- Мышечное сокращение

Содержание
- 2. Физиологические свойства скелетных мышц: 1) возбудимость ниже, чем у нервных волокон; 2) проводимость низкая (10–13 м/с);
- 3. ВИДЫ СОКРАЩЕНИЯ: изометрическое (длина мышцы не меняется, изменяется лишь напряжение (при фиксированных концах мышцы). изотоническое (длина
- 4. Изометрическое сокращение
- 5. Изотоническое сокращение
- 6. Ауксотоническое сокращение
- 7. Классификация сокращений по выполняемой работе Концентрический тип: когда внешняя нагрузка меньше, чем развиваемое мышцей напряжение (мышца
- 8. Каждая мышца — гетерогенная популяция разных типов МВ. Тип мышцы определяют, исходя из преобладания в ней
- 9. По характеру сокращения: Фазные - осуществляют энергичные сокращения (произвольная мускулатура человека) , Тонические - специализированы на
- 10. По скорости сокращения (определяется типом миозина) Быстрые - быстрый миозин (высокая активность АТФазы), Медленные - медленный
- 11. По типу окисления: Окислительные (красные): источник АТФ - аэробное окисление глюкозы. Гликолитические (белые): источник АТФ -
- 12. На практике результаты типирования МВ комбинируют и различают три типа МВ: 1А – медленные неутомляемые (нейроны
- 13. ДВИГАТЕЛЬНЫЕ ЕДИНИЦЫ -это совокупность мышечных волокон, иннервируемых одним нейроном. Чем сложнее мышечное движение, тем меньше мышечных
- 15. РЕГУЛЯЦИЯ СИЛЫ СОКРАЩЕНИЯ число активных ДЕ частота импульсации синхронизация работы ДЕ
- 16. СТРОЕНИЕ МЫШЦЫ
- 17. САРКОМЕР Анизотропные диски (А)– миозин, Изотропные диски (I)– без миозина, только актин. Актиновые нити прикреплены к
- 18. Строение саркомера
- 19. Саркомер — часть миофибриллы, расположенная между двумя последовательными Z–дисками. В состоянии покоя и полностью растянутой мышце
- 20. Толстая миофибрилла . Каждая миозиновая нить состоит из 300–400 молекул миозина и С‑белка. Каждую миозиновую нить
- 21. СТРОЕНИЕ АКТИНА: F-актин (фибриллярный белок) – полимер G-актина, тропомиозин (фибриллярный белок), тропонин (3 субъеденицы: С –
- 22. СТРОЕНИЕ МИОЗИНА: хвост – легкий меромиозин, шейка и головка – тяжелый (скрученные в суперспираль полипептидные цепи).
- 23. ТИТИН И НЕБУЛИН Титин —3000 кДа— наподобие пружины связывает концы толстых нитей с Z-линией. Другой гигантский
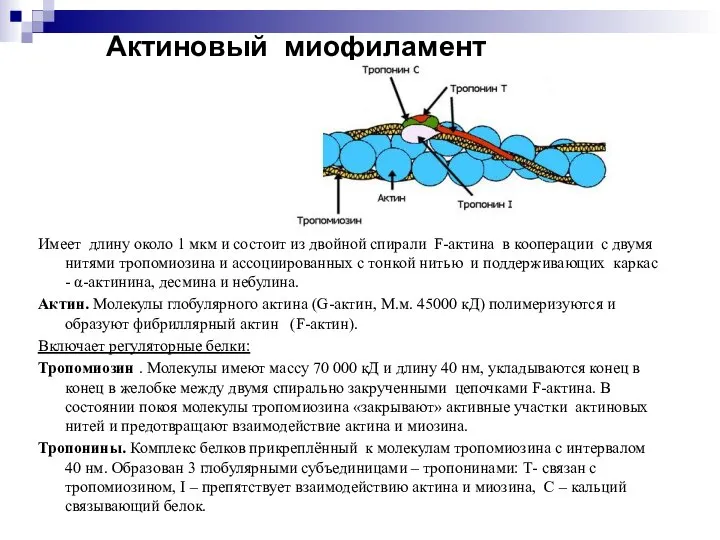
- 24. Актиновый миофиламент Имеет длину около 1 мкм и состоит из двойной спирали F-актина в кооперации с
- 25. Механизм мышечного сокращения
- 26. МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ ИСТОРИЯ: 1) Гален 2 век – живой дух, 2) линейное сокращение белка, 3)
- 27. ТЕОРИЯ СКОЛЬЖЕНИЯ НИТЕЙ предложена Хью Хаксли
- 28. Сокращение мышцы происходит в результате движения актиновой и миозиновой нити относительно друг друга (благодаря присоединению головок
- 29. 1. Ca2+ связывается с тропонином C, 2. Тропомиозин сдвигается относительно актина, 3. Открывается доступ к центрам
- 30. 5. Головки миозина изменяют свою конформацию, создавая тянущее усилие (за счёт шарнирного участка в области шейки
- 31. Схема сокращения
- 32. 7. Затем головка миозина связывается с молекулой АТФ, что приводит к отделению миозина от актина. 8.
- 34. Электро-механическое сопряжение - это процесс превращения импульса нервного волокна в сокращение мышцы
- 35. Электромеханическое сопряжение
- 36. Этапы электромеханического сопряжения Передача импульса через нервно-мышечный синапс. Возникновение ВПСП. Возникновение ПД на околосинаптической мембране. Проведение
- 37. Освобождение Са2+ из СПР и повышение его концентрации в саркоплазме на порядок (от 10-7 до 10-6
- 38. Каким образом Са2+ выделяется из СПР?
- 39. Существует несколько гипотез высвобождения Са2+ из СПР: 1. Волна деполяризации спускается по Т-трубочкам и переходит на
- 40. 2. Во внешнюю мембрану миоцита встроены потенциалзависимые и/или рецепторуправляемые Са2+-каналы. Небольшое повышение концентрации Са2+ в миоплазме
- 41. 3. Высвобождение Са2+ из СПР может индуцироваться инозитол–1,4,5–трифосфатом (ИФ3) – вторичным посредником, выделившимся из внешней мембраны
- 42. 4.Гипотеза о прямой морфофункциональной взаимосвязи между дигидропиридиновым рецептором Т-трубочек (Са2+-каналом L-типа) и рианодиновым рецептором СПР (тоже
- 43. Суммация мышечных сокращений
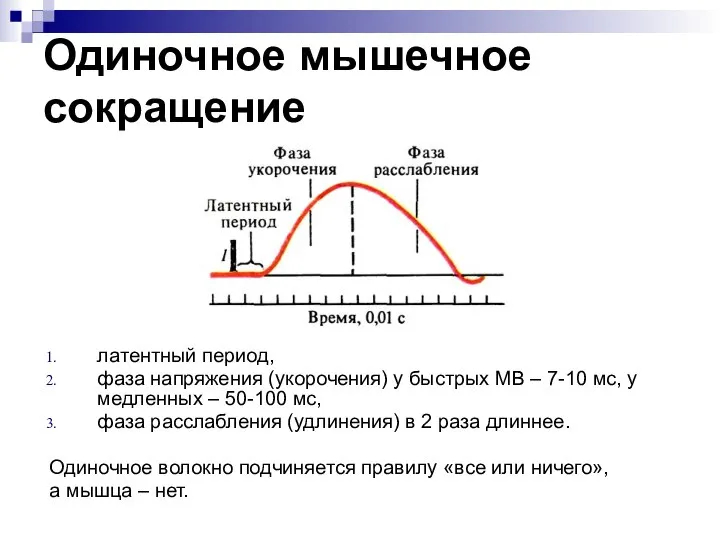
- 44. Одиночное мышечное сокращение латентный период, фаза напряжения (укорочения) у быстрых МВ – 7-10 мс, у медленных
- 45. Соотношение между длительностью ПД и сокращением мышцы
- 46. Если частота ПД больше чем длительность сокращения, то происходит явление суммации сокращений - те́танус Но последующее
- 47. гладкий тетанус возникает если последующий ПД попадает в фазу укорочения мышцы, зубчатый тетанус - если в
- 48. Теории возникновения тетануса Гельмгольц – тетанус – это механическое сложение одиночных мышечных сокращений, Введенский – тетанус
- 49. КОНТРАКТУРА - обратимое нераспространяющееся стационарное сокращение Типы контрактур: Посттетаническая (высокий остаточный Са2+) Кофеиновая – кофеин проникая
- 50. Гладкие мышцы
- 51. Физиологические особенности гладких мышц 1) нестабильный мембранный потенциал, который поддерживает мышцы в состоянии постоянного частичного сокращения
- 52. Морфологические особенности Актиновые нити прикреплены к плотным тельцам, миозиновые нити формируются только при сокращении. СПР выражен
- 53. Иннервация ГМК иннервируют симпатические (адренергические) и отчасти парасимпатические (холинергические) нервные волокна. Нейромедиаторы диффундируют из варикозных терминальных
- 54. Гуморальная регуляция В мембрану разных ГМК встроены рецепторы ацетилхолина, гистамина, атриопептина, ангиотензина, адренорецепторы и множество других.
- 55. Са+ в гладкой мышце
- 56. Холиномиметики усиливают сокращение оказывают на мышцу тот же эффект, что и ацетилхолин. Различие заключается в том,
- 57. Кураре и курареподобные препараты широко применяются в анестезиологии. Тубокурарин препятствует деполяризующему действию ацетилхолина. Дитилин приводит к
- 58. Ботулотоксин и столбнячный токсин блокируют секрецию медиатора из нервных терминалей, подавляя торможение в ЦНС. Поэтому сокращение
- 60. Скачать презентацию
Слайд 2Физиологические свойства скелетных мышц:
1) возбудимость ниже, чем у нервных волокон;
2) проводимость низкая (10–13 м/с);
3) фаза рефрактерности
Физиологические свойства скелетных мышц:
1) возбудимость ниже, чем у нервных волокон;
2) проводимость низкая (10–13 м/с);
3) фаза рефрактерности

