- Молекулярно-генетический уровень жизни. Матричные процессы

Содержание
- 2. Реализация генетической информации: матричные процессы
- 3. Экспрессия гена процесс, в ходе которого наследственная информацияпроцесс, в ходе которого наследственная информация от гена преобразуется
- 4. Основные отличия организации генов прокариот и эукариот
- 5. регуляторная область: рядом со структурными генами – промотор и оператор; на некотором расстоянии от оперона –
- 6. Организация генов прокариот оперон промотор – регуляторная последовательность, узнаваемая ферментом РНК-полимеразой оператор – регуляторная последовательность, связывающаяся
- 7. энхансер (англ. enhancer – усилитель, увеличитель) – небольшой участок ДНК, который после связывания с ним факторов
- 8. Организация генов прокариот промотор
- 9. более сложное строение регуляторного участка один структурный ген интронно-экзонная организация структурного гена сайленсеры – последовательность ДНК,
- 10. Организация генов эукариот - транскриптон
- 11. Транскрипция ДНК * Перенос информации на РНК Транскрибируется: короткий участок ДНК одна цепь ДНК – транскрибируемая
- 12. РНК-полимеразы
- 13. Инициация транскрипции первый этап транскрипции, в ходе которого происходит связывание РНК-полимеразы с промотором и образование первой
- 14. У прокариот холофермент РНК-полимераза непосредственно узнает определенные последовательности нуклеотидных пар в составе промотора: последовательность 5-ТАТААТ-3 (расположена
- 15. РНК-полимеразы эукариот не способны самостоятельно связываться с промоторами транскрибируемых генов. В присоединении к транскриптонам РНК-полимераз принимают
- 16. Промоторы эукариот устроены более сложно, чем прокариотические, и состоят из нескольких элементов. Из низ самым близким
- 17. Связавшись с промотором, РНК-полимераза вызывает локальную денатурацию ДНК, т. е. разделение цепей ДНК на протяжении примерно
- 18. последовательное удлинение растущей цепи РНК. перемещаясь вдоль двойной спирали ДНК, РНК-полимераза непрерывно раскручивает спираль впереди того
- 19. определяется особой нуклеотидной последовательностью ДНК, расположенной в зоне терминатора оперона. В бактериальных оперонах выделяют два типа
- 20. ρ-независимые терминаторы состоят из последовательностей, представляющих собой инвертированный повтор – палиндром, и располагаются за 16-20 нуклеотидных
- 21. Транскрипция в области палиндрома приводит к тому, что в получившемся РНК-транскрипте быстро образуется устойчивый элемент вторичной
- 22. ρ-зависимые терминаторы. Одним из факторов транскрипции прокариот является белок ρ. ρ-фактор – это имеющий четвертичную структуру
- 23. Процессинг РНК Посттранскрипционная модификация РНК совокупность процессов, которые приводят к превращению первичного транскрипта в зрелую РНК
- 24. Процессингу подвергаются: мРНК, тРНК, рРНК эукариот тРНК, рРНК прокариот мРНК прокариот синтезируются в активном виде! Процессинг
- 25. созревание сводится к разрезанию эндонуклеазами прерибосомной РНК на индивидуальные цепи, которые уже непосредственно участвуют в формировании
- 26. Процессинг РНК. рРНК Эукариоты: метилирование оснований нуклеазное расщепление – существуют четыре типа рРНК: 5S, 5,8S, 18S
- 27. Прокариоты: метилирование оснований нуклеазное расщепление – молекулы рибосомальной РНК совсем иные по своим свойствам (5S-, 16S-,
- 28. Процессинг РНК. тРНК Эукариоты: нуклеазное расщепление – удаление лидерной последовательности с 5'-конца, концевой с 3'-конца сплайсинг
- 29. Процессинг РНК. тРНК Прокариоты: нуклеазное расщепление – удаление лидерной последовательности с 5'-конца, концевой с 3'-конца замена
- 30. Процессинг РНК. тРНК
- 31. Пре-иРНК копирует всю нуклеотидную последовательность ДНК от промотора до терминатора транскриптона. То есть она включает концевые
- 32. Кэпирование – это присоединение 7-метил-ГТФ (7-метилгуанозинтрифосфат) к 5'-концу РНК, а также метилирование рибозы двух первых нуклеотидов:
- 33. Функции: защищает 5׳-конец мРНК от действия нуклеаз обеспечивает правильное расположение мРНК на малой субъединице рибосомы при
- 34. Полиаденилирование – при помощи полиаденилат-полимеразы с использованием молекул АТФ происходит присоединение к 3'-концу РНК от 100
- 35. https://youtu.be/fMnxyWvyZpY Процессинг РНК. м(и)РНК
- 36. Сплайсинг – вырезание интронов из пре-мРНК и сшивание экзонов с образованием мРНК Осуществляется сплайсосомой – комплексом
