- Обмен аминокислот (Раздел 4)

Содержание
- 2. ПЛАН ЛЕКЦИИ I. Переваривание белков и всасывание АК в ЖКТ. II. Аминокислотный фонд клетки. III. Метаболизм
- 3. СПИСОК СОКРАЩЕНИЙ АК - аминокислоты ЖКТ – желудочно-кишечный тракт Гл – SH – глутатион ГАМК –
- 4. I. ПЕРЕВАРИВАНИЕ БЕЛКОВ И ВСАСЫВАНИЕ АК В ЖКТ 1) Белки пищи поэтапно гидролизуются до свободных АК
- 5. I. ПЕРЕВАРИВАНИЕ БЕЛКОВ И ВСАСЫВАНИЕ АК В ЖКТ 2) Пептидазы обладают относительной субстратной специфичностью, но с
- 6. I. ПЕРЕВАРИВАНИЕ БЕЛКОВ И ВСАСЫВАНИЕ АК В ЖКТ 4) От пептидаз СОЖ и кишечника защищают муцины,
- 7. II. АМИНОКИСЛОТНЫЙ ФОНД КЛЕТКИ ИСТОЧНИКИ ПОСТУПЛЕНИЯ АК: 1) белки пищи; 2) белки тканей 3) синтез заменимых
- 8. III. Метаболизм АК в тканях: общие пути обмена АК 1. ТРАНСАМИНИРОВАНИЕ – это реакции переноса аминогруппы
- 9. Рис. 1. РЕАКЦИИ ТРАНСАМИНИРОВАНИЯ α- α- АСТ АЛТ ПФ(В6) ПФ(В6)
- 10. Рис.2 Механизм трансаминирования (Браунштейн А.Е. и Шемякин М.М.)
- 11. Б/РОЛЬ РЕАКЦИЙ ТРАНСАМИНИРОВАНИЯ Образование заменимых АК из α-кетокислот (α-КГ- коллектор, собирает аминогруппы; Глу - универсальный донор
- 12. III. Метаболизм АК в тканях: общие пути обмена АК 2. ДЕЗАМИНИРОВАНИЕ – это реакции отщеплениия α-аминогруппы
- 13. ПРЯМОЕ ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ АК НАДН+Н+ ФП1 (ЦПЭ) 3 АТФ РИС.3
- 14. Непрямое окислительное дезаминирование АК 2 этапа: РИС.4
- 15. НЕПРЯМОЕ ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ ТРАНСФЕРАЗА α- ПФ(В6) РИС. 5 2 1
- 16. Биологическая РОЛЬ РЕАКЦИЙ ДЕЗАМИНИРОВАНИЯ АК Реакции распада АК Образование α-кетокислот, к-е могут поступать В ЦТК (ЩУК,
- 17. III. Метаболизм АК в тканях: общие пути обмена АК 3. ДЕКАРБОКСИЛИРОВАНИЕ – это необратимые реакции отщеплениия
- 18. III. Метаболизм АК в тканях: общие пути обмена АК б) Декарбоксилирование ДОФА в нервной ткани и
- 19. III. Метаболизм АК в тканях: общие пути обмена АК в) Декарбоксилирование Глу: Биологическая роль ГАМК: основной
- 20. III. Метаболизм АК в тканях: общие пути обмена АК г) Декарбоксилирование 5-(ОН)-Три РИС.9 5 5 (нервная
- 21. IV. ИНАКТИВАЦИЯ БИОГЕННЫХ АМИНОВ I.Реакции метилирования (для гистамина,адреналина) РИС.10
- 22. IV. ИНАКТИВАЦИЯ БИОГЕННЫХ АМИНОВ 2. Реакции окислительного дезаминирования (для дофамина, НА, серотонина, ГАМК) РИС.11 Биологическая роль
- 23. V. Метаболизм АК в тканях: индивидуальные пути обмена АК РИС.12 Биологическая роль реакции: образование Глу, к-я
- 24. V. Метаболизм АК в тканях: индивидуальные пути синтеза АК РИС.13 б) СИНТЕЗ ТИРОЗИНА Биологическая роль реакции:
- 25. V. Метаболизм АК в тканях: индивидуальные пути обмена АК в) СИНТЕЗ ЦИСТЕИНА ИЗ МЕТИОНИНА АТФ Н2О
- 26. V. Метаболизм АК в тканях: индивидуальные пути обмена АК 1 ТРАНСМЕТИЛИРОВАНИЕ СУБСТРАТОВ (R) НЕОБХОДИМО ДЛЯ: СИНТЕЗА
- 27. Б/РОЛЬ ЦИСТЕИНА (Цис) 1) Цистеин содержат многие ферменты (ДГ, ЩФ), гормоны (кальцитонин, АДГ, инсулин), Гл-SH; Цис
- 28. VI. ПРИЛОЖЕНИЕ: ГОМОЦИСТИНУРИЯ В основе заболевания гомоцистинурия (МКБ 10: Е72.1) лежит метаболический блок на пути превращения
- 29. VI. ПРИЛОЖЕНИЕ: ГОМОЦИСТИНУРИЯ Повышенное содержание метаболитов метионина в крови и тканях оказывает токсическое действие на нервную
- 30. VI. ПРИЛОЖЕНИЕ: рис.1.Специфичность действия пепсина ПЕПСИНОГЕН ПЕПСИН Рис. 1 Пептоны Белки Желудок рН 1,5-2,0
- 31. Пептоны Рис. 2 Олигопептиды + аминокислоты Про Сер Аром. АК VI. ПРИЛОЖЕНИЕ: рис. 2. Специфичность действия
- 32. VI. ПРИЛОЖЕНИЕ: рис. 3 М.Г. Крицман (1905-1971) Окончила Одесский медицинский институт. Работала в Москве в Биохимическом
- 33. VI. ПРИЛОЖЕНИЕ: рис. 4 РИС.4
- 34. VI. ПРИЛОЖЕНИЕ: рис.5. Виды реакций дезаминирования РИС.5
- 35. VI. ПРИЛОЖЕНИЕ: рис.6. : Неокислительное дезаминирование Сер,Тре (элиминирующее) РИС. 6
- 36. VI. ПРИЛОЖЕНИЕ: рис.7 Неокислительное дезаминирование ЦИС (цистатионин - γ – лиаза, ПФ,) и неокислительное (внутримолекулярное) дезаминирование
- 37. VI.ПРИЛОЖЕНИЕ: рис.8 Непрямое дезаминирование АК с участием цикла ИМФ – АМФ (для мышечной ткани и мозга)
- 38. VI. ПРИЛОЖЕНИЕ: рис.9. Декарбоксилирование серусодержащих АК РИС. 9
- 39. VI. ПРИЛОЖЕНИЕ: рис.10. ОБМЕН Тир в меланоцитах РИС. 10 1 1
- 40. VI. ПРИЛОЖЕНИЕ: рис.11. Биосинтез мелатонина и ниацина (В3) из Три РИС.11 НАД+
- 41. СПИСОК ЛИТЕРАТУРЫ ЛК 6 Основная литература: 1. а) Биохимия: учебник/ под ред. Е.С. Северина.- М.: ГЭОТАР-
- 42. VII. СПИСОК ЛИТЕРАТУРЫ ЛК 6 Дополнительная литература: Биологическая химия: учебник. Т.Т. Березов, Б.Ф. Коровкин. – 3-е
- 44. Скачать презентацию