Слайд 3ВИДЫ СОКРАЩЕНИЯ:
изометрическое (длина мышцы не меняется, изменяется лишь напряжение (при фиксированных концах
ВИДЫ СОКРАЩЕНИЯ:
изометрическое (длина мышцы не меняется, изменяется лишь напряжение (при фиксированных концах

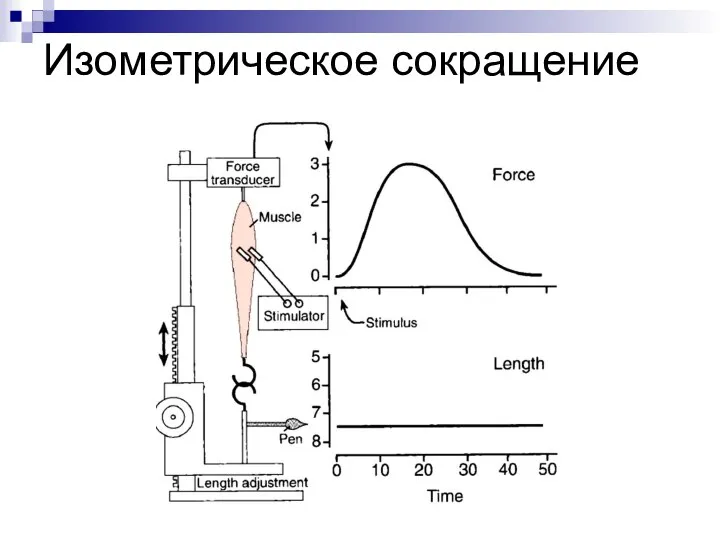
Слайд 4Изометрическое сокращение
Изометрическое сокращение
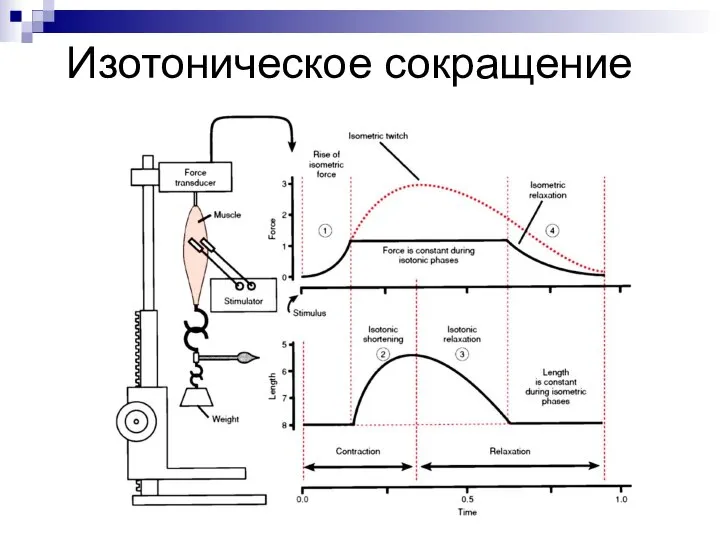
Слайд 5Изотоническое сокращение
Изотоническое сокращение
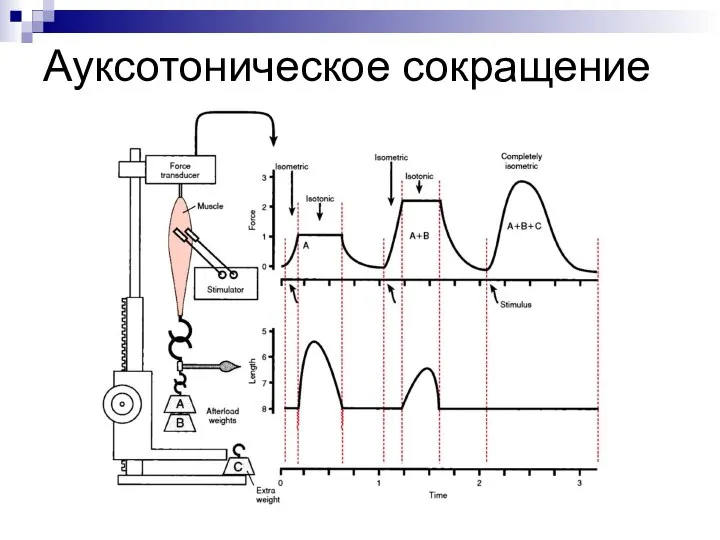
Слайд 6Ауксотоническое сокращение
Ауксотоническое сокращение
Слайд 7Классификация сокращений по выполняемой работе
Концентрический тип: когда внешняя нагрузка меньше, чем развиваемое
Классификация сокращений по выполняемой работе
Концентрический тип: когда внешняя нагрузка меньше, чем развиваемое