- 37. Точное разрезание границы интрон/экзон определяют три типа коротких последовательностей – сайты сплайсинга: 5’SS (донорный) exon-G-U 3’SS
- 38. Процессинг РНК. м(и)РНК В результате экзоны оказываются ковалентно соединенными обычной межнуклеотидной связью, а интрон уходит в
- 39. Процессинг РНК. м(и)РНК
- 40. https://youtu.be/vL1P7U5Bhx8 Процессинг РНК. м(и)РНК
- 41. Альтернативный сплайсинг – процесс, в результате которого первичный транскрипт может сплайсироваться разными способами и давать начало
- 42. Функциональное: поддержание белкового разнообразия Человек: ~20.000 генов, >100.000 белков Разнообразие белков у млекопитающих повысилось не за
- 43. Транс-сплайсинг особая форма процессинга РНК у эукариот, в ходе которого экзоны из двух разных первичных транскриптов
- 44. Аутосплайсинг – осущестляется без участия каких-либо белков, катализатором реакции становятся сами интроны РНК Выявлен для генов
- 45. Редактирование – это изменение нуклеотидной последовательности РНК Редактирование РНК включает: модификацию азотистых оснований, например, дезаминирование цитозина
- 46. * синтез белка на рибосомах, направляемый матрицей РНК Трансляция мРНК Стадии: активация аминокислот, или предварительный этап,
- 47. Проходит в цитоплазме Активация аминокислот – взаимодействие аминокислот с АТФ с образованием комплексов (аминоациладенилатов) под воздействием
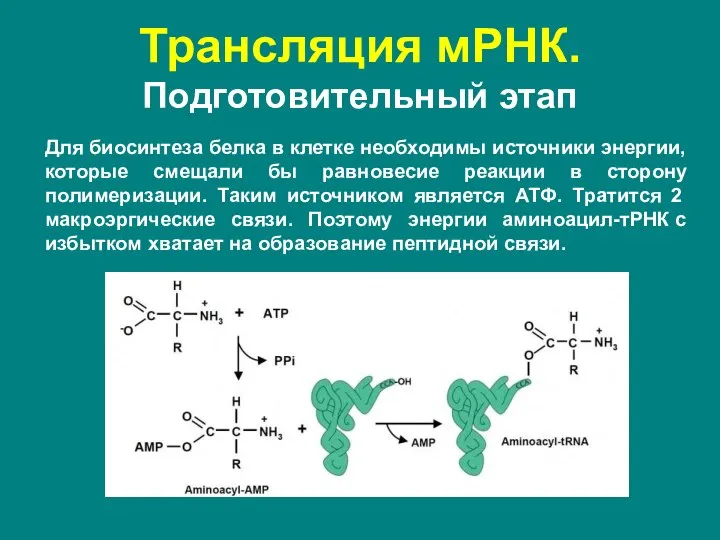
- 48. Трансляция мРНК. Подготовительный этап Для биосинтеза белка в клетке необходимы источники энергии, которые смещали бы равновесие
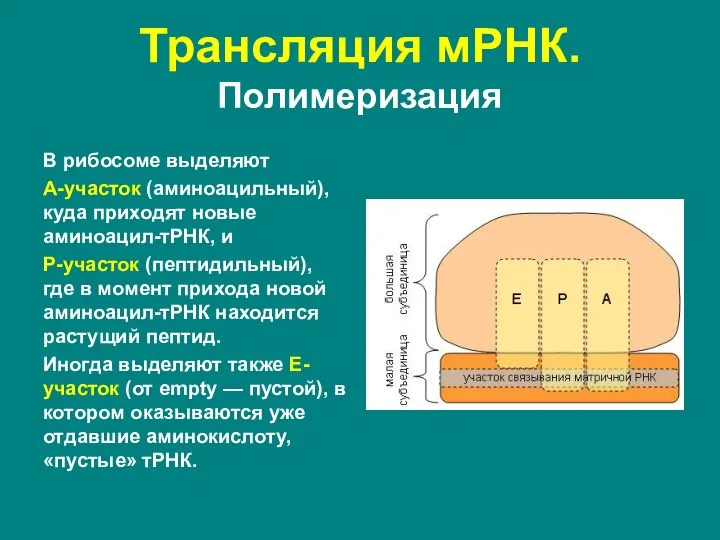
- 49. Трансляция мРНК. Полимеризация В рибосоме выделяют А-участок (аминоацильный), куда приходят новые аминоацил-тРНК, и Р-участок (пептидильный), где
- 50. Связывание малой субъединицы рибосомы с мРНК. Нахождение инициаторного, или стартового, кодона АУГ, как правило, это первый
- 51. Трансляция мРНК. Полимеризация. Инициация Механизмы инициации трансляции у про-и эукариот существенно отличаются: прокариотические рибосомы потенциально способны
- 52. Прокариоты: присоединение к МС рибосомы факторов инициации IF2-GTP (взаимодействует с тРНК), IF1 (повышает сродство МС к
- 53. Эукариоты: а) кепзависимый (сканирующий) механизм: присоединение МС рибосомы с инициирующими факторами eIF3, eIF1 и eIF2/GTP/Met-tRNAiMet к
- 54. Эукариоты: б) кепнезависимый (внутренняя инициация) механизм: присоединение МС рибосомы с инициирующими факторами на внутренний участок мРНК
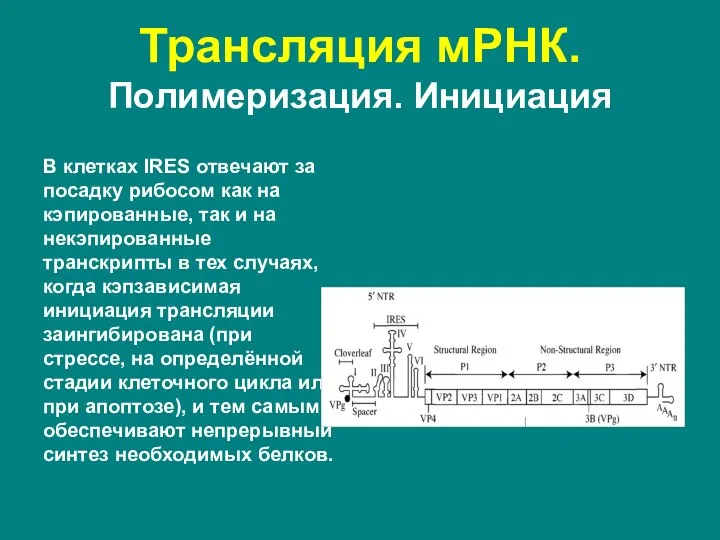
- 55. В клетках IRES отвечают за посадку рибосом как на кэпированные, так и на некэпированные транскрипты в
- 56. Трансляция мРНК. Полимеризация. Инициация
- 57. Представляет собой цикл из 3 повторяющихся событий: присоединение новой аминоацил-тРНК в А-участок в соответствии с кодоном,
- 58. Факторы элонгации: Первый фактор – EF1a у эукариот, EF-Tu – у прокариот – переносит аминоацилированную («заряженную»
- 59. Трансляция мРНК. Полимеризация. Элонгация
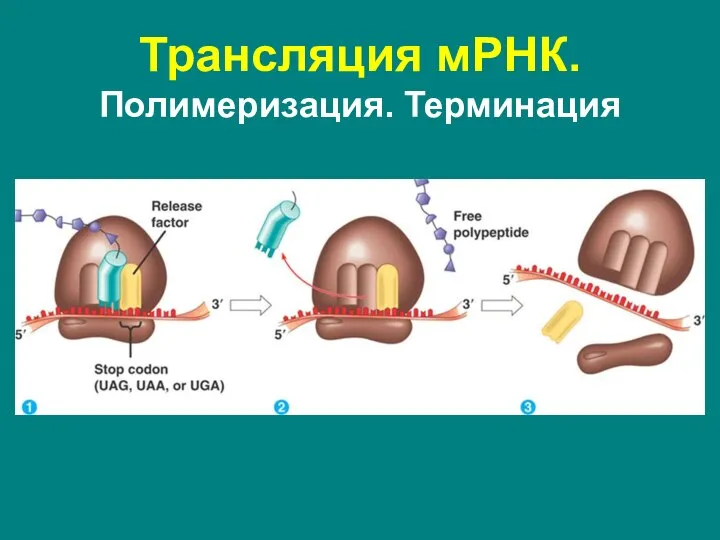
- 60. Трансляция мРНК. Полимеризация. Терминация Окончание синтеза белка, осуществляется, когда в А-сайте рибосомы оказывается один из стоп-кодонов
- 61. Трансляция мРНК. Полимеризация. Терминация
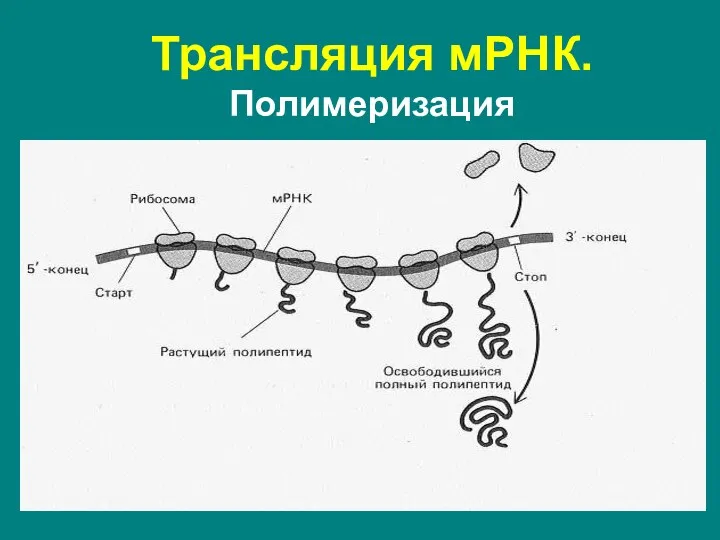
- 62. Трансляция мРНК. Полимеризация Часто на одной мРНК последовательно друг за другом синтезируют белок несколько рибосом. Это
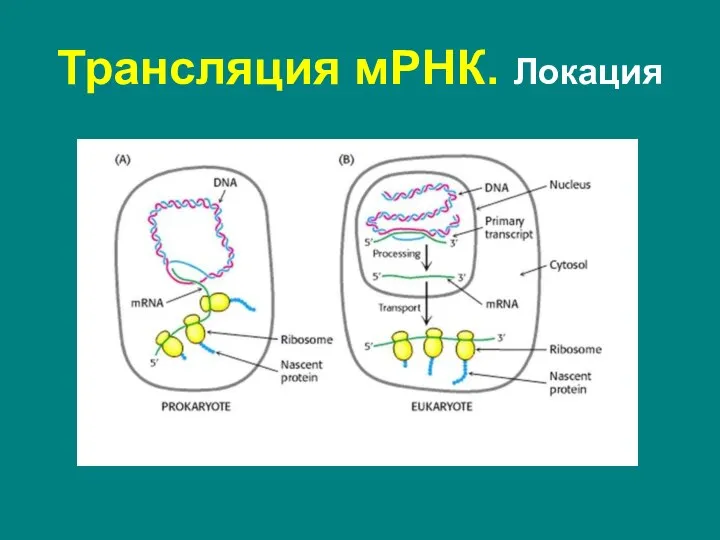
- 63. Трансляция мРНК. Локация Прокариоты: во время транскрипции в нуклеоиде или цитоплазме Эукариоты: в цитоплазме или гранулярной
- 64. Посттрансляционная модификация – это ковалентная химическая модификация белка после его синтеза на рибосоме Завершает процесс биосинтеза
- 65. К основным реакциям процессинга относятся: удаление с N-конца метионина или даже нескольких аминокислот специфичными аминопептидазами образование
- 66. присоединение химической группы к аминокислотным остаткам белковой цепи: фосфорной кислоты – к сер, тре, тир используется
- 67. включение простетической группы: углеводных остатков – гликирование требуется при синтезе гликопротеинов. гема – при синтезе гемоглобина,
- 68. Фолдинг – это процесс укладки вытянутой полипептидной цепи в правильную трехмерную пространственную структуру. Для обеспечения фолдинга
- 69. Пострансляционная модификация
- 70. Пострансляционная модификация
- 71. Пострансляционная модификация
- 73. Скачать презентацию

