- Нейрохимические гипотезы шизофрении

Содержание
- 2. Различные симптомы шизофрении связаны со специфичными областями мозга - аномально функционирующими нейронными контурами. Нейронный контур -
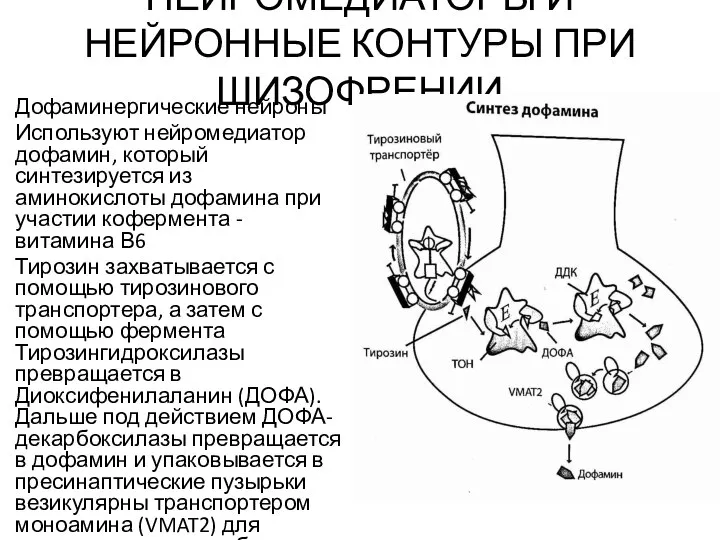
- 3. НЕЙРОМЕДИАТОРЫ И НЕЙРОННЫЕ КОНТУРЫ ПРИ ШИЗОФРЕНИИ Дофаминергические нейроны Используют нейромедиатор дофамин, который синтезируется из аминокислоты дофамина
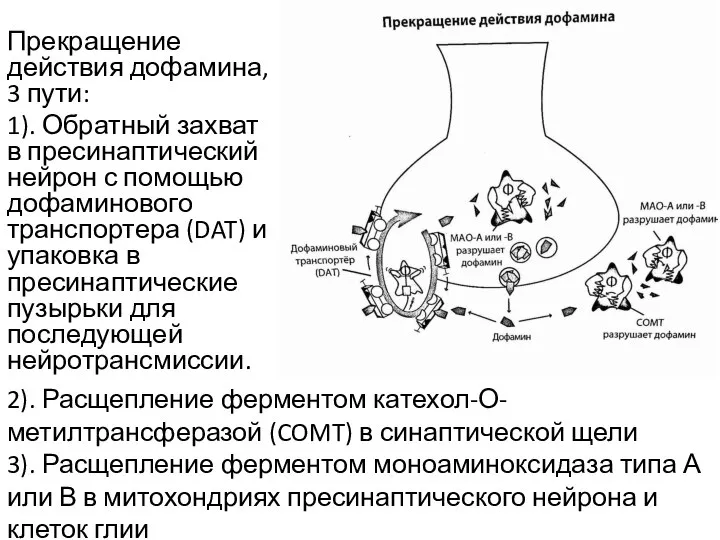
- 4. Прекращение действия дофамина, 3 пути: 1). Обратный захват в пресинаптический нейрон с помощью дофаминового транспортера (DAT)
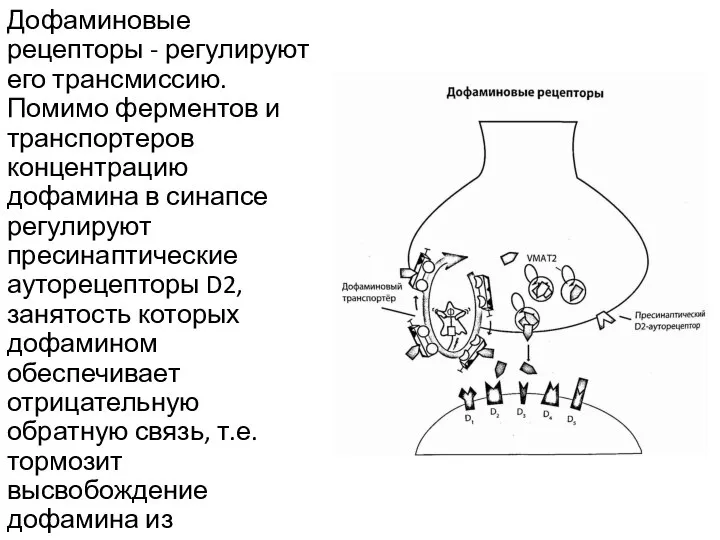
- 5. Дофаминовые рецепторы - регулируют его трансмиссию. Помимо ферментов и транспортеров концентрацию дофамина в синапсе регулируют пресинаптические
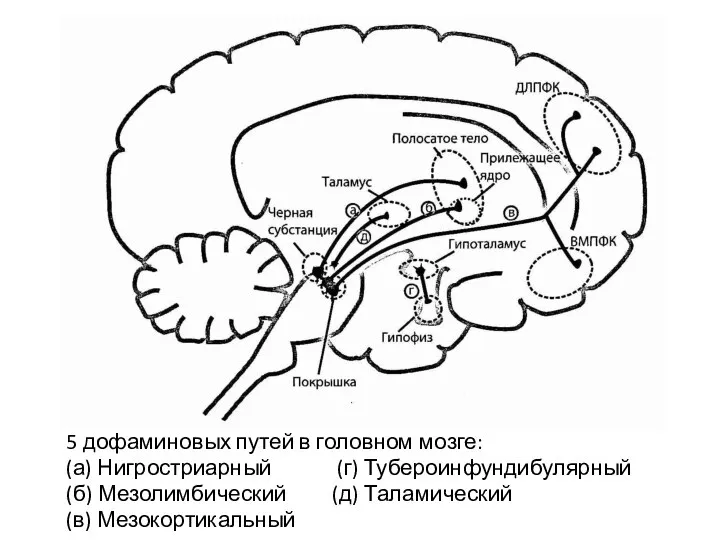
- 6. 5 дофаминовых путей в головном мозге: (а) Нигростриарный (г) Тубероинфундибулярный (б) Мезолимбический (д) Таламический (в) Мезокортикальный
- 7. Нигростриарный – проецируется от черной субстанции к базальным ганглиям или полосатому телу, является частью экстрапирамидной нервной
- 8. Тубероинфундибулярный - идет от гипоталамуса к передней доле гипофиза, регулирует секрецию пролактина. Повышение дофамина в этом
- 9. Мезолимбический - проецируется от вентральной области покрышки в стволе мозга к прилежащему ядру, находящемуся в вентральной
- 10. Мезокортикальный путь: 1 - Идущий к дорсолатеральной префронтальной коре - регулирует когнитивные и исполнительные функции 2
