- Механизм передачи информации в синапсах

Содержание
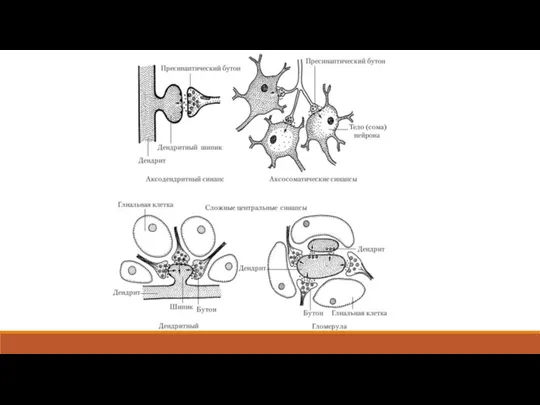
- 2. 5.1. Две разновидности синапсов Синапсом (от греч. синапсис – соединение) называют область функционального соединения одного нейрона
- 3. Если считать их со скоростью 1000 штук в секунду, то лишь через несколько тысяч лет можно
- 4. Через открывшиеся каналы проходят ионы и этот ионный ток изменяет значение мембранного потенциала покоя постсинаптической клетки.
- 5. 5.2. Передача возбуждения в нервно-мышечном синапсе Из всех существующих в организме человека синапсов наиболее простым является
- 6. Каждая веточка аксона на конце утолщается: это утолщение называют концевой пуговкой или синаптической бляшкой. В ней
- 7. Концевой пластинкой называется часть мышечной мембраны, контактирующая с нервными окончаниями. У неё складчатая поверхность, причём складки
- 8. Они состоят из пяти субъединиц, сгруппированных вокруг центральной поры. Две субъединицы из пяти одинаковы, они имеют
- 9. Когда рецепторы свяжут две молекулы ацетилхолина, конформация белковой молекулы изменяется и во всех субъединицах сдвигаются заряды
- 10. Суммарный результат ионного тока натрия и калия через хемозависимые каналы определяется преобладанием тока натрия, что приводит
- 11. Таким образом, основными этапами передачи возбуждения в нервно-мышечном синапсе являются: 1) возбуждение мотонейрона, распространение потенциала действия
- 14. 5.3. Помехи в синаптической передаче Нарушение любого из этапов синаптической передачи разлаживает деятельность синапса в целом.
- 15. Связь яда кураре с холинорецепторами обратима: если ацетилхолин накапливается в синапсе в высокой концентрации, он начинает
- 16. 5.4. Передача возбуждения в центральных синапсах Центральные синапсы, в отличие от нервно-мышечного, образованы тысячами соединений между
- 19. В возбуждающих синапсах центральной нервной системы используются, так же, как и в нервно-мышечном синапсе, хемозависимые каналы,
- 21. При таких обстоятельствах критическая деполяризация триггерной зоны возможна лишь в результате пространственной или последовательной суммации одиночных
- 22. 5.5. Постсинаптическое и пресинаптическое торможение Лишь недавно удалось найти некоторые, очень небольшие морфологические различия между возбуждающими
- 24. В том случае, когда в тормозном синапсе используются хемозависимые каналы для калия, тоже происходит гиперполяризация, так
- 25. Различие с тотальным постсинаптическим торможением заключается в том, что пресинаптическое торможение является избирательным – оно блокирует
- 26. 5.6. Функциональное значение и разновидности торможения в ЦНС Передаваясь от одного нейрона к другому, возбуждение, если
- 27. Тормозные нейроны есть во всех областях мозга, например, в спинном мозгу распространены тормозные клетки Реншоу, в
- 29. Реципрокное торможение (от лат. reciprocus – взаимный) наблюдается, например, в тех случаях, когда коллатерали входящего в
- 30. 5.7. Функциональное значение химических синапсов в переносе информации Можно с уверенностью сказать, что синапсам принадлежит решающая
- 31. 5.8. Электрические синапсы Большинство известных электрических синапсов образованы большими пресинаптическими аксонами, контактирующими со сравнительно мелкими волокнами
- 32. Коннексоны представляют собой трансмембранные белки цилиндрической формы, которые образованы шестью субъединицами и в центре имеют довольно
- 33. Направление передачи сигнала в электрических синапсах обусловлено различиями входного сопротивления контактирующих клеток. Обычно большое пресинаптическое волокно
- 34. Резюме Синапсам принадлежит ключевая роль в организации информационных потоков. Химические синапсы не просто передают сигнал, но
- 35. Вопросы для самоконтроля 61. Что происходит с выделившимся из пресинаптического окончания медиатором? А. Он диффундирует через
- 36. 63. От чего непосредственно зависит величина потенциала концевой пластинки? А. От интенсивности синтеза ацетилхолина в мотонейроне;
- 37. 65. Каким транспортным механизмом медиатор проходит через синаптическую щель к постсинаптической мембране? А. Диффузия; Б. Осмос;
- 38. 67. Преимущественный ток каких ионов обусловливает формирование потенциала концевой пластинки? А. Кальция; Б. Хлора; В. Натрия;
- 39. 69. Что из перечисленного ниже характерно для потенциала концевой пластинки? А. Образуется при использовании хемозависимых каналов;
- 40. 71. Что из указанного ниже характерно для возбуждающих постсинаптических потенциалов центральных синапсов и не характерно для
- 41. 73. Каналы каких ионов могут использоваться в тормозных синапсах? А. Калия; Б. Натрия; В. Кальция; Г.
- 42. 75. Мембранный потенциал постсинаптического нейрона равен -80 мВ, а критический уровень деполяризации – -52 мВ. На
- 43. 77. Что из указанного ниже характерно для пресинаптического торможения? А. Образование ТПСП на теле постсинаптического нейрона;
- 45. Скачать презентацию