Слайд 3Экспрессия гена
процесс, в ходе которого наследственная информацияпроцесс, в ходе которого наследственная информация от гена преобразуется
Экспрессия гена
процесс, в ходе которого наследственная информацияпроцесс, в ходе которого наследственная информация от гена преобразуется

Слайд 4Основные отличия организации генов прокариот и эукариот
Основные отличия организации генов прокариот и эукариот

Слайд 5регуляторная область: рядом со структурными генами – промотор и оператор; на некотором
регуляторная область: рядом со структурными генами – промотор и оператор; на некотором
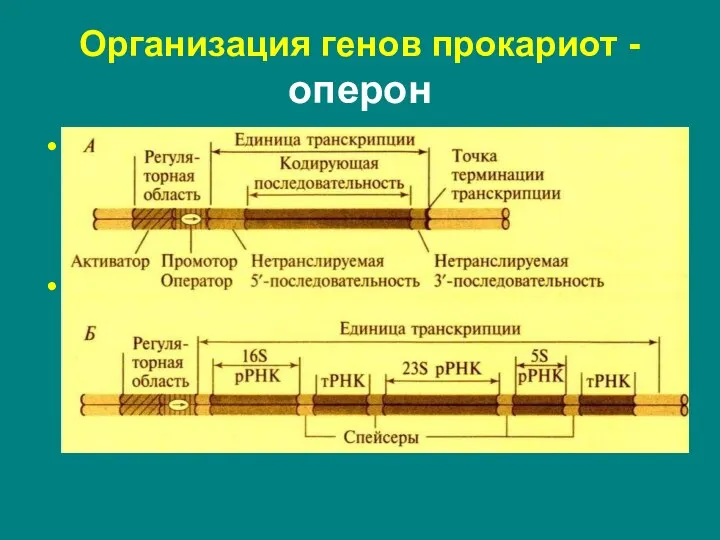
Слайд 6Организация генов прокариот оперон
промотор – регуляторная последовательность, узнаваемая ферментом РНК-полимеразой
оператор – регуляторная
Организация генов прокариот оперон
промотор – регуляторная последовательность, узнаваемая ферментом РНК-полимеразой
оператор – регуляторная

Слайд 7энхансер (англ. enhancer – усилитель, увеличитель) – небольшой участок ДНК, который после
энхансер (англ. enhancer – усилитель, увеличитель) – небольшой участок ДНК, который после

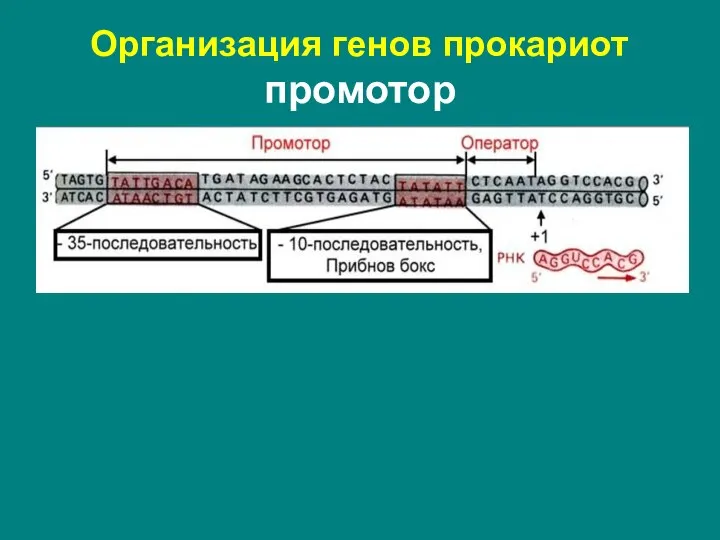
Слайд 8Организация генов прокариот промотор
Организация генов прокариот промотор
Слайд 9более сложное строение регуляторного участка
один структурный ген
интронно-экзонная организация структурного гена
сайленсеры – последовательность
более сложное строение регуляторного участка
один структурный ген
интронно-экзонная организация структурного гена
сайленсеры – последовательность

Слайд 10Организация генов эукариот - транскриптон
Организация генов эукариот - транскриптон

Слайд 11Транскрипция ДНК
* Перенос информации на РНК
Транскрибируется:
короткий участок ДНК
одна цепь ДНК
Транскрипция ДНК
* Перенос информации на РНК
Транскрибируется:
короткий участок ДНК
одна цепь ДНК

Слайд 12РНК-полимеразы
РНК-полимеразы

Слайд 13Инициация транскрипции
первый этап транскрипции, в ходе которого происходит связывание РНК-полимеразы с промотором
Инициация транскрипции
первый этап транскрипции, в ходе которого происходит связывание РНК-полимеразы с промотором

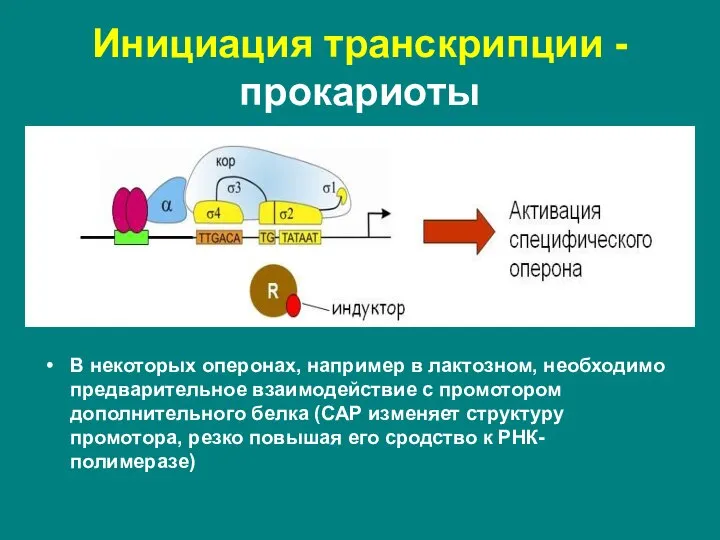
Слайд 14У прокариот холофермент РНК-полимераза непосредственно узнает определенные последовательности нуклеотидных пар в составе
У прокариот холофермент РНК-полимераза непосредственно узнает определенные последовательности нуклеотидных пар в составе
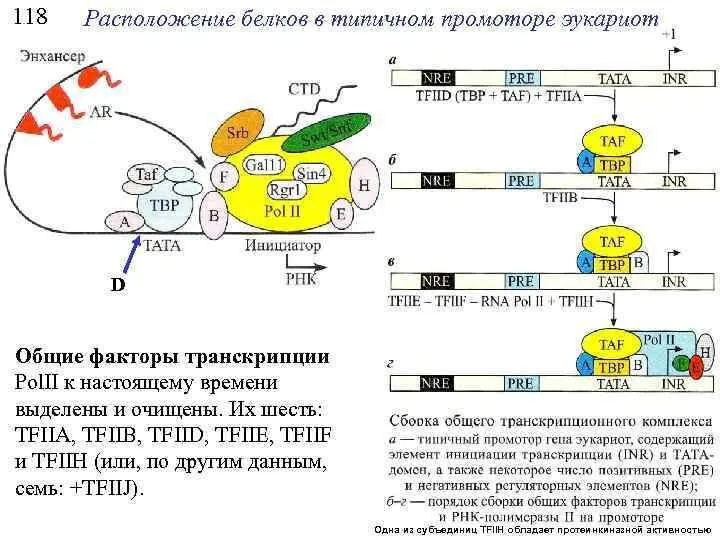
Слайд 15РНК-полимеразы эукариот не способны самостоятельно связываться с промоторами транскрибируемых генов.
В присоединении
РНК-полимеразы эукариот не способны самостоятельно связываться с промоторами транскрибируемых генов.
В присоединении
Слайд 16Промоторы эукариот устроены более сложно, чем прокариотические, и состоят из нескольких элементов.
Промоторы эукариот устроены более сложно, чем прокариотические, и состоят из нескольких элементов.