- 11. Негативные симптомы при шизофрении и состояние поведенческого дефицита обуславливаются недостаточной активностью или ненадлежащим функционированием мезокортикальных дофаминовых
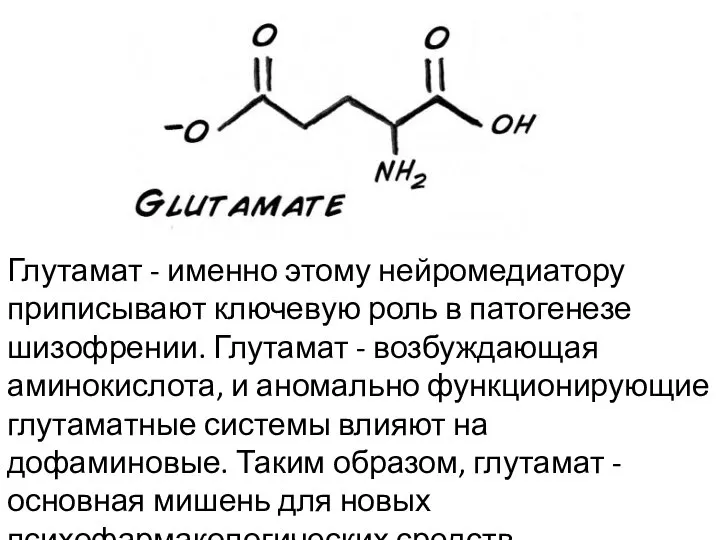
- 12. Глутамат - именно этому нейромедиатору приписывают ключевую роль в патогенезе шизофрении. Глутамат - возбуждающая аминокислота, и
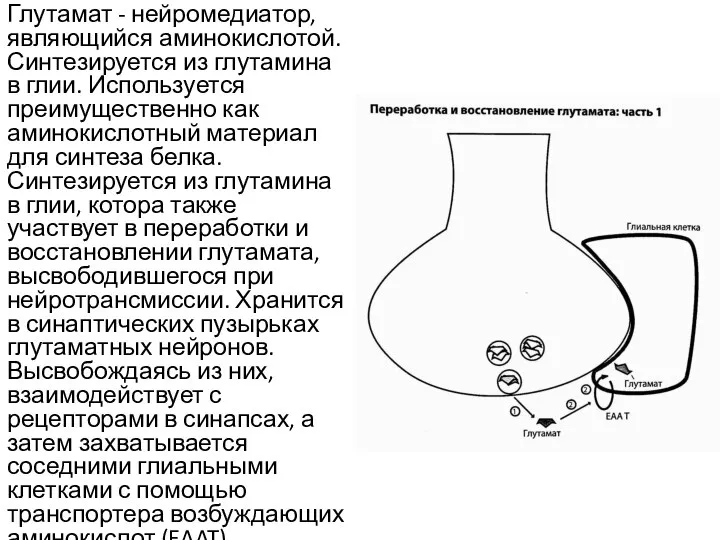
- 13. Глутамат - нейромедиатор, являющийся аминокислотой. Синтезируется из глутамина в глии. Используется преимущественно как аминокислотный материал для
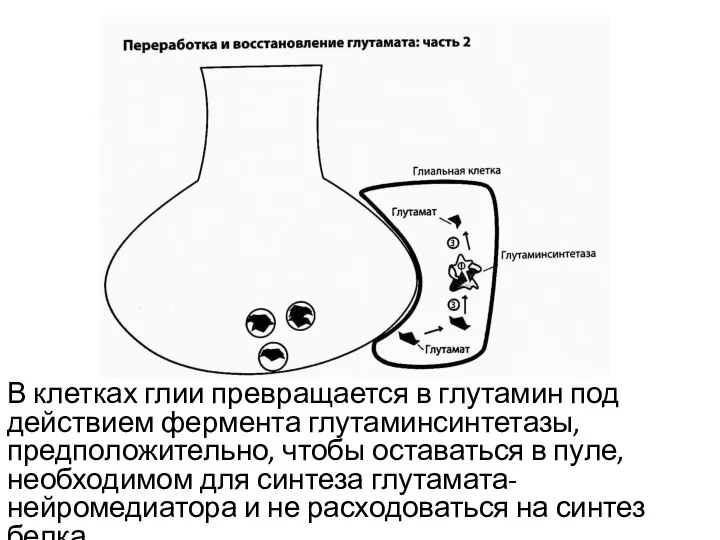
- 14. В клетках глии превращается в глутамин под действием фермента глутаминсинтетазы, предположительно, чтобы оставаться в пуле, необходимом
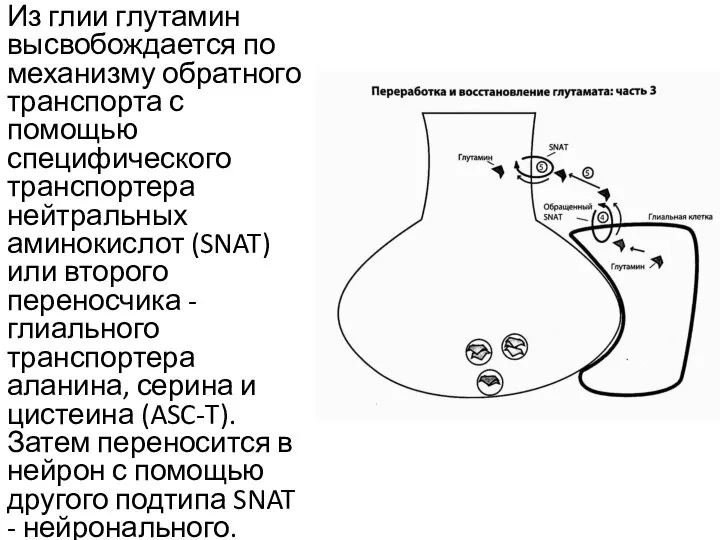
- 15. Из глии глутамин высвобождается по механизму обратного транспорта с помощью специфического транспортера нейтральных аминокислот (SNAT) или
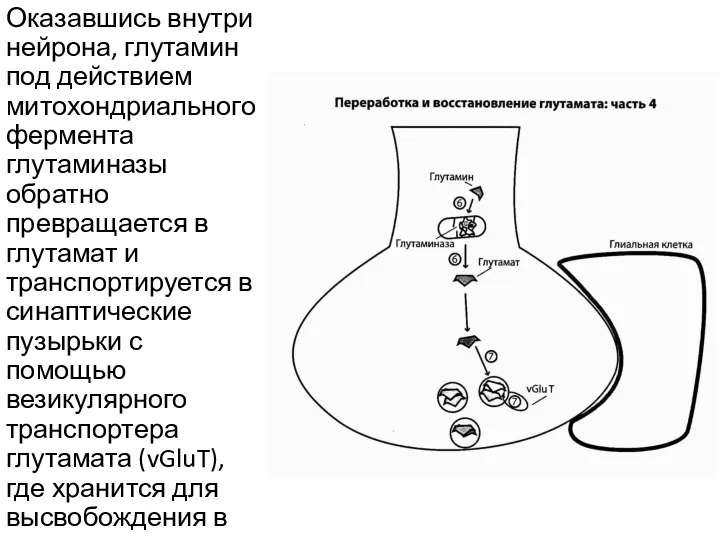
- 16. Оказавшись внутри нейрона, глутамин под действием митохондриального фермента глутаминазы обратно превращается в глутамат и транспортируется в
- 18. В глутаматергической нейротрансмиссии задействованы 4 подтипа глутаматных рецепторов, названия которых происходят от связывающихся с ними агонистов:
- 20. Глицин - комедиатор NMDA-рецепторов Действие глутамата на NMDA-рецепторы частично зависит от наличия комедиаторов - глицина или
- 22. D-серин - комедиатор NMDA-рецепторов. L-серин попадает в глиальные клетки с помощью транспортера L-серина, и фермент серин-рацемаза