Слайд 8Каждая мышца — гетерогенная популяция разных типов МВ. Тип мышцы определяют, исходя из
Каждая мышца — гетерогенная популяция разных типов МВ. Тип мышцы определяют, исходя из

Слайд 9По характеру сокращения:
Фазные - осуществляют энергичные сокращения (произвольная мускулатура человека) ,
Тонические -
По характеру сокращения:
Фазные - осуществляют энергичные сокращения (произвольная мускулатура человека) ,
Тонические -

Слайд 10По скорости сокращения
(определяется типом миозина)
Быстрые - быстрый миозин (высокая активность АТФазы),
Медленные
По скорости сокращения
(определяется типом миозина)
Быстрые - быстрый миозин (высокая активность АТФазы),
Медленные

Слайд 11По типу окисления:
Окислительные (красные): источник АТФ - аэробное окисление глюкозы.
Гликолитические (белые): источник
По типу окисления:
Окислительные (красные): источник АТФ - аэробное окисление глюкозы.
Гликолитические (белые): источник

Слайд 12На практике результаты типирования МВ комбинируют и различают три типа МВ:
1А –
На практике результаты типирования МВ комбинируют и различают три типа МВ:
1А –

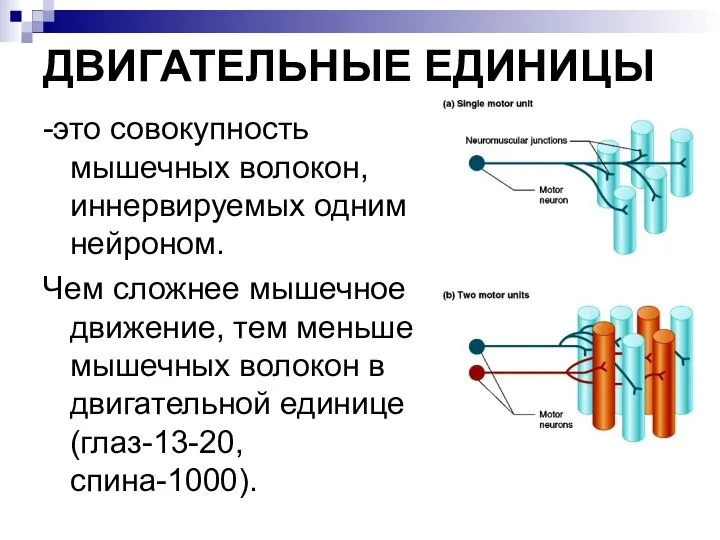
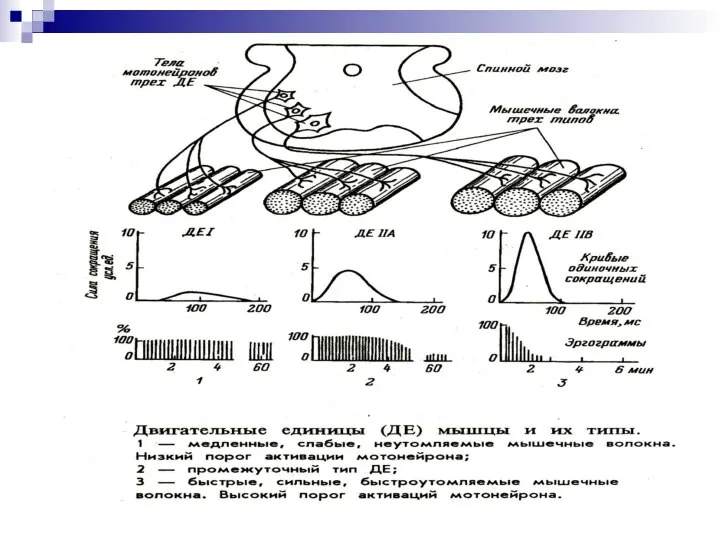
Слайд 13ДВИГАТЕЛЬНЫЕ ЕДИНИЦЫ
-это совокупность мышечных волокон, иннервируемых одним нейроном.
Чем сложнее мышечное движение, тем
ДВИГАТЕЛЬНЫЕ ЕДИНИЦЫ
-это совокупность мышечных волокон, иннервируемых одним нейроном.
Чем сложнее мышечное движение, тем
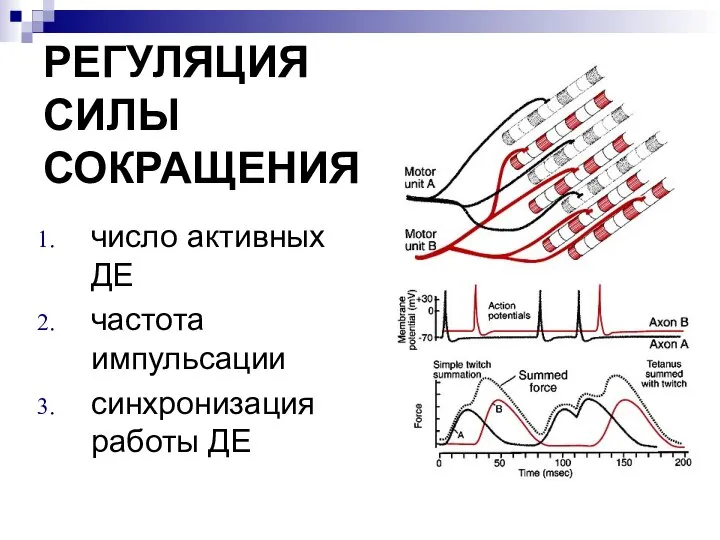
Слайд 15РЕГУЛЯЦИЯ
СИЛЫ
СОКРАЩЕНИЯ
число активных ДЕ
частота импульсации
синхронизация работы ДЕ
РЕГУЛЯЦИЯ
СИЛЫ
СОКРАЩЕНИЯ
число активных ДЕ
частота импульсации
синхронизация работы ДЕ
Слайд 16СТРОЕНИЕ
МЫШЦЫ
СТРОЕНИЕ
МЫШЦЫ
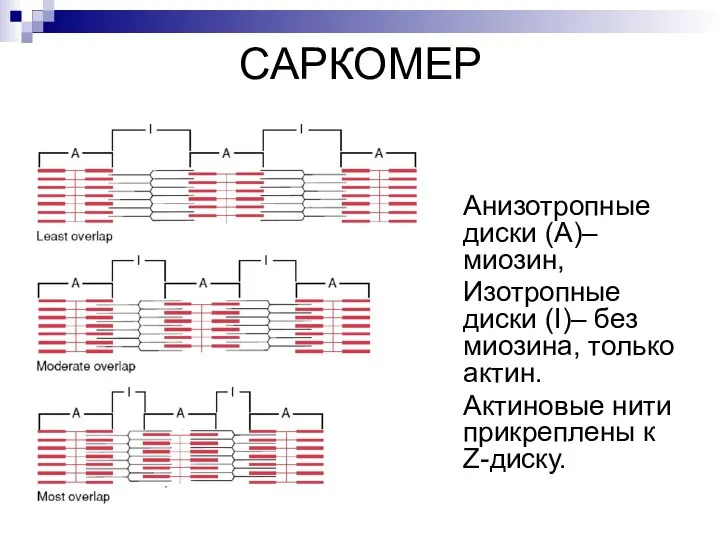
Слайд 17САРКОМЕР
Анизотропные диски (А)– миозин,
Изотропные диски (I)– без миозина, только актин.
Актиновые нити прикреплены
САРКОМЕР
Анизотропные диски (А)– миозин,
Изотропные диски (I)– без миозина, только актин.
Актиновые нити прикреплены
Слайд 18Строение саркомера
Строение саркомера
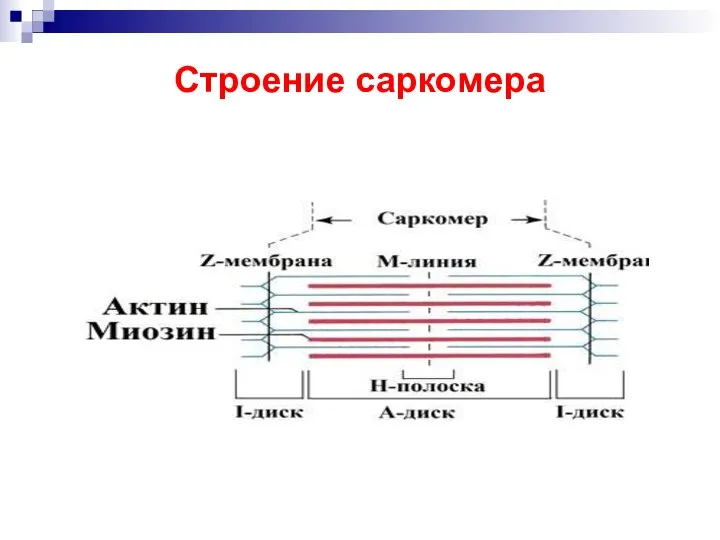
Слайд 19 Саркомер — часть миофибриллы, расположенная между двумя последовательными Z–дисками.
В состоянии
Саркомер — часть миофибриллы, расположенная между двумя последовательными Z–дисками.
В состоянии

