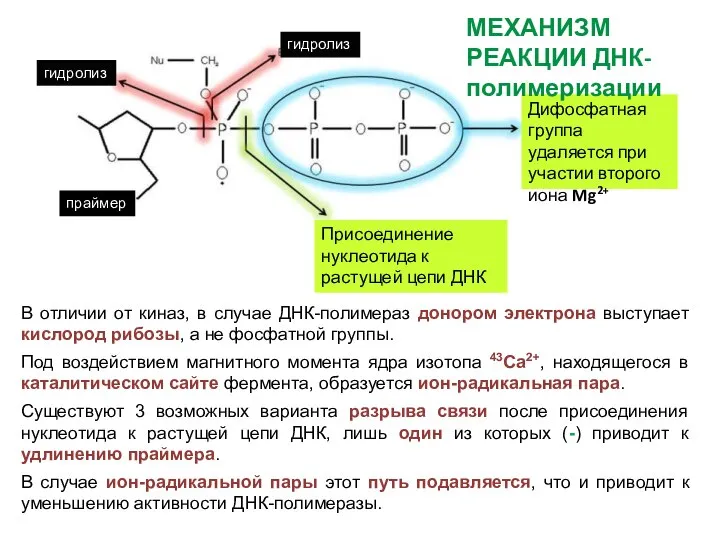
Слайд 33Влияние МИЭ - Ме2+ на кинетику катализа, обеспечиваемого ДНК-полимеразой β из клеток
Влияние МИЭ - Ме2+ на кинетику катализа, обеспечиваемого ДНК-полимеразой β из клеток
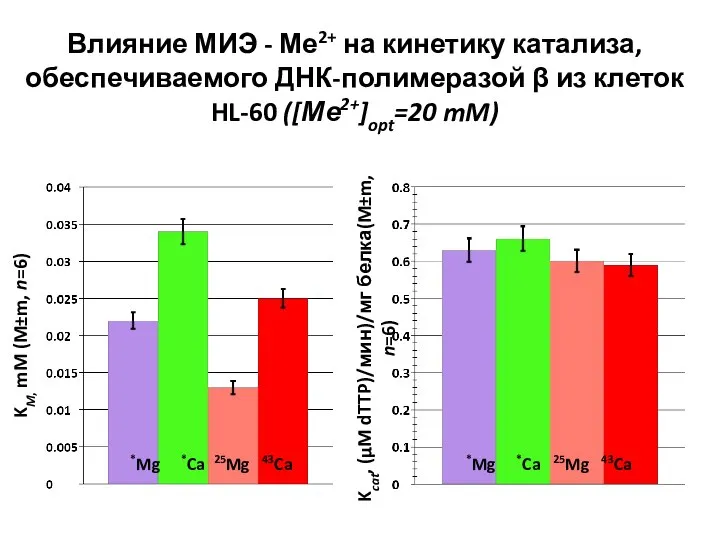
Слайд 34Условия инкубации фермента оптимизированы:
рН 8.0, + 37 °С, 20 мМ MeCl2,
Условия инкубации фермента оптимизированы:
рН 8.0, + 37 °С, 20 мМ MeCl2,
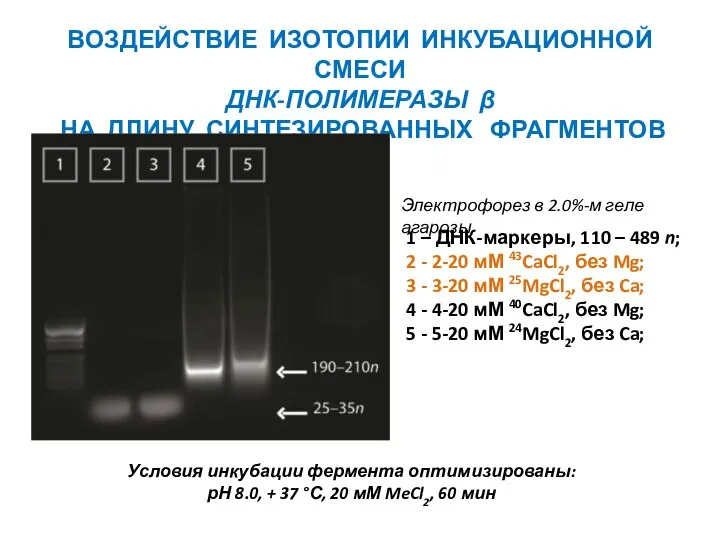
Слайд 35n
n
n
Относительная активность ДНК-полимеразы β
Изменение каталитической активности ДНК-полимеразы β
как проявление МИЭ 43Ca2+
n
n
n
Относительная активность ДНК-полимеразы β
Изменение каталитической активности ДНК-полимеразы β