- 24. Хотя глутамат как возбуждающий нейромедиатор может оказывать воздействие практически на все нейроны в головном мозге, существуют
- 26. Гипотеза о гипофункции NMDA-рецепторов при шизофрении - основная современная нейрохимическая гипотеза шизофрении, согласно которой глутаматная активность
- 28. Гипотетический участок глутаматной дисфункции при шизофрении - на некоторых вставочных ГАМКергических нейронах коры. По всей видимости,
- 30. В парвальбумин-содержащих вставочных ГАМКергических нейронах префронтальной коры головного мозга пациентов с шизофренией имеются и другие дефекты,
- 32. Кортико-стволовые глутаматные пути регулируют выход глутамата из коры в нейротрансмиттерный центр ствола мозга (вентральную область покрышки),
- 34. Гипофункция NMDA-рецепторов глутаматергических синапсов в вентральном гиппокампе тоже может приводить к мезолимбической дофаминовой гиперактивности. В норме
- 35. Г
- 36. Связь гипотезы о гипофункции NMDA-рецепторов при шизофрении с дофаминовой гипотизой: негативные симптомы Корково-стволовой глутаматный путь посредством
- 38. Скачать презентацию
Слайд 2Различные симптомы шизофрении связаны со специфичными областями мозга - аномально функционирующими нейронными
Различные симптомы шизофрении связаны со специфичными областями мозга - аномально функционирующими нейронными

Слайд 3НЕЙРОМЕДИАТОРЫ И НЕЙРОННЫЕ КОНТУРЫ ПРИ ШИЗОФРЕНИИ
Дофаминергические нейроны
Используют нейромедиатор дофамин, который синтезируется из
НЕЙРОМЕДИАТОРЫ И НЕЙРОННЫЕ КОНТУРЫ ПРИ ШИЗОФРЕНИИ
