- Мейоз

Содержание
- 2. Мейоз (от греч. meiosis – уменьшение) – особый тип деления клетки, при котором происходит кратное уменьшение
- 3. Мейоз состоит из двух следующих друг за другом делений, которым предшествует однократное удвоением ДНК: редукционного (мейоз
- 4. Лептотена– начало спирализации и уплотнения хромосом, которые становятся различимыми в ядре, приобретая нитевидную форму. Зиготена. Основное
- 5. Профаза 1 3. Пахитена - основное событие этой фазы – кроссинговер (от англ. crossing-over – перекрест)
- 6. 4. Диплотена – начало отталкивания гомологичных хромосом друг от друга в составе каждого бивалента, которое начинается
- 7. Метафаза I (МI) – биваленты располагаются в плоскости экватора, образуя метафазную пластинку. В отличие от митоза,
- 8. Анафаза I (AI). В отличие от митоза центромеры не делятся. Соединенные ранее хиазмами концы гомологичных хромосом
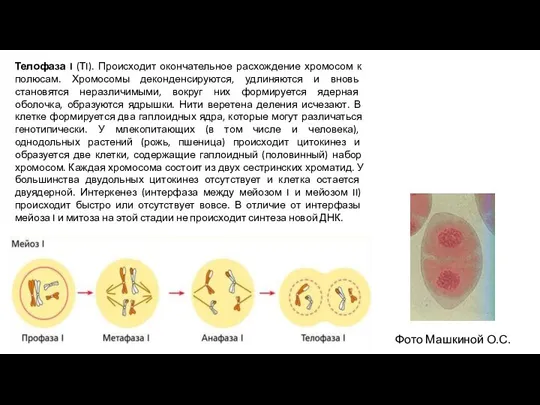
- 9. Телофаза I (ТI). Происходит окончательное расхождение хромосом к полюсам. Хромосомы деконденсируются, удлиняются и вновь становятся неразличимыми,
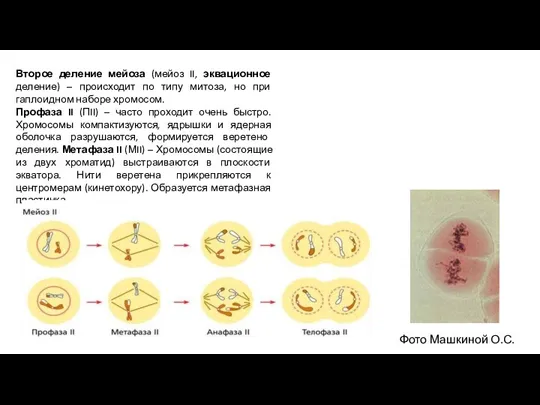
- 10. Второе деление мейоза (мейоз II, эквационное деление) – происходит по типу митоза, но при гаплоидном наборе
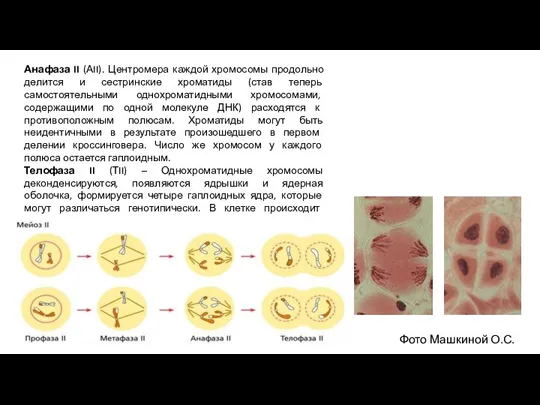
- 11. Анафаза II (АII). Центромера каждой хромосомы продольно делится и сестринские хроматиды (став теперь самостоятельными однохроматидными хромосомами,
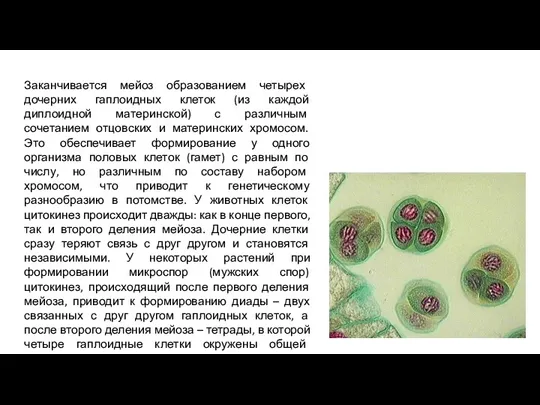
- 12. Заканчивается мейоз образованием четырех дочерних гаплоидных клеток (из каждой диплоидной материнской) с различным сочетанием отцовских и
- 13. Поддержание видового постоянства числа хромосом при половом размножении (сохранение свойственного видам кариотипа). У организмов, размножающихся половым
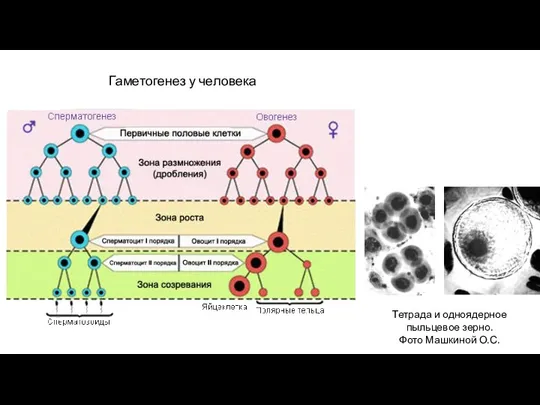
- 14. Тетрада и одноядерное пыльцевое зерно. Фото Машкиной О.С. Гаметогенез у человека
- 17. Скачать презентацию