Слайд 20Толстая миофибрилла .
Каждая миозиновая нить состоит из 300–400 молекул миозина и
Толстая миофибрилла .
Каждая миозиновая нить состоит из 300–400 молекул миозина и

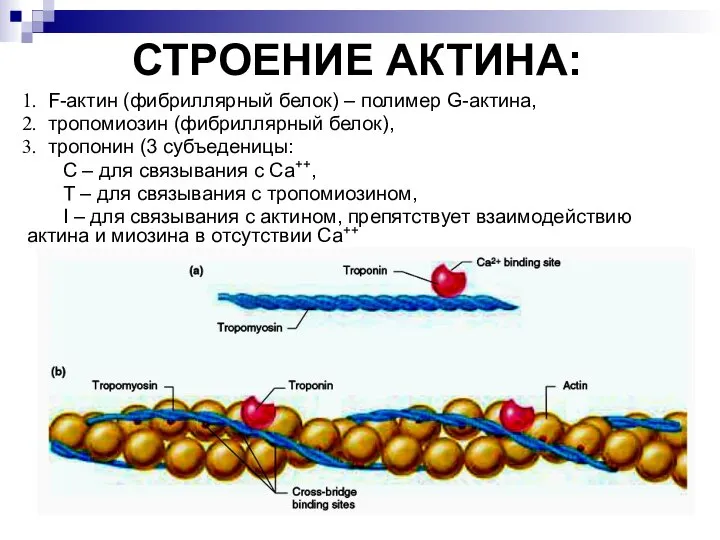
Слайд 21СТРОЕНИЕ АКТИНА:
F-актин (фибриллярный белок) – полимер G-актина,
тропомиозин (фибриллярный белок),
тропонин (3
СТРОЕНИЕ АКТИНА:
F-актин (фибриллярный белок) – полимер G-актина,
тропомиозин (фибриллярный белок),
тропонин (3
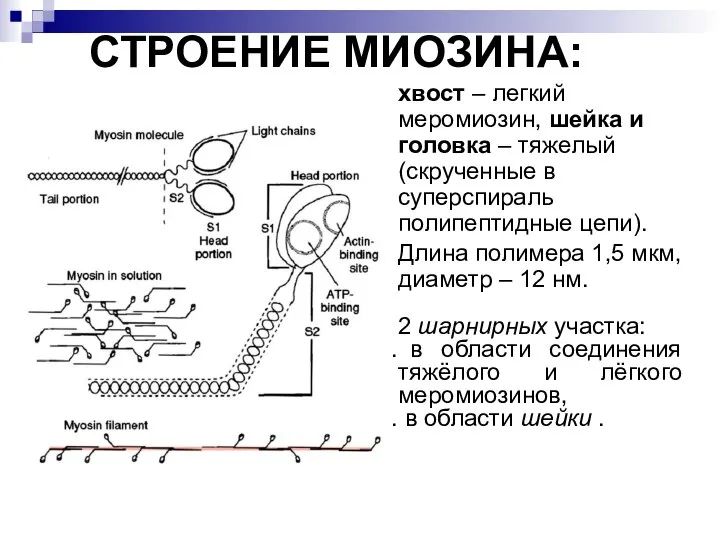
Слайд 22СТРОЕНИЕ МИОЗИНА:
хвост – легкий меромиозин, шейка и головка – тяжелый (скрученные в
СТРОЕНИЕ МИОЗИНА:
хвост – легкий меромиозин, шейка и головка – тяжелый (скрученные в
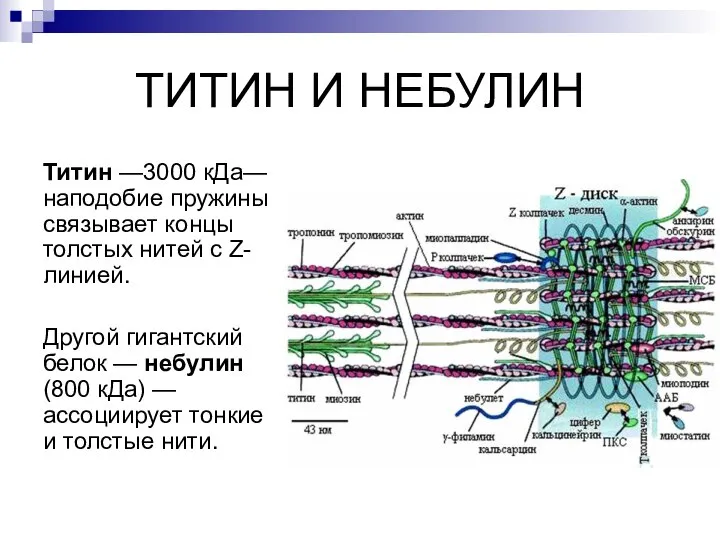
Слайд 23ТИТИН И НЕБУЛИН
Титин —3000 кДа— наподобие пружины связывает концы толстых нитей с Z-линией.
ТИТИН И НЕБУЛИН
Титин —3000 кДа— наподобие пружины связывает концы толстых нитей с Z-линией.
Слайд 24
Актиновый миофиламент
Имеет длину около 1 мкм и состоит из двойной спирали F-актина
Актиновый миофиламент
Имеет длину около 1 мкм и состоит из двойной спирали F-актина
Слайд 25Механизм мышечного сокращения
Механизм мышечного сокращения