Дофаминергические нейроны
Используют нейромедиатор дофамин, который синтезируется из
Слайд 4Прекращение действия дофамина, 3 пути:
1). Обратный захват в пресинаптический нейрон с помощью
Прекращение действия дофамина, 3 пути:
1). Обратный захват в пресинаптический нейрон с помощью
Слайд 5Дофаминовые рецепторы - регулируют его трансмиссию. Помимо ферментов и транспортеров концентрацию дофамина
Дофаминовые рецепторы - регулируют его трансмиссию. Помимо ферментов и транспортеров концентрацию дофамина
Слайд 65 дофаминовых путей в головном мозге:
(а) Нигростриарный (г) Тубероинфундибулярный
(б) Мезолимбический (д) Таламический
(в)
5 дофаминовых путей в головном мозге:
(а) Нигростриарный (г) Тубероинфундибулярный
(б) Мезолимбический (д) Таламический
(в)
Слайд 7Нигростриарный – проецируется от черной субстанции к базальным ганглиям или полосатому телу,
Нигростриарный – проецируется от черной субстанции к базальным ганглиям или полосатому телу,
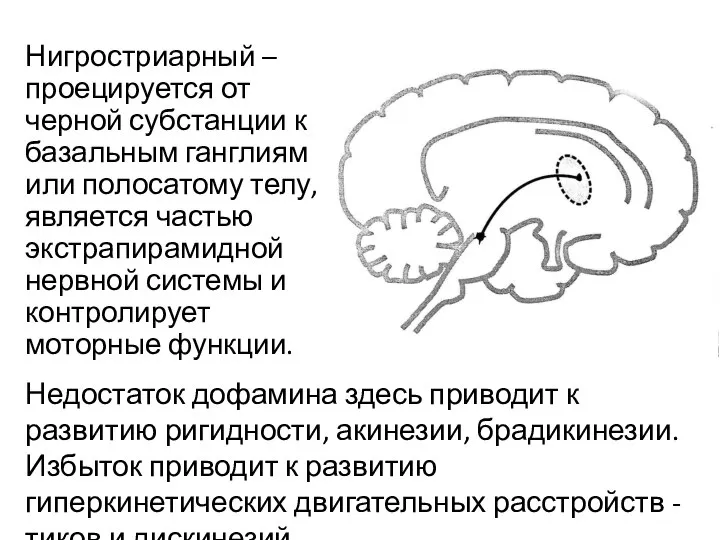
Слайд 8Тубероинфундибулярный - идет от гипоталамуса к передней доле гипофиза, регулирует секрецию пролактина.
Тубероинфундибулярный - идет от гипоталамуса к передней доле гипофиза, регулирует секрецию пролактина.
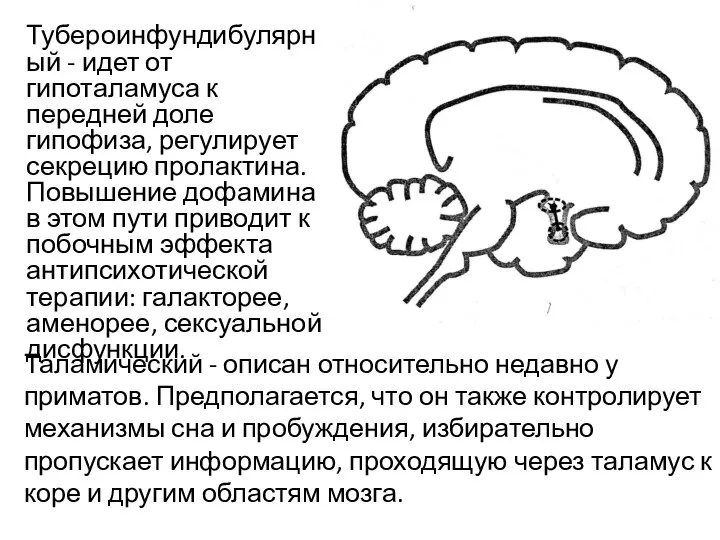
Слайд 9Мезолимбический - проецируется от вентральной области покрышки в стволе мозга к прилежащему
Мезолимбический - проецируется от вентральной области покрышки в стволе мозга к прилежащему
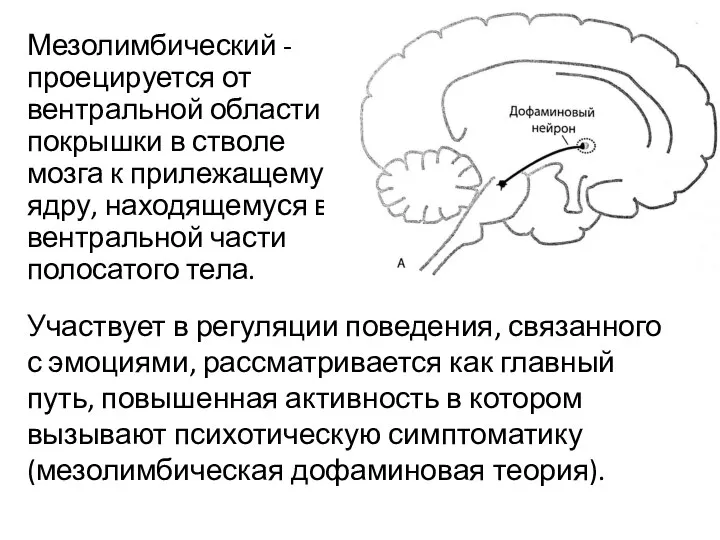
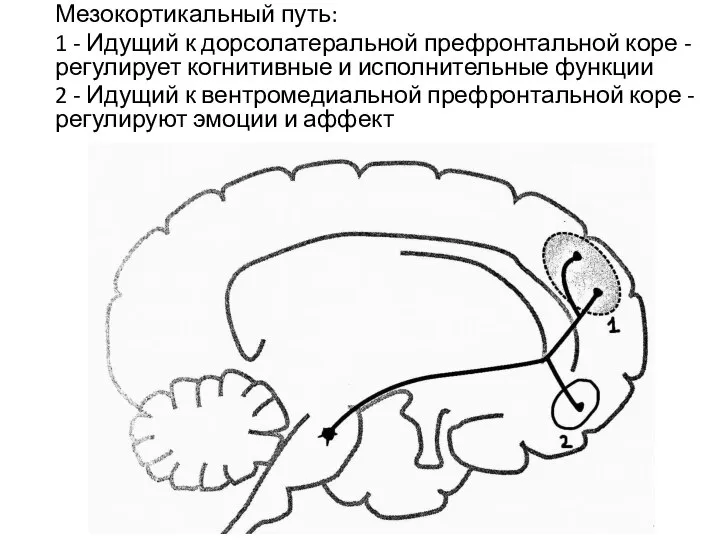