Слайд 3Если считать их со скоростью 1000 штук в секунду, то лишь через
Если считать их со скоростью 1000 штук в секунду, то лишь через

Слайд 4Через открывшиеся каналы проходят ионы и этот ионный ток изменяет значение мембранного
Через открывшиеся каналы проходят ионы и этот ионный ток изменяет значение мембранного

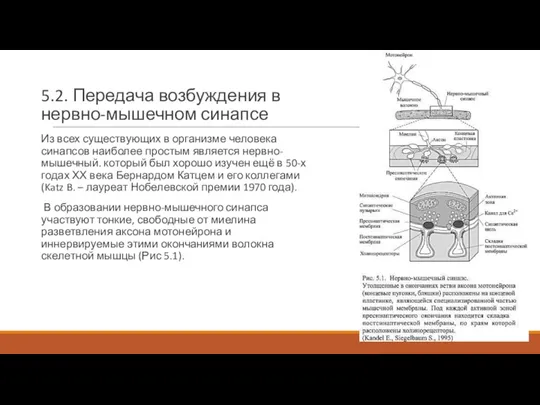
Слайд 55.2. Передача возбуждения в нервно-мышечном синапсе
Из всех существующих в организме человека
5.2. Передача возбуждения в нервно-мышечном синапсе
Из всех существующих в организме человека
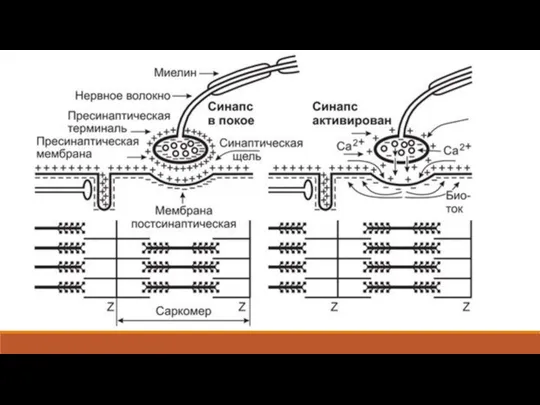
Слайд 6Каждая веточка аксона на конце утолщается: это утолщение называют концевой пуговкой или
Каждая веточка аксона на конце утолщается: это утолщение называют концевой пуговкой или

Слайд 7Концевой пластинкой называется часть мышечной мембраны, контактирующая с нервными окончаниями. У неё
Концевой пластинкой называется часть мышечной мембраны, контактирующая с нервными окончаниями. У неё

Слайд 8Они состоят из пяти субъединиц, сгруппированных вокруг центральной поры.
Две субъединицы из
Они состоят из пяти субъединиц, сгруппированных вокруг центральной поры.
Две субъединицы из

Слайд 9Когда рецепторы свяжут две молекулы ацетилхолина, конформация белковой молекулы изменяется и во
Когда рецепторы свяжут две молекулы ацетилхолина, конформация белковой молекулы изменяется и во