Слайд 26МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ
ИСТОРИЯ:
1) Гален 2 век – живой дух,
2) линейное
МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ
ИСТОРИЯ:
1) Гален 2 век – живой дух,
2) линейное

Слайд 27ТЕОРИЯ СКОЛЬЖЕНИЯ НИТЕЙ
предложена Хью Хаксли
ТЕОРИЯ СКОЛЬЖЕНИЯ НИТЕЙ
предложена Хью Хаксли

Слайд 28Сокращение мышцы происходит в результате движения актиновой и миозиновой нити относительно друг
Сокращение мышцы происходит в результате движения актиновой и миозиновой нити относительно друг

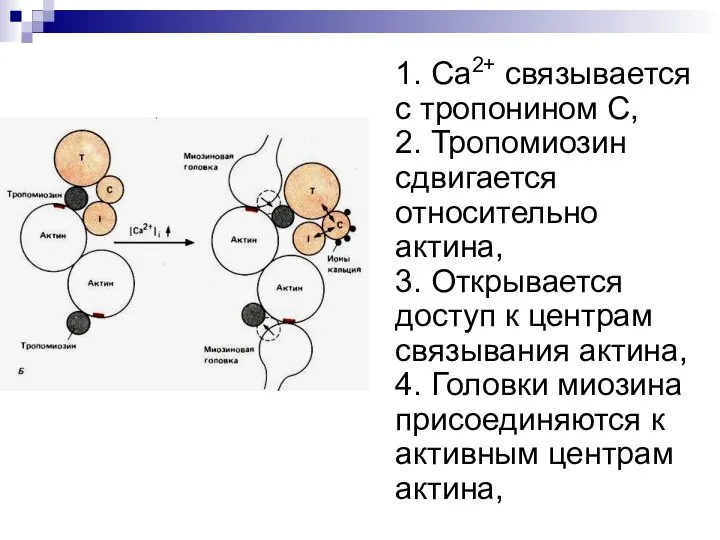
Слайд 291. Ca2+ связывается с тропонином C,
2. Тропомиозин сдвигается относительно актина,
3. Открывается доступ
1. Ca2+ связывается с тропонином C, 2. Тропомиозин сдвигается относительно актина, 3. Открывается доступ
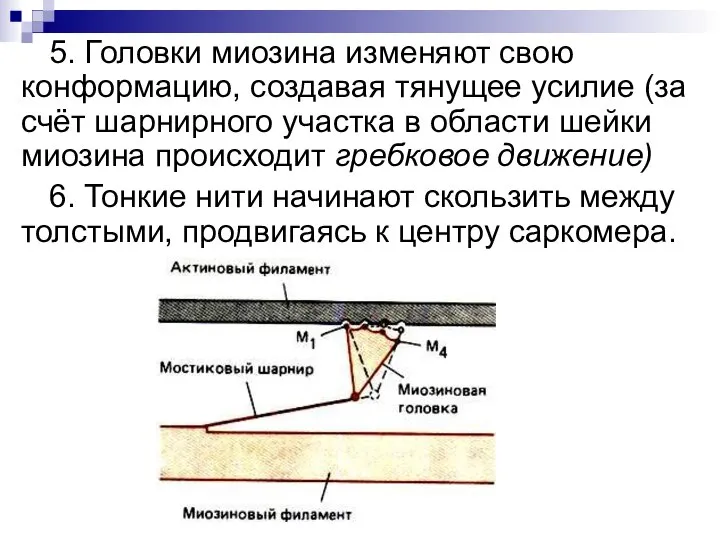
Слайд 305. Головки миозина изменяют свою конформацию, создавая тянущее усилие (за счёт шарнирного
5. Головки миозина изменяют свою конформацию, создавая тянущее усилие (за счёт шарнирного
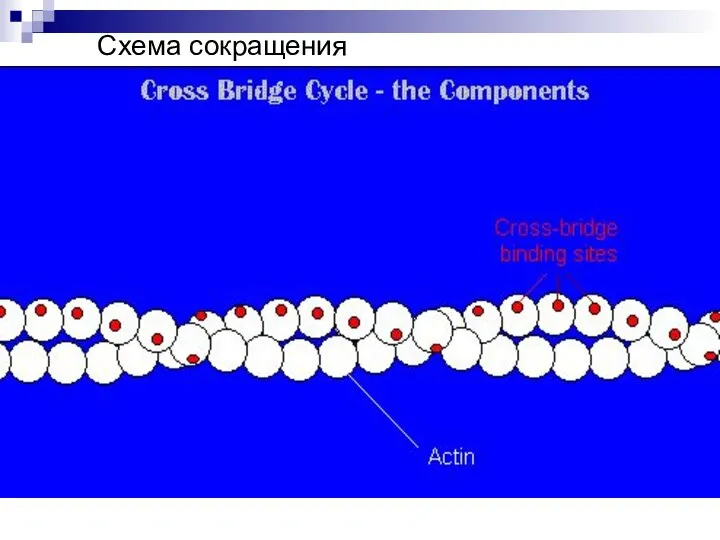
Слайд 31Схема сокращения
Схема сокращения
Слайд 327. Затем головка миозина связывается с молекулой АТФ, что приводит к отделению
7. Затем головка миозина связывается с молекулой АТФ, что приводит к отделению