Слайд 10Мезокортикальный путь:
1 - Идущий к дорсолатеральной префронтальной коре - регулирует когнитивные и
Мезокортикальный путь:
1 - Идущий к дорсолатеральной префронтальной коре - регулирует когнитивные и
Слайд 11Негативные симптомы при шизофрении и состояние поведенческого дефицита обуславливаются недостаточной активностью или
Негативные симптомы при шизофрении и состояние поведенческого дефицита обуславливаются недостаточной активностью или

Слайд 12Глутамат - именно этому нейромедиатору приписывают ключевую роль в патогенезе шизофрении. Глутамат
Глутамат - именно этому нейромедиатору приписывают ключевую роль в патогенезе шизофрении. Глутамат
Слайд 13Глутамат - нейромедиатор, являющийся аминокислотой. Синтезируется из глутамина в глии. Используется преимущественно
Глутамат - нейромедиатор, являющийся аминокислотой. Синтезируется из глутамина в глии. Используется преимущественно
Слайд 14В клетках глии превращается в глутамин под действием фермента глутаминсинтетазы, предположительно, чтобы
В клетках глии превращается в глутамин под действием фермента глутаминсинтетазы, предположительно, чтобы
Слайд 15Из глии глутамин высвобождается по механизму обратного транспорта с помощью специфического транспортера
Из глии глутамин высвобождается по механизму обратного транспорта с помощью специфического транспортера
Слайд 16Оказавшись внутри нейрона, глутамин под действием митохондриального фермента глутаминазы обратно превращается в
Оказавшись внутри нейрона, глутамин под действием митохондриального фермента глутаминазы обратно превращается в