как проявление МИЭ 43Ca2+
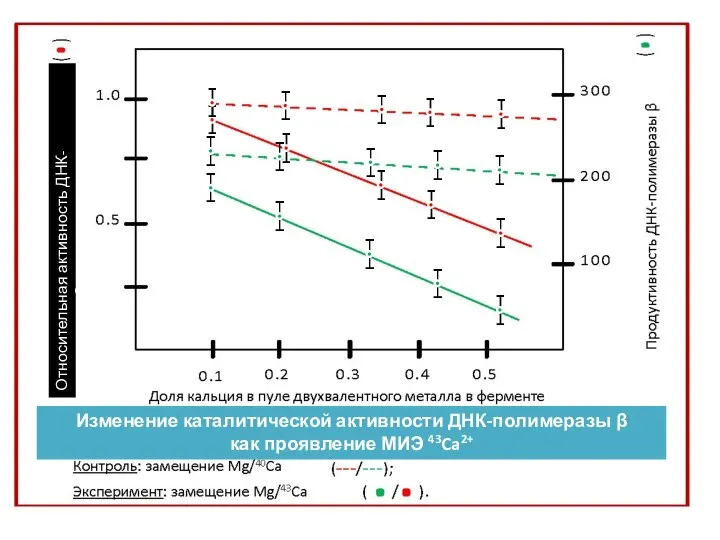
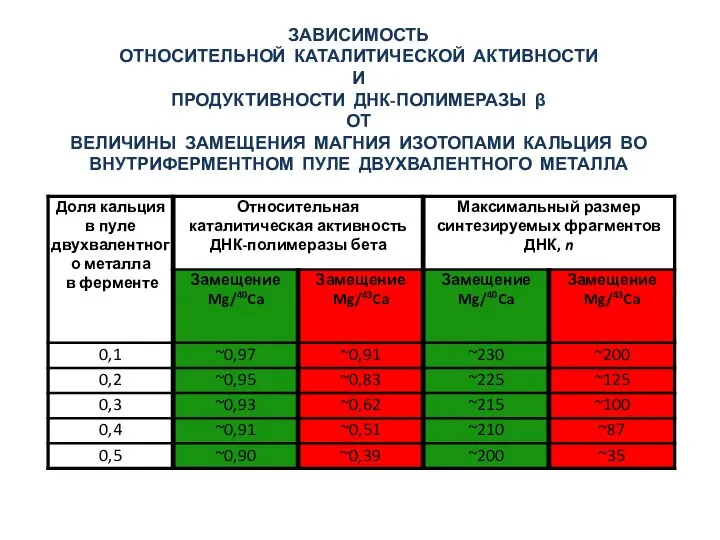
Слайд 36ЗАВИСИМОСТЬ
ОТНОСИТЕЛЬНОЙ КАТАЛИТИЧЕСКОЙ АКТИВНОСТИ
И
ПРОДУКТИВНОСТИ ДНК-ПОЛИМЕРАЗЫ β
ОТ
ВЕЛИЧИНЫ ЗАМЕЩЕНИЯ МАГНИЯ
ЗАВИСИМОСТЬ
ОТНОСИТЕЛЬНОЙ КАТАЛИТИЧЕСКОЙ АКТИВНОСТИ
И
ПРОДУКТИВНОСТИ ДНК-ПОЛИМЕРАЗЫ β
ОТ
ВЕЛИЧИНЫ ЗАМЕЩЕНИЯ МАГНИЯ
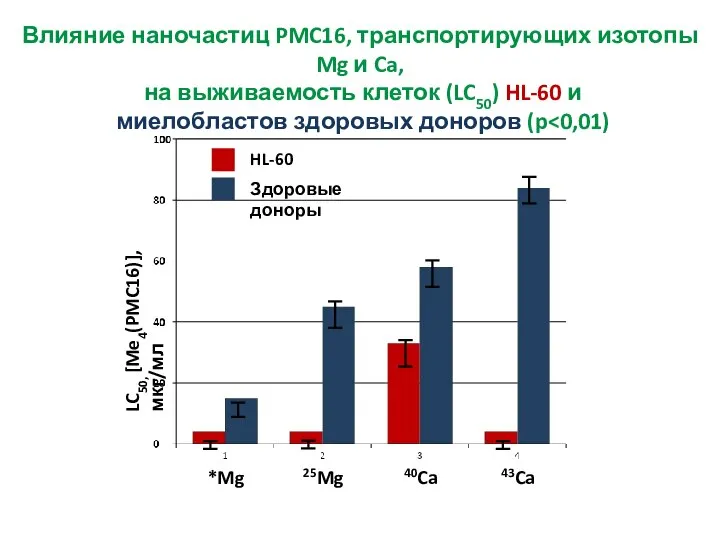
Слайд 37Влияние наночастиц PMC16, транспортирующих изотопы Mg и Ca,
на выживаемость клеток (LC50)
Влияние наночастиц PMC16, транспортирующих изотопы Mg и Ca,
на выживаемость клеток (LC50)
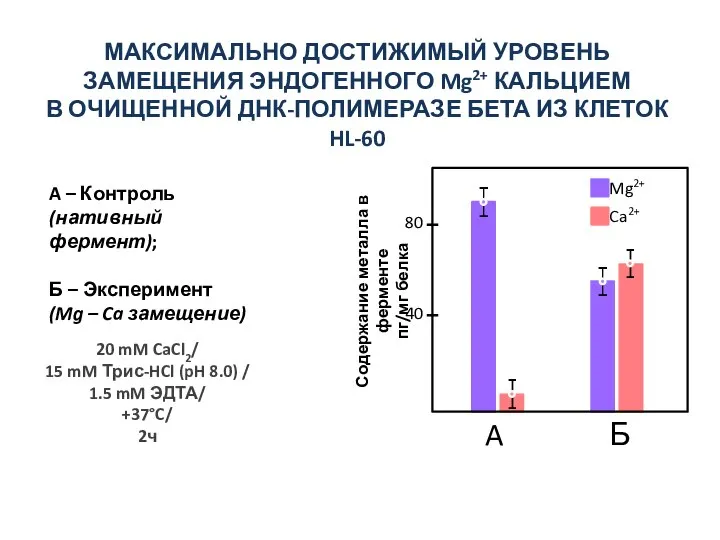
Слайд 38МАКСИМАЛЬНО ДОСТИЖИМЫЙ УРОВЕНЬ