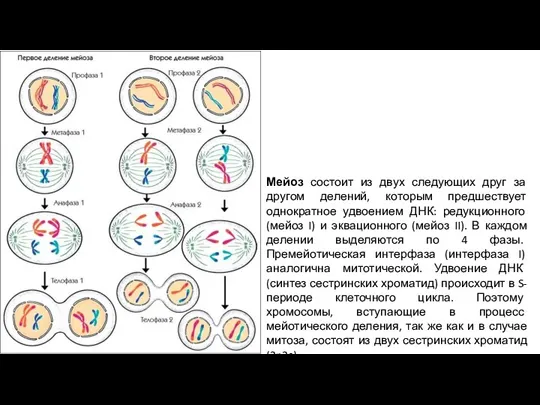
Слайд 3Мейоз состоит из двух следующих друг за другом делений, которым предшествует однократное
Мейоз состоит из двух следующих друг за другом делений, которым предшествует однократное
Слайд 4Лептотена– начало спирализации и уплотнения хромосом, которые становятся различимыми в ядре, приобретая
Лептотена– начало спирализации и уплотнения хромосом, которые становятся различимыми в ядре, приобретая

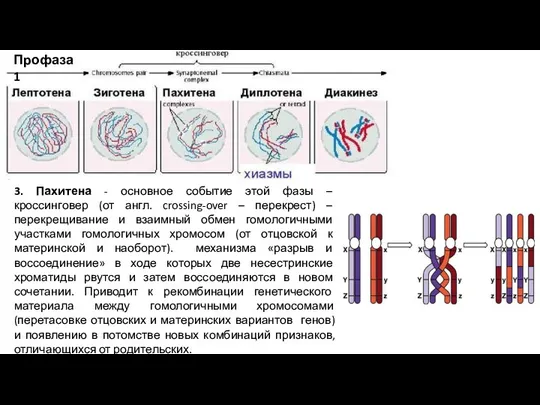
Слайд 5Профаза 1
3. Пахитена - основное событие этой фазы – кроссинговер (от
Профаза 1
3. Пахитена - основное событие этой фазы – кроссинговер (от
Слайд 64. Диплотена – начало отталкивания гомологичных хромосом друг от друга в составе
4. Диплотена – начало отталкивания гомологичных хромосом друг от друга в составе