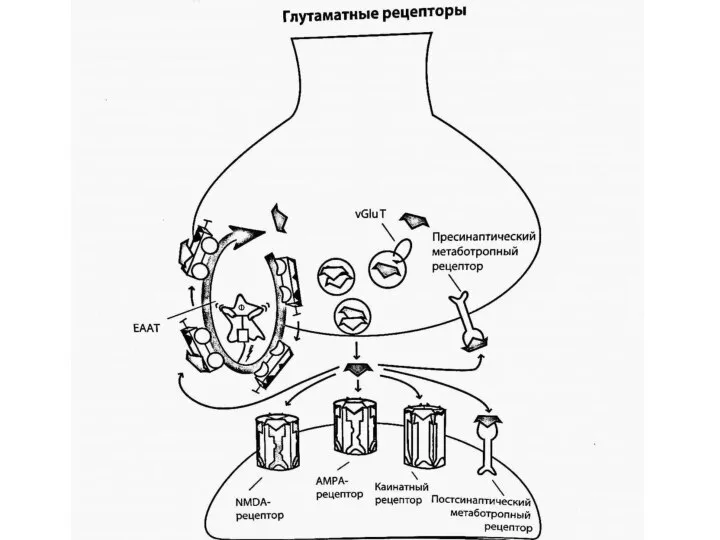
Слайд 18В глутаматергической нейротрансмиссии задействованы 4 подтипа глутаматных рецепторов, названия которых происходят от
В глутаматергической нейротрансмиссии задействованы 4 подтипа глутаматных рецепторов, названия которых происходят от

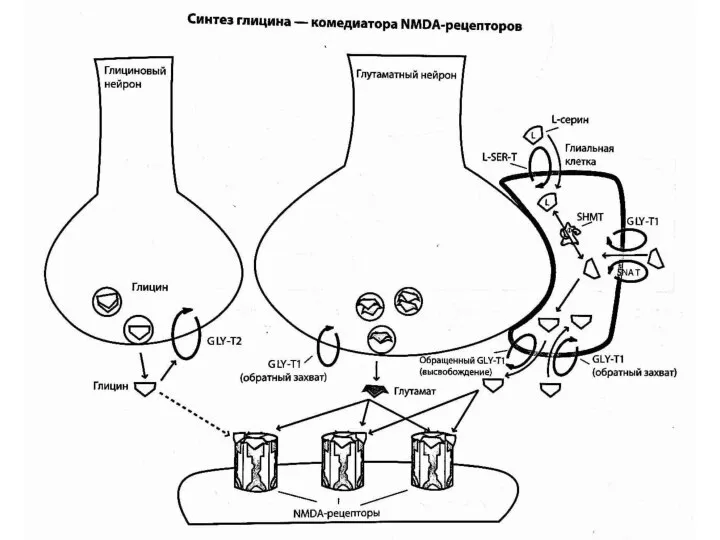
Слайд 20Глицин - комедиатор NMDA-рецепторов
Действие глутамата на NMDA-рецепторы частично зависит от наличия комедиаторов
Глицин - комедиатор NMDA-рецепторов Действие глутамата на NMDA-рецепторы частично зависит от наличия комедиаторов

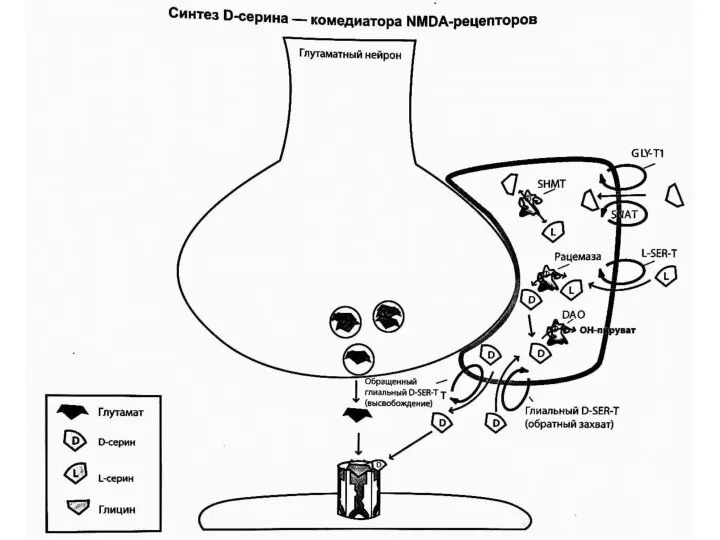
Слайд 22D-серин - комедиатор NMDA-рецепторов.
L-серин попадает в глиальные клетки с помощью транспортера L-серина,
D-серин - комедиатор NMDA-рецепторов. L-серин попадает в глиальные клетки с помощью транспортера L-серина,

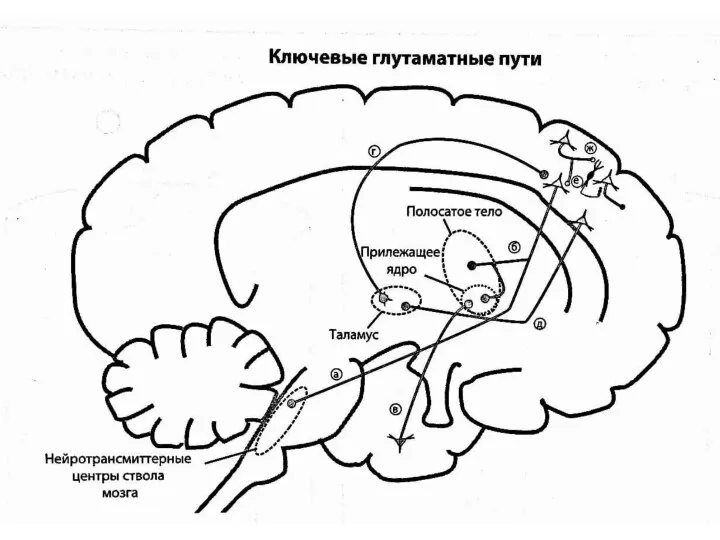
Слайд 24Хотя глутамат как возбуждающий нейромедиатор может оказывать воздействие практически на все нейроны
Хотя глутамат как возбуждающий нейромедиатор может оказывать воздействие практически на все нейроны

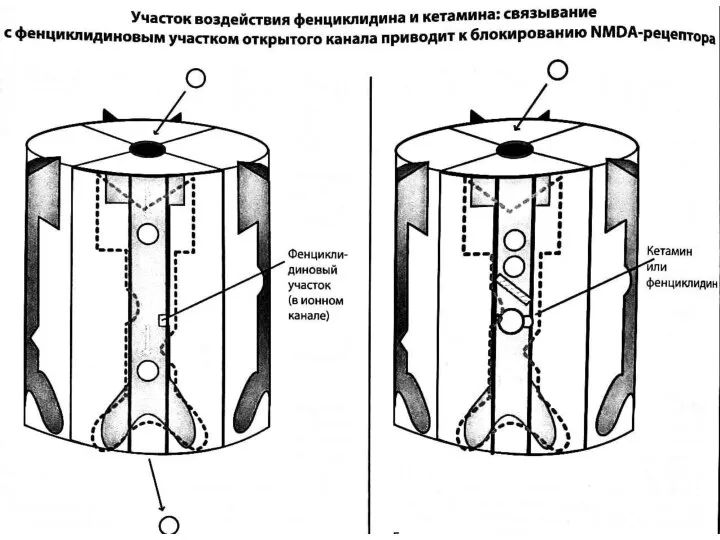
Слайд 26Гипотеза о гипофункции NMDA-рецепторов при шизофрении - основная современная нейрохимическая гипотеза шизофрении,
Гипотеза о гипофункции NMDA-рецепторов при шизофрении - основная современная нейрохимическая гипотеза шизофрении,

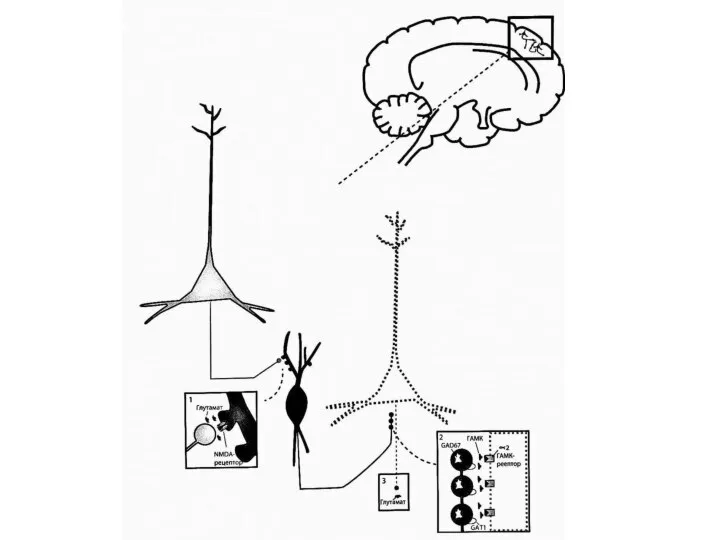
Слайд 28Гипотетический участок глутаматной дисфункции при шизофрении - на некоторых вставочных ГАМКергических нейронах