ЗАМЕЩЕНИЯ ЭНДОГЕННОГО Mg2+ КАЛЬЦИЕМ
В ОЧИЩЕННОЙ ДНК-ПОЛИМЕРАЗЕ БЕТА ИЗ
МАКСИМАЛЬНО ДОСТИЖИМЫЙ УРОВЕНЬ
ЗАМЕЩЕНИЯ ЭНДОГЕННОГО Mg2+ КАЛЬЦИЕМ
В ОЧИЩЕННОЙ ДНК-ПОЛИМЕРАЗЕ БЕТА ИЗ
Слайд 39Кривые доза-эффект
препаратов порфирин-фуллеренов (Ме4[PMC16])
для клеток линии HL60 (миелобластный лейкоз)
Цитотоксический
Кривые доза-эффект
препаратов порфирин-фуллеренов (Ме4[PMC16])
для клеток линии HL60 (миелобластный лейкоз)
Цитотоксический
![Кривые доза-эффект препаратов порфирин-фуллеренов (Ме4[PMC16]) для клеток линии HL60 (миелобластный лейкоз) Цитотоксический](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1064917/slide-38.jpg)
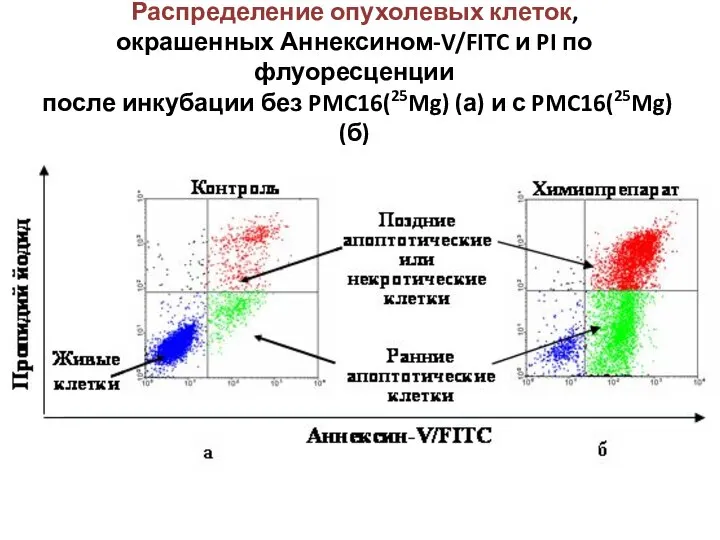