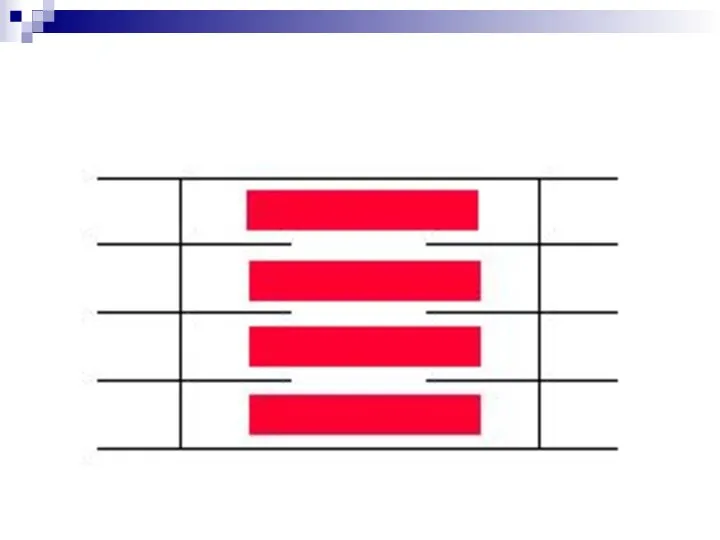
Слайд 34Электро-механическое сопряжение
- это процесс превращения импульса нервного волокна в сокращение мышцы
Электро-механическое сопряжение
- это процесс превращения импульса нервного волокна в сокращение мышцы

Слайд 35Электромеханическое сопряжение
Электромеханическое сопряжение
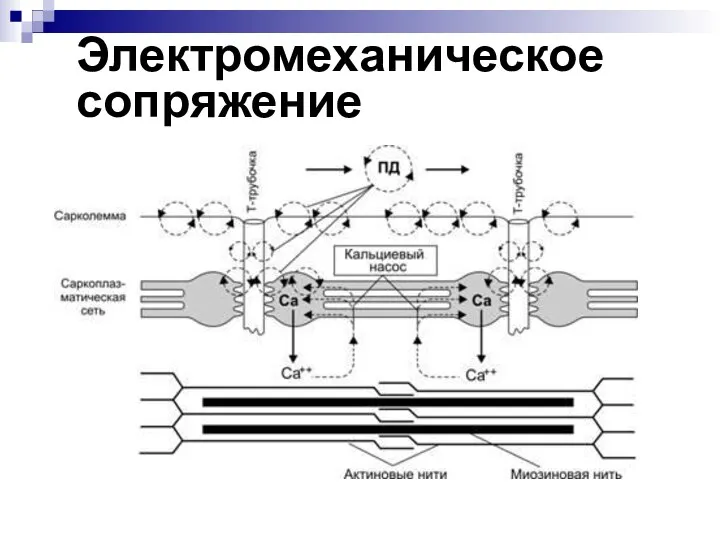
Слайд 36Этапы электромеханического сопряжения
Передача импульса через нервно-мышечный синапс.
Возникновение ВПСП.
Возникновение ПД на околосинаптической
Этапы электромеханического сопряжения
Передача импульса через нервно-мышечный синапс.
Возникновение ВПСП.
Возникновение ПД на околосинаптической

Слайд 37Освобождение Са2+ из СПР и повышение его концентрации в саркоплазме на порядок
Освобождение Са2+ из СПР и повышение его концентрации в саркоплазме на порядок

Слайд 38Каким образом Са2+ выделяется из СПР?
Каким образом Са2+ выделяется из СПР?

Слайд 39Существует несколько гипотез высвобождения Са2+ из СПР:
1. Волна деполяризации спускается по Т-трубочкам
Существует несколько гипотез высвобождения Са2+ из СПР:
1. Волна деполяризации спускается по Т-трубочкам

Слайд 402. Во внешнюю мембрану миоцита встроены потенциалзависимые и/или рецепторуправляемые Са2+-каналы. Небольшое повышение
2. Во внешнюю мембрану миоцита встроены потенциалзависимые и/или рецепторуправляемые Са2+-каналы. Небольшое повышение

Слайд 413. Высвобождение Са2+ из СПР может индуцироваться инозитол–1,4,5–трифосфатом (ИФ3) – вторичным посредником,
3. Высвобождение Са2+ из СПР может индуцироваться инозитол–1,4,5–трифосфатом (ИФ3) – вторичным посредником,
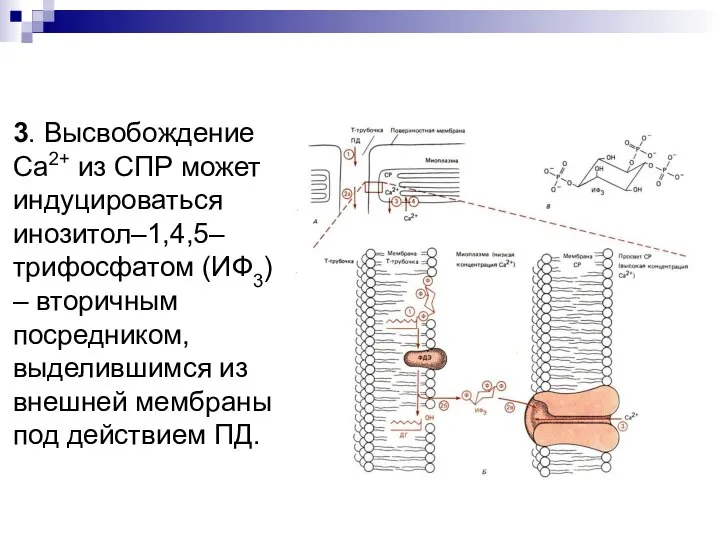
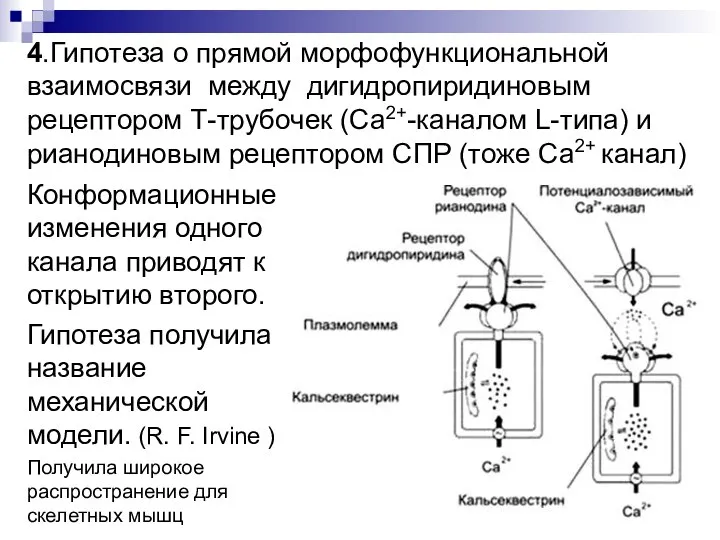
Слайд 424.Гипотеза о прямой морфофункциональной взаимосвязи между дигидропиридиновым рецептором Т-трубочек (Са2+-каналом L-типа) и
4.Гипотеза о прямой морфофункциональной взаимосвязи между дигидропиридиновым рецептором Т-трубочек (Са2+-каналом L-типа) и
Слайд 43Суммация мышечных сокращений
Суммация мышечных сокращений