Гипотетический участок глутаматной дисфункции при шизофрении - на некоторых вставочных ГАМКергических нейронах

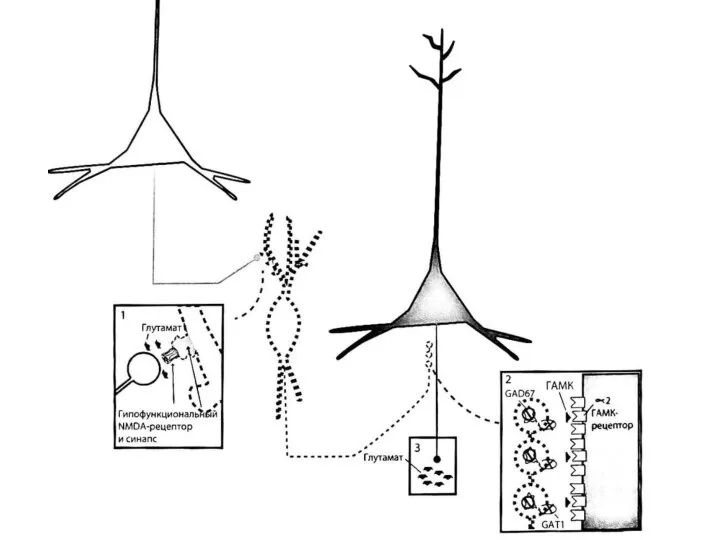
Слайд 30В парвальбумин-содержащих вставочных ГАМКергических нейронах префронтальной коры головного мозга пациентов с шизофренией
В парвальбумин-содержащих вставочных ГАМКергических нейронах префронтальной коры головного мозга пациентов с шизофренией

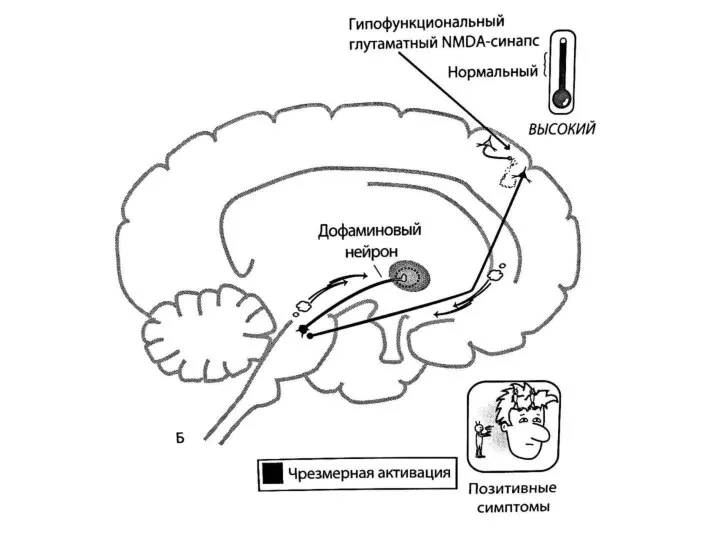
Слайд 32
Кортико-стволовые глутаматные пути регулируют выход глутамата из коры в нейротрансмиттерный центр ствола
Кортико-стволовые глутаматные пути регулируют выход глутамата из коры в нейротрансмиттерный центр ствола

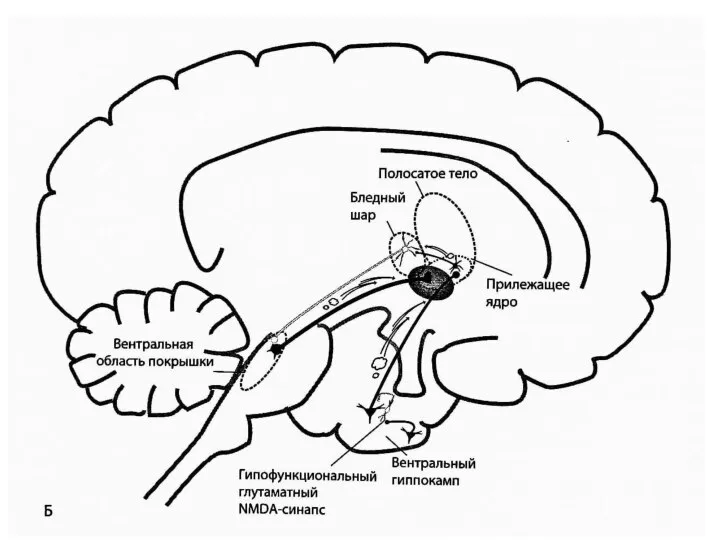
Слайд 34Гипофункция NMDA-рецепторов глутаматергических синапсов в вентральном гиппокампе тоже может приводить к мезолимбической
Гипофункция NMDA-рецепторов глутаматергических синапсов в вентральном гиппокампе тоже может приводить к мезолимбической

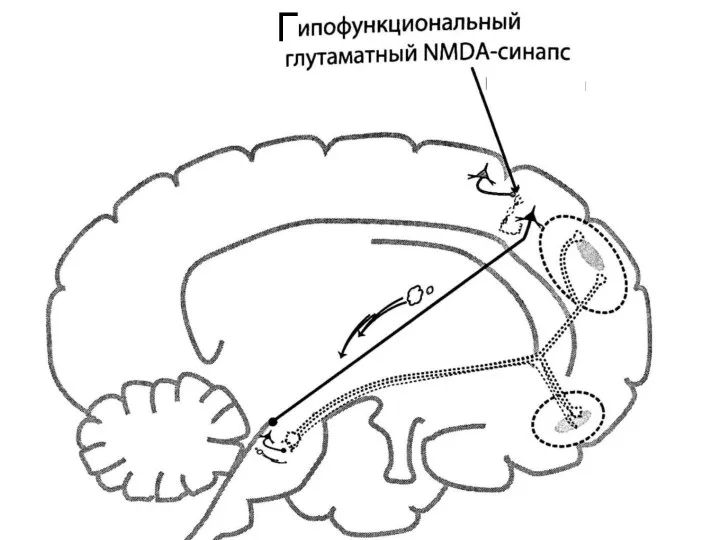
Слайд 36Связь гипотезы о гипофункции NMDA-рецепторов при шизофрении с дофаминовой гипотизой: негативные симптомы
Корково-стволовой
Связь гипотезы о гипофункции NMDA-рецепторов при шизофрении с дофаминовой гипотизой: негативные симптомы Корково-стволовой

 Презентация на тему ТЕЛА И ВЕЩЕСТВА
Презентация на тему ТЕЛА И ВЕЩЕСТВА  Основные группы микроорганизмов
Основные группы микроорганизмов Отгадай грызуна зоопарка
Отгадай грызуна зоопарка Грибы. Часть 2
Грибы. Часть 2 Эти забавные домашние питомцы