Слайд 40Распределение опухолевых клеток,
окрашенных Аннексином-V/FITC и PI по флуоресценции
после инкубации без
Распределение опухолевых клеток, окрашенных Аннексином-V/FITC и PI по флуоресценции после инкубации без
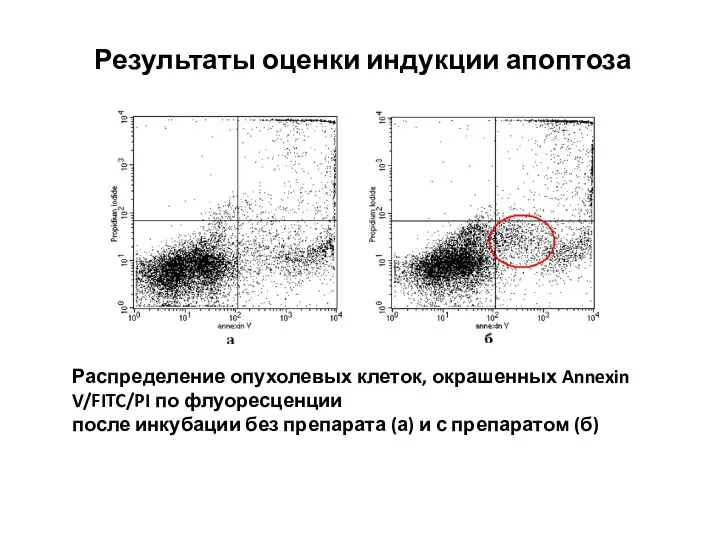
Слайд 41Результаты оценки индукции апоптоза
Распределение опухолевых клеток, окрашенных Annexin V/FITC/PI по флуоресценции
Результаты оценки индукции апоптоза
Распределение опухолевых клеток, окрашенных Annexin V/FITC/PI по флуоресценции
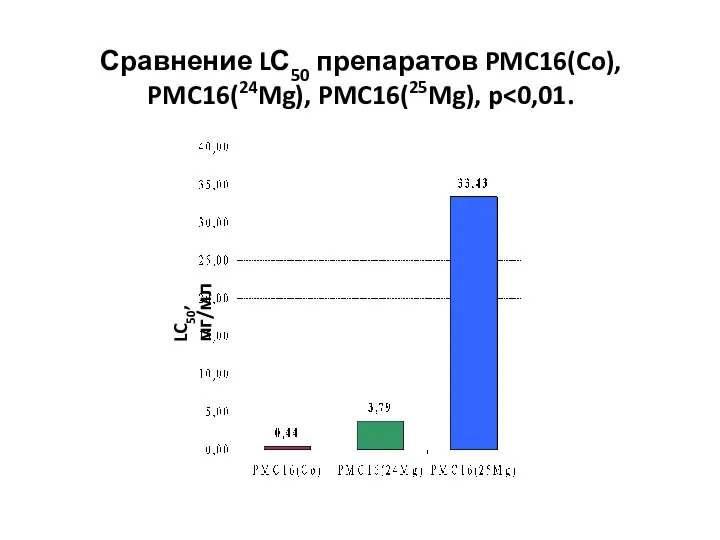
Слайд 42Сравнение LС50 препаратов PMC16(Co), PMC16(24Mg), PMC16(25Mg), p<0,01.
LC50, мг/мл
Сравнение LС50 препаратов PMC16(Co), PMC16(24Mg), PMC16(25Mg), p<0,01.