Слайд 17Связавшись с промотором, РНК-полимераза вызывает локальную денатурацию ДНК, т. е. разделение цепей
Связавшись с промотором, РНК-полимераза вызывает локальную денатурацию ДНК, т. е. разделение цепей
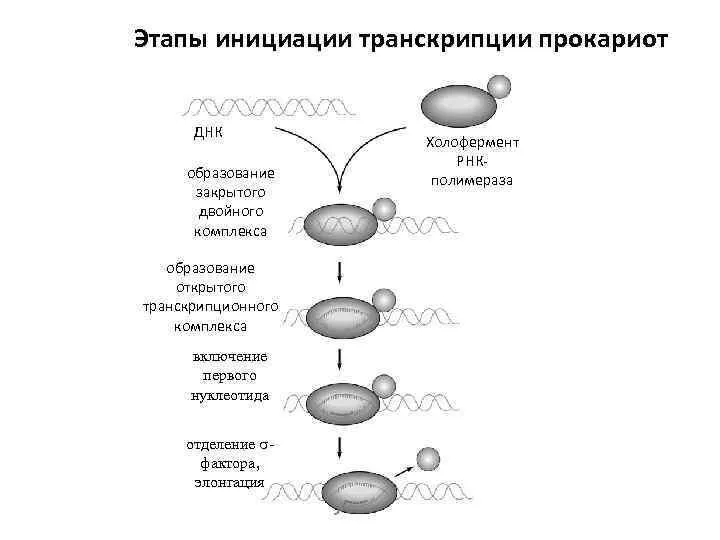
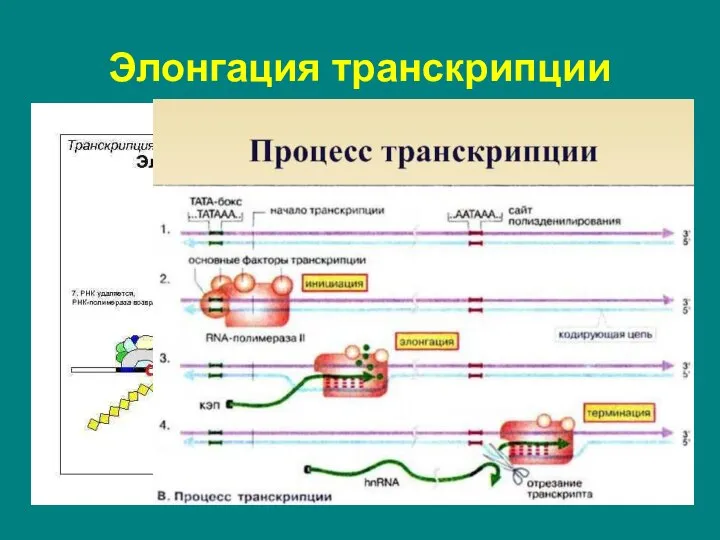
Слайд 18последовательное удлинение растущей цепи РНК.
перемещаясь вдоль двойной спирали ДНК, РНК-полимераза непрерывно раскручивает
последовательное удлинение растущей цепи РНК.
перемещаясь вдоль двойной спирали ДНК, РНК-полимераза непрерывно раскручивает
Слайд 19определяется особой нуклеотидной последовательностью ДНК, расположенной в зоне терминатора оперона.
В бактериальных оперонах
определяется особой нуклеотидной последовательностью ДНК, расположенной в зоне терминатора оперона.
В бактериальных оперонах

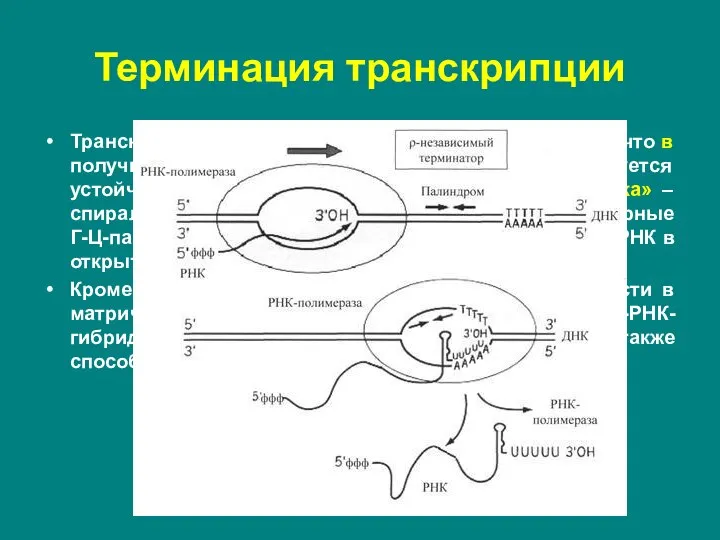
Слайд 20ρ-независимые терминаторы состоят из последовательностей, представляющих собой инвертированный повтор – палиндром, и
ρ-независимые терминаторы состоят из последовательностей, представляющих собой инвертированный повтор – палиндром, и

Слайд 21Транскрипция в области палиндрома приводит к тому, что в получившемся РНК-транскрипте быстро
Транскрипция в области палиндрома приводит к тому, что в получившемся РНК-транскрипте быстро
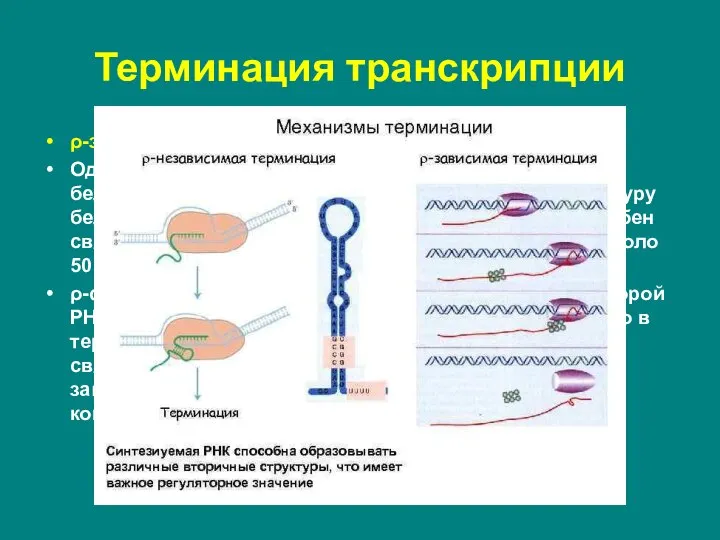
Слайд 22ρ-зависимые терминаторы.
Одним из факторов транскрипции прокариот является белок ρ. ρ-фактор –
ρ-зависимые терминаторы.
Одним из факторов транскрипции прокариот является белок ρ. ρ-фактор –
Слайд 23Процессинг РНК
Посттранскрипционная модификация РНК
совокупность процессов, которые приводят к превращению
первичного транскрипта в
зрелую РНК
Процессинг РНК
Посттранскрипционная модификация РНК
совокупность процессов, которые приводят к превращению
первичного транскрипта в
зрелую РНК