Слайд 44Одиночное мышечное сокращение
латентный период,
фаза напряжения (укорочения) у быстрых МВ – 7-10 мс,
Одиночное мышечное сокращение
латентный период,
фаза напряжения (укорочения) у быстрых МВ – 7-10 мс,
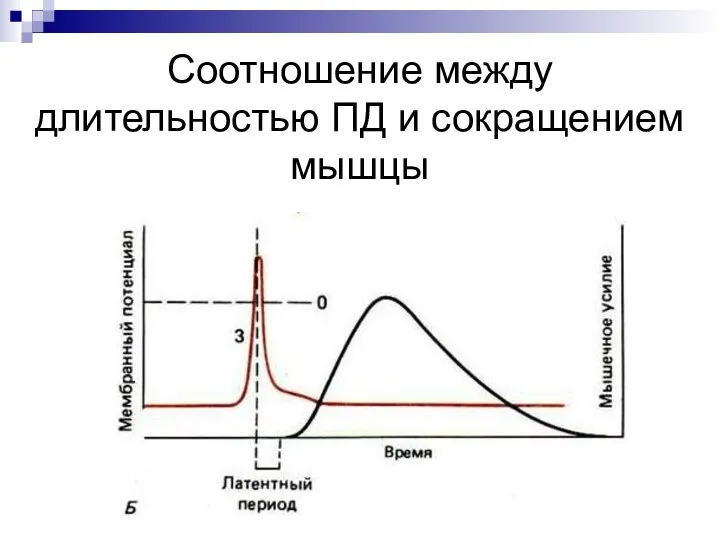
Слайд 45Соотношение между длительностью ПД и сокращением мышцы
Соотношение между длительностью ПД и сокращением мышцы
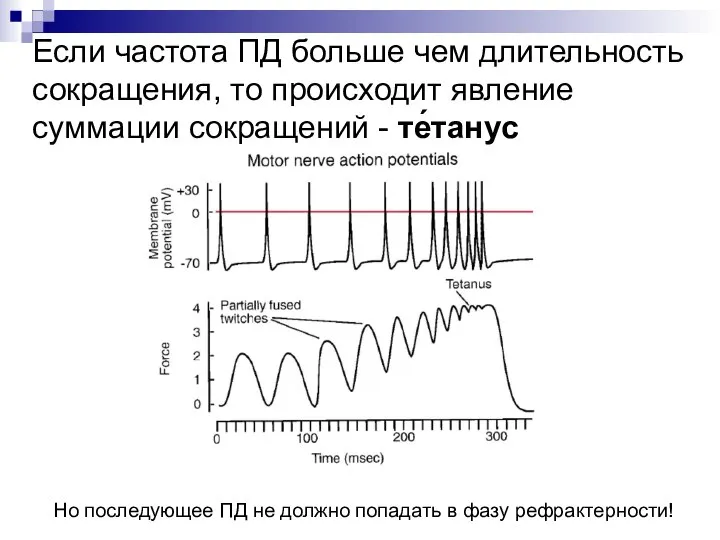
Слайд 46Если частота ПД больше чем длительность сокращения, то происходит явление суммации сокращений
Если частота ПД больше чем длительность сокращения, то происходит явление суммации сокращений
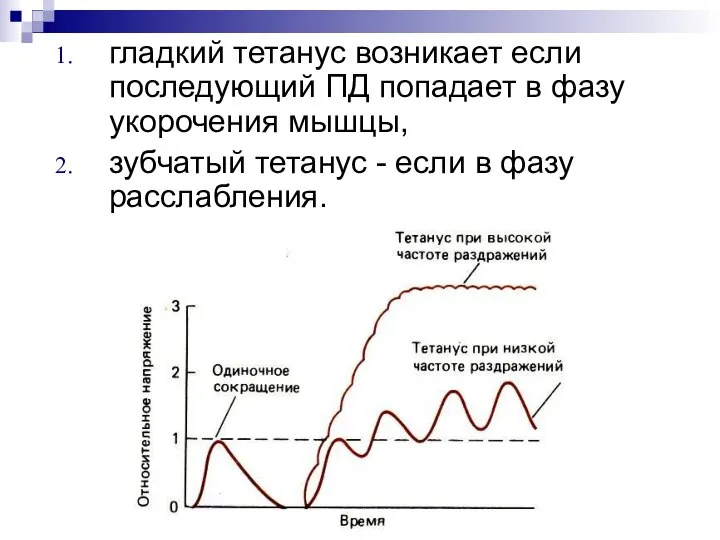
Слайд 47гладкий тетанус возникает если последующий ПД попадает в фазу укорочения мышцы,
зубчатый тетанус
гладкий тетанус возникает если последующий ПД попадает в фазу укорочения мышцы,
зубчатый тетанус
Слайд 48Теории возникновения тетануса
Гельмгольц – тетанус – это механическое сложение одиночных мышечных сокращений,
Теории возникновения тетануса
Гельмгольц – тетанус – это механическое сложение одиночных мышечных сокращений,

Слайд 49КОНТРАКТУРА
- обратимое нераспространяющееся стационарное сокращение
Типы контрактур:
Посттетаническая (высокий остаточный Са2+)
Кофеиновая – кофеин проникая
КОНТРАКТУРА
- обратимое нераспространяющееся стационарное сокращение
Типы контрактур:
Посттетаническая (высокий остаточный Са2+)
Кофеиновая – кофеин проникая

Слайд 50Гладкие мышцы
Гладкие мышцы

Слайд 51Физиологические особенности гладких мышц
1) нестабильный мембранный потенциал, который поддерживает мышцы в состоянии постоянного
Физиологические особенности гладких мышц
1) нестабильный мембранный потенциал, который поддерживает мышцы в состоянии постоянного