Слайд 10Суммарный результат ионного тока натрия и калия через хемозависимые каналы определяется преобладанием
Суммарный результат ионного тока натрия и калия через хемозависимые каналы определяется преобладанием

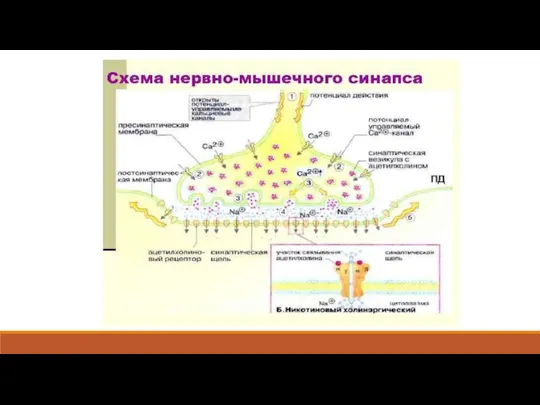
Слайд 11Таким образом, основными этапами передачи возбуждения в нервно-мышечном синапсе являются:
1) возбуждение
Таким образом, основными этапами передачи возбуждения в нервно-мышечном синапсе являются:
1) возбуждение

Слайд 145.3. Помехи в синаптической передаче
Нарушение любого из этапов синаптической передачи разлаживает
5.3. Помехи в синаптической передаче
Нарушение любого из этапов синаптической передачи разлаживает

Слайд 15Связь яда кураре с холинорецепторами обратима: если ацетилхолин накапливается в синапсе в
Связь яда кураре с холинорецепторами обратима: если ацетилхолин накапливается в синапсе в

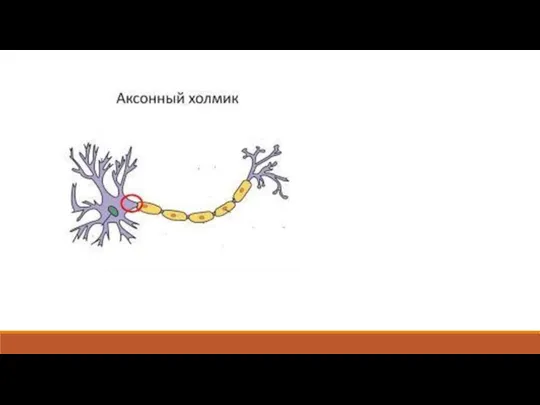
Слайд 165.4. Передача возбуждения в центральных синапсах
Центральные синапсы, в отличие от нервно-мышечного,
5.4. Передача возбуждения в центральных синапсах
Центральные синапсы, в отличие от нервно-мышечного,

Слайд 19В возбуждающих синапсах центральной нервной системы используются, так же, как и в
В возбуждающих синапсах центральной нервной системы используются, так же, как и в


Слайд 21При таких обстоятельствах критическая деполяризация триггерной зоны возможна лишь в результате пространственной
При таких обстоятельствах критическая деполяризация триггерной зоны возможна лишь в результате пространственной

Слайд 225.5. Постсинаптическое и пресинаптическое торможение
Лишь недавно удалось найти некоторые, очень небольшие
5.5. Постсинаптическое и пресинаптическое торможение
Лишь недавно удалось найти некоторые, очень небольшие


Слайд 24В том случае, когда в тормозном синапсе используются хемозависимые каналы для калия,
В том случае, когда в тормозном синапсе используются хемозависимые каналы для калия,

Слайд 25Различие с тотальным постсинаптическим торможением заключается в том, что пресинаптическое торможение является
Различие с тотальным постсинаптическим торможением заключается в том, что пресинаптическое торможение является

Слайд 265.6. Функциональное значение и разновидности торможения в ЦНС
Передаваясь от одного нейрона
5.6. Функциональное значение и разновидности торможения в ЦНС
Передаваясь от одного нейрона

Слайд 27Тормозные нейроны есть во всех областях мозга, например, в спинном мозгу распространены
Тормозные нейроны есть во всех областях мозга, например, в спинном мозгу распространены