Слайд 24Процессингу подвергаются:
мРНК, тРНК, рРНК эукариот
тРНК, рРНК прокариот
мРНК прокариот синтезируются
в
Процессингу подвергаются:
мРНК, тРНК, рРНК эукариот
тРНК, рРНК прокариот
мРНК прокариот синтезируются
в

Слайд 25созревание сводится к разрезанию эндонуклеазами прерибосомной РНК на индивидуальные цепи, которые уже
созревание сводится к разрезанию эндонуклеазами прерибосомной РНК на индивидуальные цепи, которые уже

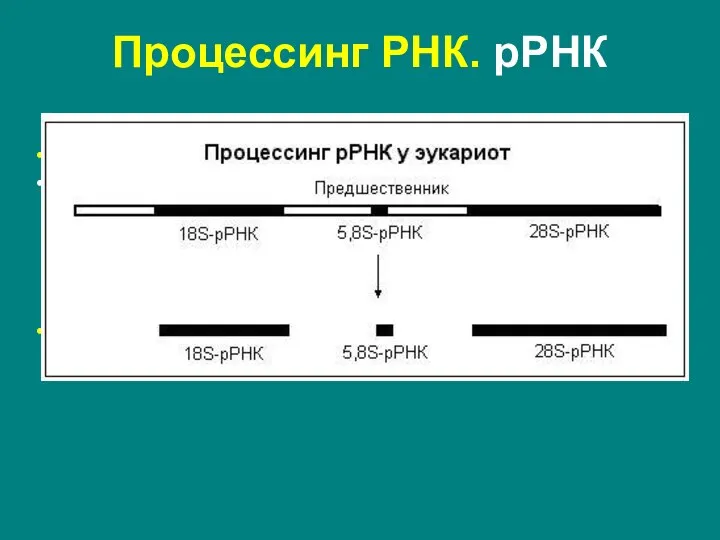
Слайд 26Процессинг РНК. рРНК
Эукариоты:
метилирование оснований
нуклеазное расщепление – существуют четыре типа рРНК: 5S,
Процессинг РНК. рРНК
Эукариоты:
метилирование оснований
нуклеазное расщепление – существуют четыре типа рРНК: 5S,
Слайд 27Прокариоты:
метилирование оснований
нуклеазное расщепление – молекулы рибосомальной РНК совсем иные по своим свойствам
Прокариоты:
метилирование оснований
нуклеазное расщепление – молекулы рибосомальной РНК совсем иные по своим свойствам

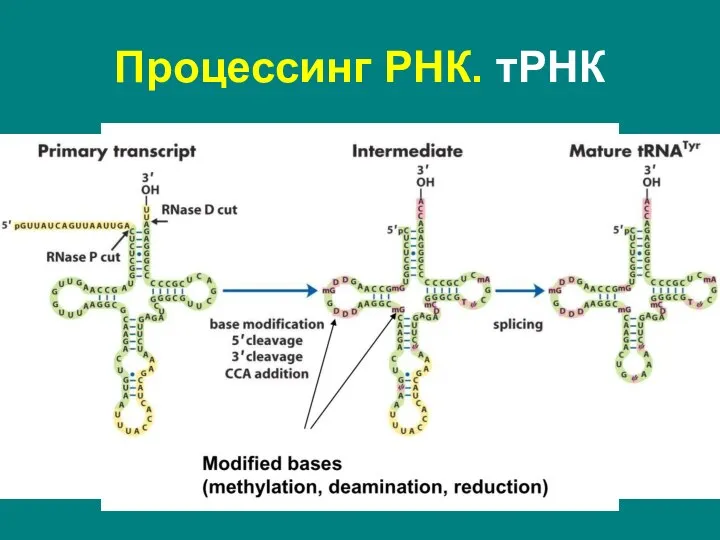
Слайд 28Процессинг РНК. тРНК
Эукариоты:
нуклеазное расщепление – удаление лидерной последовательности с 5'-конца, концевой с
Процессинг РНК. тРНК
Эукариоты:
нуклеазное расщепление – удаление лидерной последовательности с 5'-конца, концевой с

Слайд 29Процессинг РНК. тРНК
Прокариоты:
нуклеазное расщепление – удаление лидерной последовательности с 5'-конца, концевой с
Процессинг РНК. тРНК
Прокариоты:
нуклеазное расщепление – удаление лидерной последовательности с 5'-конца, концевой с

Слайд 30Процессинг РНК. тРНК
Процессинг РНК. тРНК
Слайд 31Пре-иРНК копирует всю нуклеотидную последовательность ДНК от промотора до терминатора транскриптона.
То
Пре-иРНК копирует всю нуклеотидную последовательность ДНК от промотора до терминатора транскриптона.
То

Слайд 32Кэпирование – это присоединение 7-метил-ГТФ (7-метилгуанозинтрифосфат) к 5'-концу РНК, а также метилирование
Кэпирование – это присоединение 7-метил-ГТФ (7-метилгуанозинтрифосфат) к 5'-концу РНК, а также метилирование

Слайд 33Функции:
защищает 5׳-конец мРНК от действия нуклеаз
обеспечивает правильное расположение мРНК на малой
Функции:
защищает 5׳-конец мРНК от действия нуклеаз
обеспечивает правильное расположение мРНК на малой

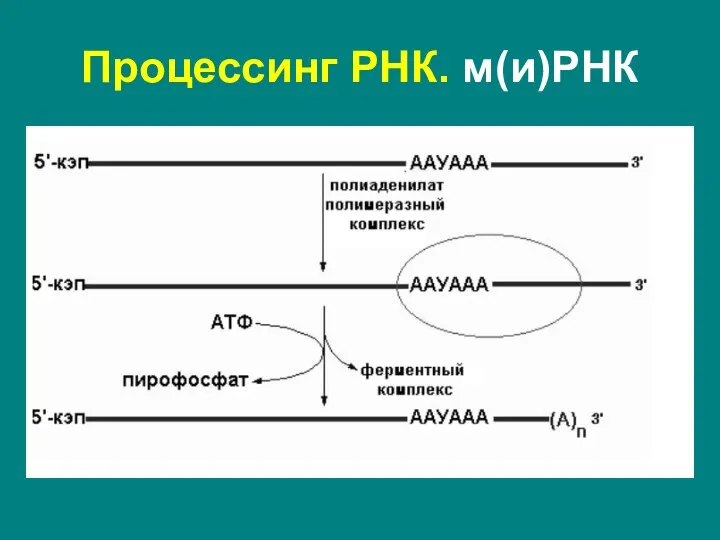
Слайд 34Полиаденилирование – при помощи полиаденилат-полимеразы с использованием молекул АТФ происходит присоединение к 3'-концу
Полиаденилирование – при помощи полиаденилат-полимеразы с использованием молекул АТФ происходит присоединение к 3'-концу
Слайд 35https://youtu.be/fMnxyWvyZpY
Процессинг РНК. м(и)РНК
https://youtu.be/fMnxyWvyZpY
Процессинг РНК. м(и)РНК