Слайд 52Морфологические особенности
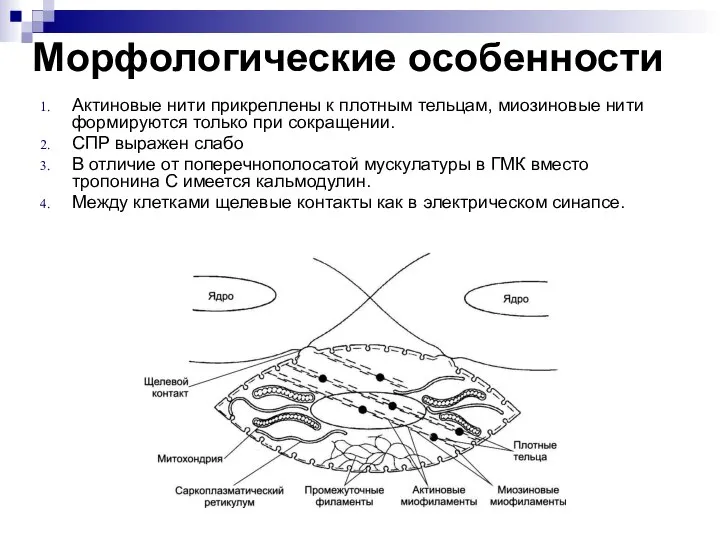
Актиновые нити прикреплены к плотным тельцам, миозиновые нити формируются только при
Морфологические особенности
Актиновые нити прикреплены к плотным тельцам, миозиновые нити формируются только при
Слайд 53Иннервация
ГМК иннервируют симпатические (адренергические) и отчасти парасимпатические (холинергические) нервные волокна.
Нейромедиаторы диффундируют
Иннервация
ГМК иннервируют симпатические (адренергические) и отчасти парасимпатические (холинергические) нервные волокна.
Нейромедиаторы диффундируют

Слайд 54Гуморальная регуляция
В мембрану разных ГМК встроены рецепторы ацетилхолина, гистамина, атриопептина, ангиотензина, адренорецепторы
Гуморальная регуляция
В мембрану разных ГМК встроены рецепторы ацетилхолина, гистамина, атриопептина, ангиотензина, адренорецепторы

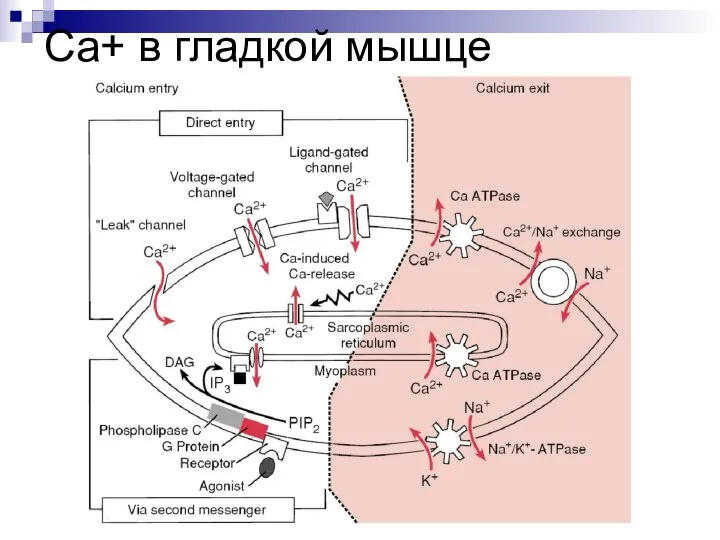
Слайд 55Са+ в гладкой мышце
Са+ в гладкой мышце
Слайд 56Холиномиметики усиливают сокращение
оказывают на мышцу тот же эффект, что и ацетилхолин. Различие
Холиномиметики усиливают сокращение
оказывают на мышцу тот же эффект, что и ацетилхолин. Различие

Слайд 57Кураре и курареподобные препараты широко применяются в анестезиологии.
Тубокурарин препятствует деполяризующему действию
Кураре и курареподобные препараты широко применяются в анестезиологии.
Тубокурарин препятствует деполяризующему действию

Слайд 58Ботулотоксин и столбнячный токсин блокируют секрецию медиатора из нервных терминалей, подавляя торможение
Ботулотоксин и столбнячный токсин блокируют секрецию медиатора из нервных терминалей, подавляя торможение

 Формула цветка
Формула цветка Корнеальный секвестр кошек
Корнеальный секвестр кошек Дыхание
Дыхание Малиновка-Зарянка
Малиновка-Зарянка Экология в нашем классе. 1 класс
Экология в нашем классе. 1 класс Презентация на тему Увеличительные приборы и правила работы с ними
Презентация на тему Увеличительные приборы и правила работы с ними  Мастер - класс по изготовлению травяных мешочков (аромасаше)
Мастер - класс по изготовлению травяных мешочков (аромасаше) Эволюция. 9-11 классы
Эволюция. 9-11 классы Строение и функции спинного мозга
Строение и функции спинного мозга Вiруси
Вiруси Строение и значение кожи
Строение и значение кожи Презентация на тему Лесные рекордсмены
Презентация на тему Лесные рекордсмены  Птицы Свердловской области
Птицы Свердловской области Опорно-двигательная система человека. Гигиена ОДС
Опорно-двигательная система человека. Гигиена ОДС Фотосинтез. Фазы фотосинтеза
Фотосинтез. Фазы фотосинтеза Клетки и ткани человека
Клетки и ткани человека А.Н. Энгельгардт и его роль в развитии агрономии
А.Н. Энгельгардт и его роль в развитии агрономии En el bosque
En el bosque Строение кости. Типы соединения костей
Строение кости. Типы соединения костей Состав крови. Плазма и форменные элементы. Эритроциты. Гемоглобин-оксигемоглобин. СОЭ. Эритропоэз. Лейкоциты и их виды
Состав крови. Плазма и форменные элементы. Эритроциты. Гемоглобин-оксигемоглобин. СОЭ. Эритропоэз. Лейкоциты и их виды Моделирование транспорта питательных веществ в живых организмах. 6 класс
Моделирование транспорта питательных веществ в живых организмах. 6 класс строение и значение нервной системы (1)
строение и значение нервной системы (1) Активность почвенной микрофлоры в агроценозе
Активность почвенной микрофлоры в агроценозе Зимующие птицы
Зимующие птицы Патологические формы эритроцитов
Патологические формы эритроцитов Жизнедеятельность организмов. Размножение растений
Жизнедеятельность организмов. Размножение растений Презентация на тему ОБМЕН ВЕЩЕСТВ
Презентация на тему ОБМЕН ВЕЩЕСТВ  Тип Кишечнополостные, тип Плоские черви, тип Круглые черви, тип Кольчатые черви
Тип Кишечнополостные, тип Плоские черви, тип Круглые черви, тип Кольчатые черви