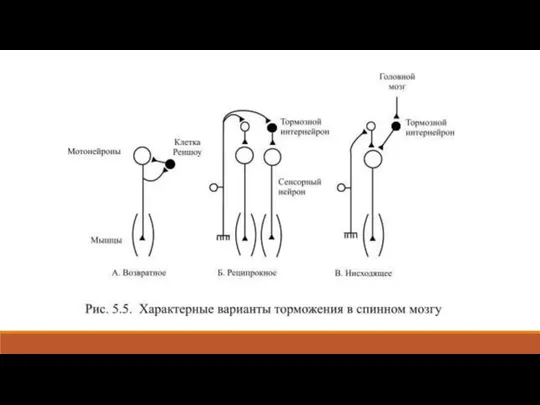
Слайд 29Реципрокное торможение (от лат. reciprocus – взаимный) наблюдается, например, в тех случаях,
Реципрокное торможение (от лат. reciprocus – взаимный) наблюдается, например, в тех случаях,

Слайд 305.7. Функциональное значение химических синапсов в переносе информации
Можно с уверенностью сказать,
5.7. Функциональное значение химических синапсов в переносе информации
Можно с уверенностью сказать,

Слайд 315.8. Электрические синапсы
Большинство известных электрических синапсов образованы большими пресинаптическими аксонами, контактирующими
5.8. Электрические синапсы
Большинство известных электрических синапсов образованы большими пресинаптическими аксонами, контактирующими

Слайд 32Коннексоны представляют собой трансмембранные белки цилиндрической формы, которые образованы шестью субъединицами и
Коннексоны представляют собой трансмембранные белки цилиндрической формы, которые образованы шестью субъединицами и

Слайд 33Направление передачи сигнала в электрических синапсах обусловлено различиями входного сопротивления контактирующих клеток.
Направление передачи сигнала в электрических синапсах обусловлено различиями входного сопротивления контактирующих клеток.

Слайд 34Резюме
Синапсам принадлежит ключевая роль в организации информационных потоков. Химические синапсы не просто
Резюме
Синапсам принадлежит ключевая роль в организации информационных потоков. Химические синапсы не просто

Слайд 35Вопросы для самоконтроля
61. Что происходит с выделившимся из пресинаптического окончания медиатором?
Вопросы для самоконтроля
61. Что происходит с выделившимся из пресинаптического окончания медиатором?

Слайд 3663. От чего непосредственно зависит величина потенциала концевой пластинки?
А. От интенсивности
63. От чего непосредственно зависит величина потенциала концевой пластинки?
А. От интенсивности

Слайд 3765. Каким транспортным механизмом медиатор проходит через синаптическую щель к постсинаптической мембране?
65. Каким транспортным механизмом медиатор проходит через синаптическую щель к постсинаптической мембране?

Слайд 3867. Преимущественный ток каких ионов обусловливает формирование потенциала концевой пластинки?
А. Кальция;
67. Преимущественный ток каких ионов обусловливает формирование потенциала концевой пластинки?
А. Кальция;

Слайд 3969. Что из перечисленного ниже характерно для потенциала концевой пластинки?
А. Образуется
69. Что из перечисленного ниже характерно для потенциала концевой пластинки?
А. Образуется

Слайд 4071. Что из указанного ниже характерно для возбуждающих постсинаптических потенциалов центральных синапсов
71. Что из указанного ниже характерно для возбуждающих постсинаптических потенциалов центральных синапсов

Слайд 4173. Каналы каких ионов могут использоваться в тормозных синапсах?
А. Калия;
Б.
73. Каналы каких ионов могут использоваться в тормозных синапсах?
А. Калия;
Б.

Слайд 4275. Мембранный потенциал постсинаптического нейрона равен -80 мВ, а критический уровень деполяризации
75. Мембранный потенциал постсинаптического нейрона равен -80 мВ, а критический уровень деполяризации

Слайд 4377. Что из указанного ниже характерно для пресинаптического торможения?
А. Образование ТПСП
77. Что из указанного ниже характерно для пресинаптического торможения?
А. Образование ТПСП