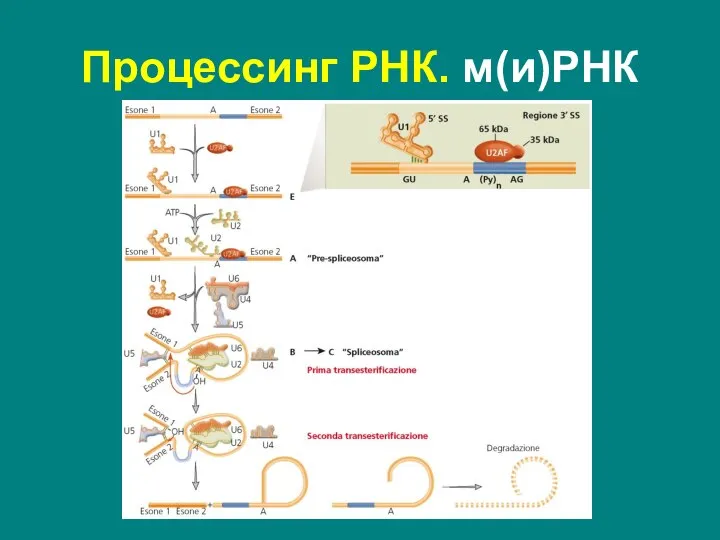
Слайд 36Сплайсинг – вырезание интронов из пре-мРНК и сшивание экзонов с образованием мРНК
Осуществляется
Сплайсинг – вырезание интронов из пре-мРНК и сшивание экзонов с образованием мРНК
Осуществляется

Слайд 37Точное разрезание границы интрон/экзон определяют три типа коротких последовательностей – сайты сплайсинга:
5’SS
Точное разрезание границы интрон/экзон определяют три типа коротких последовательностей – сайты сплайсинга:
5’SS

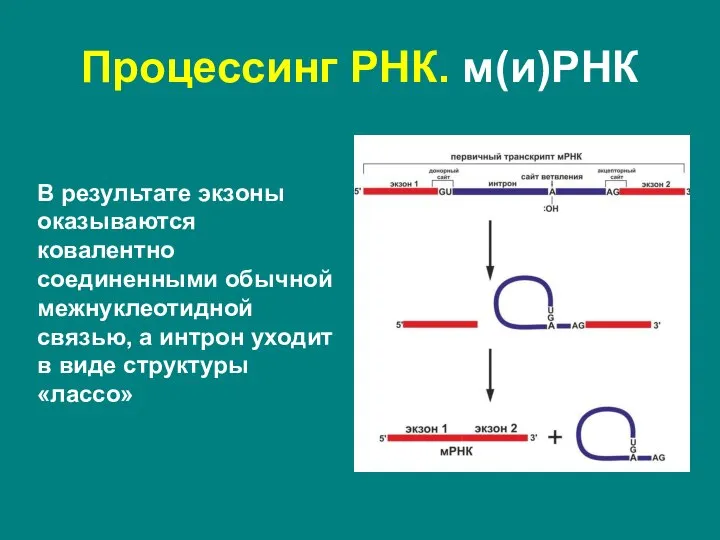
Слайд 38Процессинг РНК. м(и)РНК
В результате экзоны оказываются ковалентно соединенными обычной межнуклеотидной связью, а
Процессинг РНК. м(и)РНК
В результате экзоны оказываются ковалентно соединенными обычной межнуклеотидной связью, а
Слайд 39Процессинг РНК. м(и)РНК
Процессинг РНК. м(и)РНК
Слайд 40https://youtu.be/vL1P7U5Bhx8
Процессинг РНК. м(и)РНК
https://youtu.be/vL1P7U5Bhx8
Процессинг РНК. м(и)РНК

Слайд 41Альтернативный сплайсинг –
процесс, в результате которого первичный транскрипт может сплайсироваться разными
Альтернативный сплайсинг –
процесс, в результате которого первичный транскрипт может сплайсироваться разными
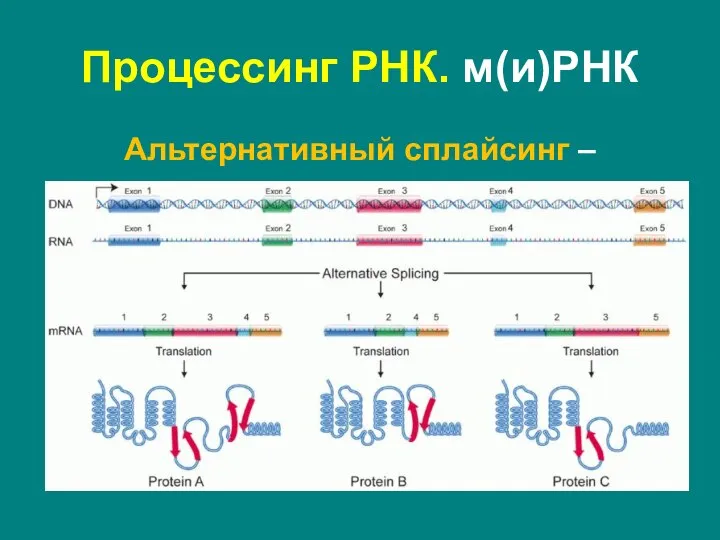
Слайд 42Функциональное:
поддержание белкового разнообразия
Человек: ~20.000 генов, >100.000 белков
Разнообразие белков у млекопитающих повысилось не
Функциональное:
поддержание белкового разнообразия
Человек: ~20.000 генов, >100.000 белков
Разнообразие белков у млекопитающих повысилось не

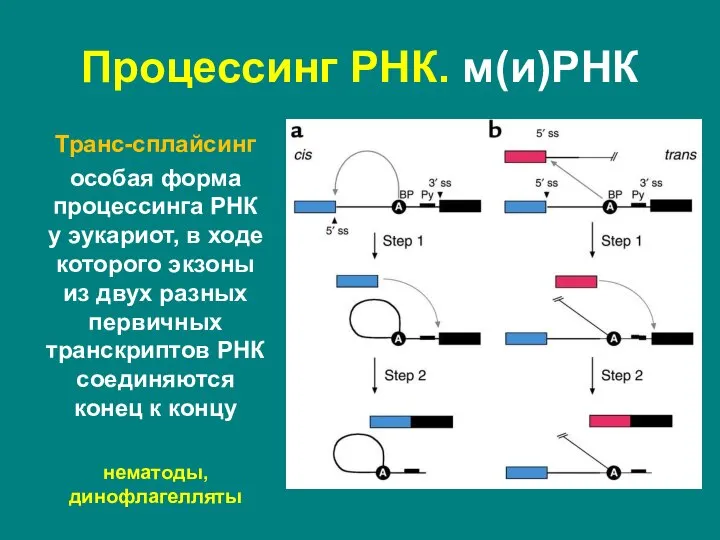
Слайд 43Транс-сплайсинг
особая форма процессинга РНК у эукариот, в ходе которого экзоны из двух
Транс-сплайсинг
особая форма процессинга РНК у эукариот, в ходе которого экзоны из двух
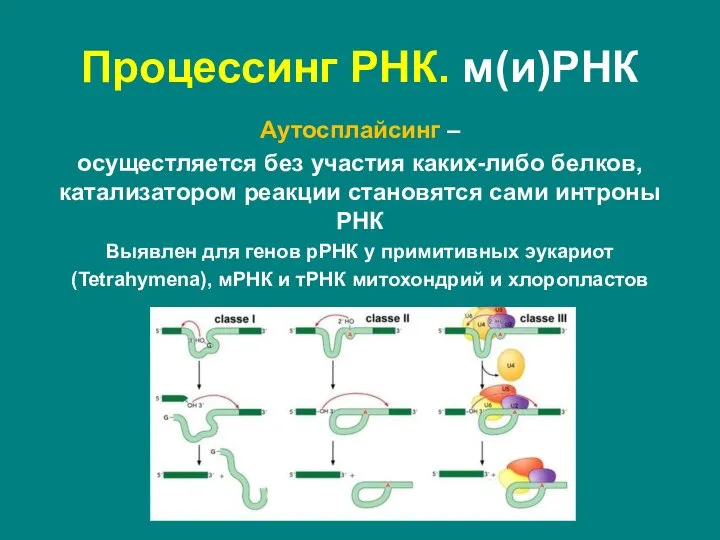
Слайд 44Аутосплайсинг –
осущестляется без участия каких-либо белков, катализатором реакции становятся сами интроны
Аутосплайсинг –
осущестляется без участия каких-либо белков, катализатором реакции становятся сами интроны
Слайд 45Редактирование – это изменение нуклеотидной последовательности РНК
Редактирование РНК включает:
модификацию азотистых оснований, например,
Редактирование – это изменение нуклеотидной последовательности РНК
Редактирование РНК включает:
модификацию азотистых оснований, например,