Эти забавные домашние питомцы Грибная пора. Интерактивная игра
Грибная пора. Интерактивная игра Питание ягодных культур
Питание ягодных культур Көбелек сабағы
Көбелек сабағы Группы биотических связей между организмами
Группы биотических связей между организмами Тип членистоногие
Тип членистоногие Близнецы в генетике
Близнецы в генетике Что растёт на клумбе
Что растёт на клумбе Особенности сенсорной функции у детей и подростков
Особенности сенсорной функции у детей и подростков Педагогическая физиология детей и подростков
Педагогическая физиология детей и подростков Деление клетки (митоз)
Деление клетки (митоз) Механизмы регуляции ферментативной активности
Механизмы регуляции ферментативной активности Органы цветкового растения
Органы цветкового растения Изучение автоматизированной системы управления воздушным режимом
Изучение автоматизированной системы управления воздушным режимом Ученика 9б класса СОШ № 288 КОШЕЛЕВА НИКИТЫ
Ученика 9б класса СОШ № 288 КОШЕЛЕВА НИКИТЫ Защита растений в ландшафте
Защита растений в ландшафте ГМО. Аргументы за
ГМО. Аргументы за Паразитизм
Паразитизм Презентация на тему Первые вестники весны - Первоцветы
Презентация на тему Первые вестники весны - Первоцветы  Картофель - пищевое паслёновое растение
Картофель - пищевое паслёновое растение Презентация на тему Человеческий мозг
Презентация на тему Человеческий мозг  Отдел покрытосеменные: капустные, маковые
Отдел покрытосеменные: капустные, маковые Розы (фотографии)
Розы (фотографии) Растения
Растения