 Пептидные биорегуляторы - доказанный эффект увеличения периода эффективности человека
Пептидные биорегуляторы - доказанный эффект увеличения периода эффективности человека Дыхание. Глава 17
Дыхание. Глава 17 Времена года в Крыму
Времена года в Крыму Презентация на тему "Лимфатическая система" - презентации по Биологии
Презентация на тему "Лимфатическая система" - презентации по Биологии Зрительное восприятие изображений
Зрительное восприятие изображений Интерактивная игра-викторина Мир цветов
Интерактивная игра-викторина Мир цветов Стандартизация титрованных растворов. Интенсификация - процесс – растворение
Стандартизация титрованных растворов. Интенсификация - процесс – растворение Размножение и размножение млекопитающих
Размножение и размножение млекопитающих Размножение, рост и развитие организмов. Цветковые растения (покрытосеменные)
Размножение, рост и развитие организмов. Цветковые растения (покрытосеменные) Вода и здоровье человека
Вода и здоровье человека Витамины
Витамины Растения Красной книги России
Растения Красной книги России Белки. Определение
Белки. Определение Фенологический календарь
Фенологический календарь Ткани 8 кл
Ткани 8 кл Насекомые вредители. Своя игра
Насекомые вредители. Своя игра Презентация на тему Дарвинизм
Презентация на тему Дарвинизм  Шушпанчик сиреневый. Краткое описание вида, ареала обитания и отслеживание роста численности
Шушпанчик сиреневый. Краткое описание вида, ареала обитания и отслеживание роста численности Сочные плоды
Сочные плоды Тип кольчатые черви
Тип кольчатые черви Элементы эмбриологии
Элементы эмбриологии Антуриум - самый многочисленный род семейства Ароидных (Araceae)
Антуриум - самый многочисленный род семейства Ароидных (Araceae) Анатомия центральной нервной системы. Конечный мозг
Анатомия центральной нервной системы. Конечный мозг Взаимосвязь между структурой, свойствами и функциями клеточной мембраны
Взаимосвязь между структурой, свойствами и функциями клеточной мембраны Пищеварительная система рыб
Пищеварительная система рыб Органы выделения
Органы выделения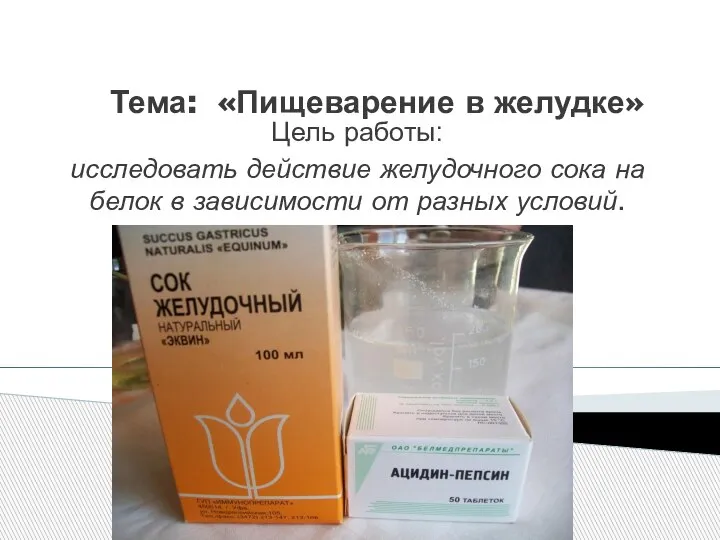 Пищеварение в желудке. Цель: исследовать действие желудочного сока на белок в зависимости от разных условий
Пищеварение в желудке. Цель: исследовать действие желудочного сока на белок в зависимости от разных условий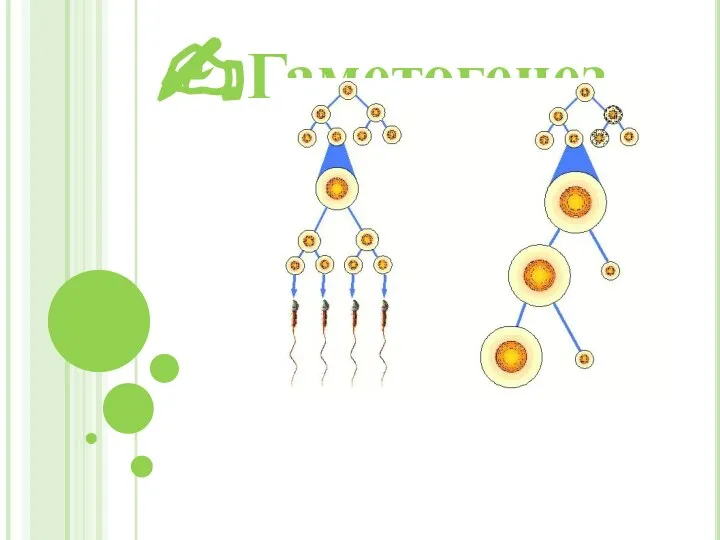 Гаметогенез
Гаметогенез