Слайд 46* синтез белка на рибосомах, направляемый матрицей РНК
Трансляция мРНК
Стадии:
активация аминокислот, или предварительный
* синтез белка на рибосомах, направляемый матрицей РНК
Трансляция мРНК
Стадии:
активация аминокислот, или предварительный

Слайд 47Проходит в цитоплазме
Активация аминокислот – взаимодействие аминокислот с АТФ с образованием комплексов
Проходит в цитоплазме
Активация аминокислот – взаимодействие аминокислот с АТФ с образованием комплексов

Слайд 48Трансляция мРНК. Подготовительный этап
Для биосинтеза белка в клетке необходимы источники энергии, которые
Трансляция мРНК. Подготовительный этап
Для биосинтеза белка в клетке необходимы источники энергии, которые
Слайд 49Трансляция мРНК. Полимеризация
В рибосоме выделяют
А-участок (аминоацильный), куда приходят новые аминоацил-тРНК, и
Трансляция мРНК. Полимеризация
В рибосоме выделяют
А-участок (аминоацильный), куда приходят новые аминоацил-тРНК, и
Слайд 50Связывание малой субъединицы рибосомы с мРНК.
Нахождение инициаторного, или стартового, кодона АУГ, как правило, это первый
Связывание малой субъединицы рибосомы с мРНК.
Нахождение инициаторного, или стартового, кодона АУГ, как правило, это первый

Слайд 51Трансляция мРНК. Полимеризация. Инициация
Механизмы инициации трансляции у про-и эукариот существенно отличаются:
прокариотические рибосомы потенциально способны находить
Трансляция мРНК. Полимеризация. Инициация
Механизмы инициации трансляции у про-и эукариот существенно отличаются:
прокариотические рибосомы потенциально способны находить

Слайд 52Прокариоты:
присоединение к МС рибосомы факторов инициации IF2-GTP (взаимодействует с тРНК), IF1 (повышает
Прокариоты:
присоединение к МС рибосомы факторов инициации IF2-GTP (взаимодействует с тРНК), IF1 (повышает

Слайд 53Эукариоты:
а) кепзависимый (сканирующий) механизм:
присоединение МС рибосомы с инициирующими факторами eIF3, eIF1 и
Эукариоты:
а) кепзависимый (сканирующий) механизм:
присоединение МС рибосомы с инициирующими факторами eIF3, eIF1 и

Слайд 54Эукариоты:
б) кепнезависимый (внутренняя инициация) механизм:
присоединение МС рибосомы с инициирующими факторами на внутренний
Эукариоты:
б) кепнезависимый (внутренняя инициация) механизм:
присоединение МС рибосомы с инициирующими факторами на внутренний

Слайд 55В клетках IRES отвечают за посадку рибосом как на кэпированные, так и
В клетках IRES отвечают за посадку рибосом как на кэпированные, так и
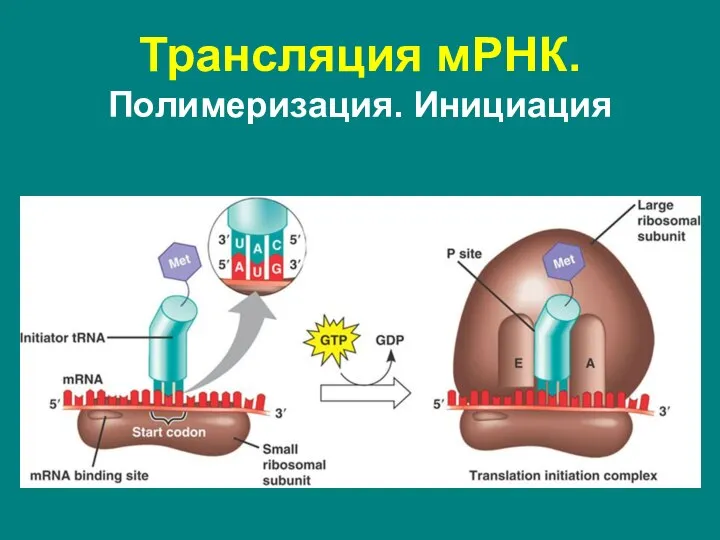
Слайд 56Трансляция мРНК. Полимеризация. Инициация
Трансляция мРНК. Полимеризация. Инициация
Слайд 57Представляет собой цикл из 3 повторяющихся событий:
присоединение новой аминоацил-тРНК в А-участок в соответствии
Представляет собой цикл из 3 повторяющихся событий:
присоединение новой аминоацил-тРНК в А-участок в соответствии

Слайд 58Факторы элонгации:
Первый фактор – EF1a у эукариот, EF-Tu – у прокариот –
Факторы элонгации:
Первый фактор – EF1a у эукариот, EF-Tu – у прокариот –

Слайд 59Трансляция мРНК. Полимеризация. Элонгация
Трансляция мРНК. Полимеризация. Элонгация
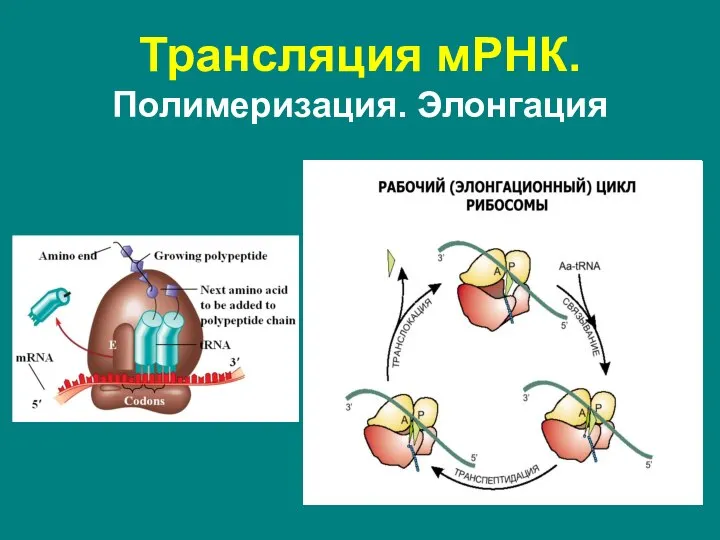
Слайд 60Трансляция мРНК. Полимеризация. Терминация
Окончание синтеза белка, осуществляется, когда в А-сайте рибосомы оказывается
Трансляция мРНК. Полимеризация. Терминация
Окончание синтеза белка, осуществляется, когда в А-сайте рибосомы оказывается

Слайд 61Трансляция мРНК. Полимеризация. Терминация
Трансляция мРНК. Полимеризация. Терминация
Слайд 62Трансляция мРНК. Полимеризация
Часто на одной мРНК последовательно друг за другом синтезируют белок
Трансляция мРНК. Полимеризация
Часто на одной мРНК последовательно друг за другом синтезируют белок
Слайд 63Трансляция мРНК. Локация
Прокариоты:
во время транскрипции в нуклеоиде или цитоплазме
Эукариоты:
в цитоплазме или гранулярной
Трансляция мРНК. Локация
Прокариоты:
во время транскрипции в нуклеоиде или цитоплазме
Эукариоты:
в цитоплазме или гранулярной
Слайд 64Посттрансляционная модификация – это ковалентная химическая модификация белка после его синтеза на рибосоме
Завершает процесс биосинтеза белка!
Увеличивает разнообразие белков в
Посттрансляционная модификация – это ковалентная химическая модификация белка после его синтеза на рибосоме
Завершает процесс биосинтеза белка!
Увеличивает разнообразие белков в