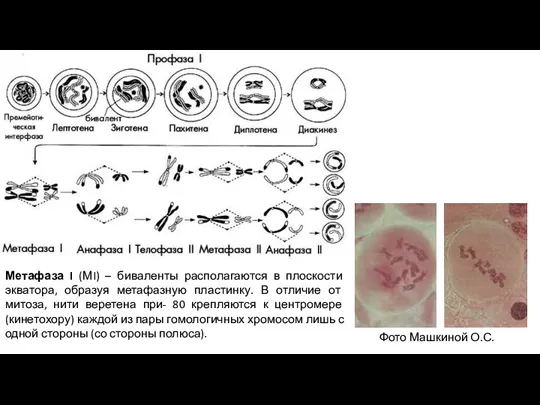
Слайд 7Метафаза I (МI) – биваленты располагаются в плоскости экватора, образуя метафазную пластинку.
Метафаза I (МI) – биваленты располагаются в плоскости экватора, образуя метафазную пластинку.
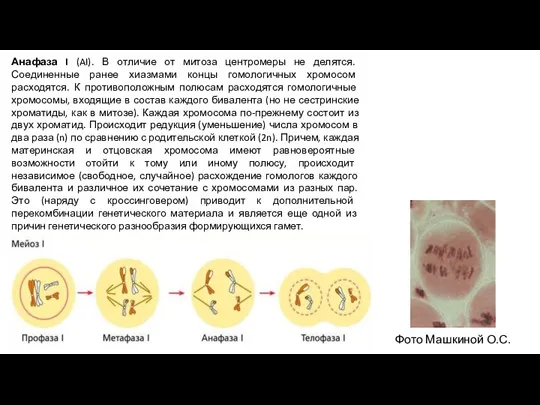
Слайд 8Анафаза I (AI). В отличие от митоза центромеры не делятся. Соединенные ранее
Анафаза I (AI). В отличие от митоза центромеры не делятся. Соединенные ранее
Слайд 9Телофаза I (ТI). Происходит окончательное расхождение хромосом к полюсам. Хромосомы деконденсируются, удлиняются
Телофаза I (ТI). Происходит окончательное расхождение хромосом к полюсам. Хромосомы деконденсируются, удлиняются
Слайд 10Второе деление мейоза (мейоз II, эквационное деление) – происходит по типу митоза,
Второе деление мейоза (мейоз II, эквационное деление) – происходит по типу митоза,
Слайд 11Анафаза II (АII). Центромера каждой хромосомы продольно делится и сестринские хроматиды (став
Анафаза II (АII). Центромера каждой хромосомы продольно делится и сестринские хроматиды (став
Слайд 12Заканчивается мейоз образованием четырех дочерних гаплоидных клеток (из каждой диплоидной материнской) с
Заканчивается мейоз образованием четырех дочерних гаплоидных клеток (из каждой диплоидной материнской) с
Слайд 13Поддержание видового постоянства числа хромосом при половом размножении (сохранение свойственного видам кариотипа).

Слайд 14Тетрада и одноядерное
пыльцевое зерно.
Фото Машкиной О.С.
Гаметогенез у человека
Тетрада и одноядерное
пыльцевое зерно.
Фото Машкиной О.С.
Гаметогенез у человека

 Основы генетики. Закономерности, установленные Г. Менделем
Основы генетики. Закономерности, установленные Г. Менделем Свойства биосферы
Свойства биосферы Дыхание
Дыхание Презентация на тему Мхи и папоротники (5 класс)
Презентация на тему Мхи и папоротники (5 класс)  ДНҚ РНҚ және хромосомадар. Олардың ағза үшін маңызы
ДНҚ РНҚ және хромосомадар. Олардың ағза үшін маңызы Я и мой питомец. Фотовыставка
Я и мой питомец. Фотовыставка Отдел Моховидные
Отдел Моховидные Белки и их свойства
Белки и их свойства Обмен веществ и превращение энергии
Обмен веществ и превращение энергии Výživa rodičů má významný vliv na funkci imunitního systému jejich potomků
Výživa rodičů má významný vliv na funkci imunitního systému jejich potomků Aqua omnia sant Вода существует везде
Aqua omnia sant Вода существует везде Рысь - родственник кошки
Рысь - родственник кошки Генетика. Почему мы похожи на своих родителей?
Генетика. Почему мы похожи на своих родителей? Развитие половых клеток. Мейоз
Развитие половых клеток. Мейоз Клиникалық диагностикалық зерттеу-биологиялық субстраттардың физико-химиялық қасиеттерін анықтайды
Клиникалық диагностикалық зерттеу-биологиялық субстраттардың физико-химиялық қасиеттерін анықтайды Мадагаскарский дневной геккон, или зелёная фельзума
Мадагаскарский дневной геккон, или зелёная фельзума Основы зоогигиены
Основы зоогигиены Функции жиров
Функции жиров Птицы зимой
Птицы зимой Химический состав живого организма. Неорганические вещества
Химический состав живого организма. Неорганические вещества Большие кошки (начало)
Большие кошки (начало) Неклеточные формы жизни. Вирусы и бактериофаги
Неклеточные формы жизни. Вирусы и бактериофаги Большая подборка по второй части. 26 задание
Большая подборка по второй части. 26 задание Дыхание и кровообращение
Дыхание и кровообращение Растительный покров земли
Растительный покров земли Опорно-двигательная система человека
Опорно-двигательная система человека Обонятельный анализатор
Обонятельный анализатор Ткан. Строение и функции. Урок 3
Ткан. Строение и функции. Урок 3