LC50, мг/мл
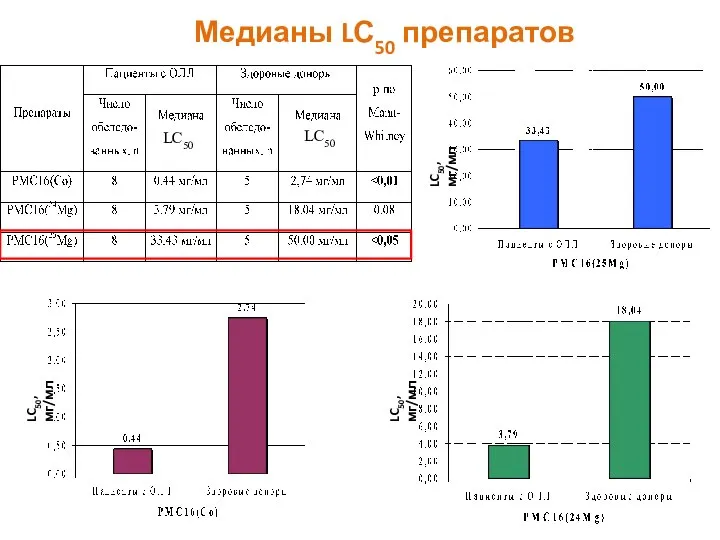
Слайд 43Медианы LС50 препаратов
LC50, мг/мл
LC50, мг/мл
LC50, мг/мл
LC50
LC50
Медианы LС50 препаратов
LC50, мг/мл
LC50, мг/мл
LC50, мг/мл
LC50
LC50
Слайд 44
PMC16
25Mg2+,43Ca2+
PMC16
PMC16
25Mg2+,43Ca2+
25Mg2+, 43Ca2+
КК, αФГК, ПК, АТФ-синтаза
∆[дНТФ]↑
Поддержка Анаболизма
Перинуклеальные Пермеазы
25Mg2+, 43Ca2+
Пересыщение
ядерного
PMC16
25Mg2+,43Ca2+
PMC16
PMC16
25Mg2+,43Ca2+
25Mg2+, 43Ca2+
КК, αФГК, ПК, АТФ-синтаза
∆[дНТФ]↑
Поддержка Анаболизма
Перинуклеальные Пермеазы
25Mg2+, 43Ca2+
Пересыщение
ядерного
![PMC16 25Mg2+,43Ca2+ PMC16 PMC16 25Mg2+,43Ca2+ 25Mg2+, 43Ca2+ КК, αФГК, ПК, АТФ-синтаза ∆[дНТФ]↑](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1064917/slide-43.jpg)
 Орган зрения
Орган зрения Презентация на тему Викторина «Карнавал животных»
Презентация на тему Викторина «Карнавал животных»  Птица года 2021 года. Кобчик
Птица года 2021 года. Кобчик Лимфатическая система человека
Лимфатическая система человека Патофизиология клетки
Патофизиология клетки Химический состав пищи (урок № 8)
Химический состав пищи (урок № 8) Молекулярный диетолог
Молекулярный диетолог Биологические методы борьбы с вредителями
Биологические методы борьбы с вредителями ЭПС, рибосомы
ЭПС, рибосомы Основные этапы антропогенеза
Основные этапы антропогенеза Аконит
Аконит Кто такие земноводные
Кто такие земноводные Экосистема Мергелевая гряда
Экосистема Мергелевая гряда Рост и развитие растений
Рост и развитие растений Чим звірі відрізняються від інших тварин (1 клас)
Чим звірі відрізняються від інших тварин (1 клас) Органы чувств
Органы чувств Функциональная организация коры больших полушарий (поля коры по К. Бродману)
Функциональная организация коры больших полушарий (поля коры по К. Бродману) Презентация на тему НЕРВНАЯ СИСТЕМА ЧЕЛОВЕКА
Презентация на тему НЕРВНАЯ СИСТЕМА ЧЕЛОВЕКА  Зрение – ведущая модальность человека
Зрение – ведущая модальность человека Вирусы. Классификация
Вирусы. Классификация Хищные растений
Хищные растений Природные сообщества леса
Природные сообщества леса Путешествие по книге. Наследство индейцев. Часть 3
Путешествие по книге. Наследство индейцев. Часть 3 Комнатные растения и уход за ними
Комнатные растения и уход за ними Механизм дыхания. Жизненная ёмкость лёгких
Механизм дыхания. Жизненная ёмкость лёгких Пищеварение полостное, внутриклеточное
Пищеварение полостное, внутриклеточное Рыбы. 7
Рыбы. 7 Растительный и животный мир Крыма
Растительный и животный мир Крыма