Слайд 65К основным реакциям процессинга относятся:
удаление с N-конца метионина или даже нескольких аминокислот
К основным реакциям процессинга относятся:
удаление с N-конца метионина или даже нескольких аминокислот

Слайд 66присоединение химической группы к аминокислотным остаткам белковой цепи:
фосфорной кислоты – к сер,
присоединение химической группы к аминокислотным остаткам белковой цепи:
фосфорной кислоты – к сер,

Слайд 67включение простетической группы:
углеводных остатков – гликирование требуется при синтезе гликопротеинов.
гема – при
включение простетической группы:
углеводных остатков – гликирование требуется при синтезе гликопротеинов.
гема – при

Слайд 68Фолдинг – это процесс укладки вытянутой полипептидной цепи в правильную трехмерную пространственную
Фолдинг – это процесс укладки вытянутой полипептидной цепи в правильную трехмерную пространственную

Слайд 69Пострансляционная модификация
Пострансляционная модификация
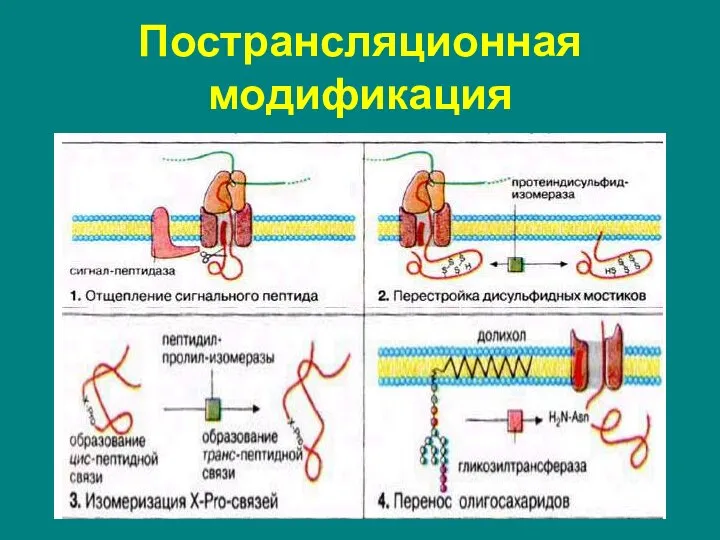
Слайд 70Пострансляционная модификация
Пострансляционная модификация
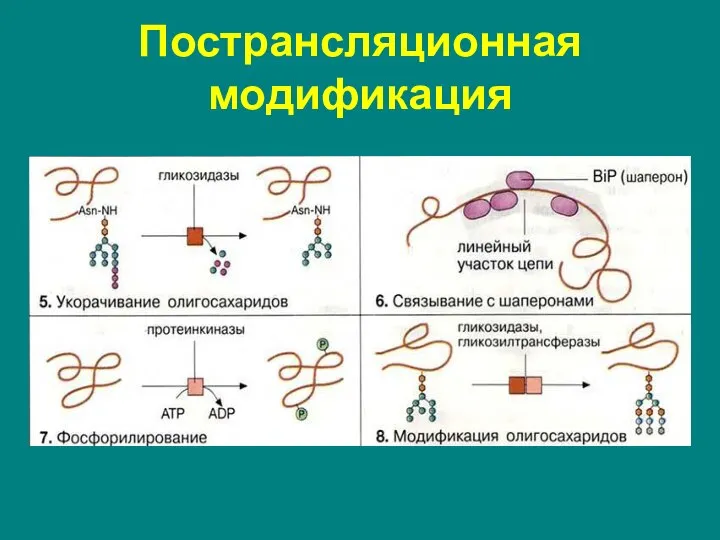
Слайд 71Пострансляционная модификация
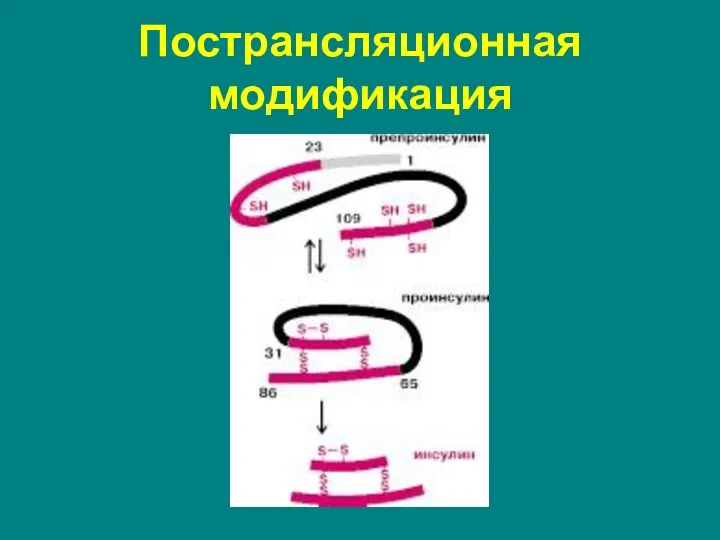
Пострансляционная модификация
 Строение и функции растительного побега
Строение и функции растительного побега Факторы антропогенеза
Факторы антропогенеза Основы физиологии равновесия
Основы физиологии равновесия Акция к дню защиты бездомных животных
Акция к дню защиты бездомных животных Клеточные и неклеточные формы жизни
Клеточные и неклеточные формы жизни Опорные конспекты (6 класс)
Опорные конспекты (6 класс) Основы цитологии. Клетка
Основы цитологии. Клетка Птицы Тамбовской области
Птицы Тамбовской области Рододендроны из дендрария
Рододендроны из дендрария Сумчатые грызуны в Австралии
Сумчатые грызуны в Австралии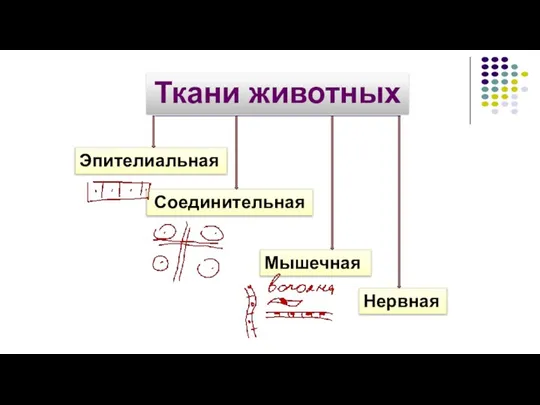 4.Ткани
4.Ткани Обработка данных секвенирования
Обработка данных секвенирования Акулы и скаты
Акулы и скаты Развитие жизни на Земле
Развитие жизни на Земле Барботирование семян
Барботирование семян Общая биология
Общая биология Рост и развитие аквариумных рыб
Рост и развитие аквариумных рыб Насекомые
Насекомые Tsns
Tsns Лабораторная работа №3 Изучение способов поглощения пищи у животных (4)
Лабораторная работа №3 Изучение способов поглощения пищи у животных (4) Генотип как целостная система. Взаимодействие генов
Генотип как целостная система. Взаимодействие генов Презентация на тему Осанка человека
Презентация на тему Осанка человека  Роль бактерий в жизни человека
Роль бактерий в жизни человека Сумчатые Млекопитающие. 7 класс
Сумчатые Млекопитающие. 7 класс Животные водоема: моллюски
Животные водоема: моллюски Кости
Кости Цветковые растения, размножаемые семенами-. (5 класс)
Цветковые растения, размножаемые семенами-. (5 класс) Типы деления клеток
Типы деления клеток