Слайд 3 СПИСОК СОКРАЩЕНИЙ
АК - аминокислоты
ЖКТ – желудочно-кишечный тракт
Гл – SH –
СПИСОК СОКРАЩЕНИЙ
АК - аминокислоты
ЖКТ – желудочно-кишечный тракт
Гл – SH –

Слайд 4 I. ПЕРЕВАРИВАНИЕ БЕЛКОВ И ВСАСЫВАНИЕ АК В ЖКТ
1) Белки пищи поэтапно
I. ПЕРЕВАРИВАНИЕ БЕЛКОВ И ВСАСЫВАНИЕ АК В ЖКТ
1) Белки пищи поэтапно

Слайд 5 I. ПЕРЕВАРИВАНИЕ БЕЛКОВ И ВСАСЫВАНИЕ АК В ЖКТ
2) Пептидазы обладают
I. ПЕРЕВАРИВАНИЕ БЕЛКОВ И ВСАСЫВАНИЕ АК В ЖКТ
2) Пептидазы обладают

Слайд 6I. ПЕРЕВАРИВАНИЕ БЕЛКОВ И ВСАСЫВАНИЕ АК В ЖКТ
4) От пептидаз СОЖ
I. ПЕРЕВАРИВАНИЕ БЕЛКОВ И ВСАСЫВАНИЕ АК В ЖКТ
4) От пептидаз СОЖ

Слайд 7 II. АМИНОКИСЛОТНЫЙ ФОНД КЛЕТКИ
ИСТОЧНИКИ ПОСТУПЛЕНИЯ АК:
1) белки пищи; 2)
II. АМИНОКИСЛОТНЫЙ ФОНД КЛЕТКИ
ИСТОЧНИКИ ПОСТУПЛЕНИЯ АК:
1) белки пищи; 2)

Слайд 8III. Метаболизм АК в тканях: общие пути обмена АК
1. ТРАНСАМИНИРОВАНИЕ –
III. Метаболизм АК в тканях: общие пути обмена АК
1. ТРАНСАМИНИРОВАНИЕ –

Слайд 9 Рис. 1. РЕАКЦИИ ТРАНСАМИНИРОВАНИЯ
α-
α-
АСТ
АЛТ
ПФ(В6)
ПФ(В6)
Рис. 1. РЕАКЦИИ ТРАНСАМИНИРОВАНИЯ
α-
α-
АСТ
АЛТ
ПФ(В6)
ПФ(В6)

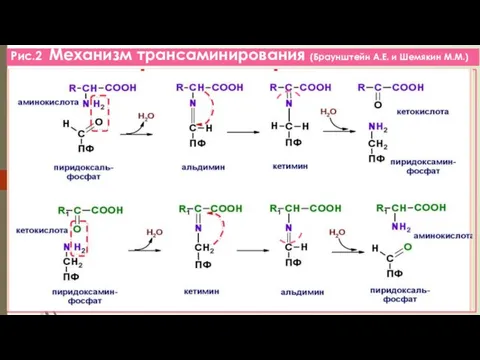
Слайд 10Рис.2 Механизм трансаминирования (Браунштейн А.Е. и Шемякин М.М.)
Рис.2 Механизм трансаминирования (Браунштейн А.Е. и Шемякин М.М.)
Слайд 11 Б/РОЛЬ РЕАКЦИЙ ТРАНСАМИНИРОВАНИЯ
Образование заменимых АК из α-кетокислот
(α-КГ- коллектор, собирает аминогруппы;
Б/РОЛЬ РЕАКЦИЙ ТРАНСАМИНИРОВАНИЯ
Образование заменимых АК из α-кетокислот
(α-КГ- коллектор, собирает аминогруппы;

Слайд 12III. Метаболизм АК в тканях: общие пути обмена АК
2. ДЕЗАМИНИРОВАНИЕ –
III. Метаболизм АК в тканях: общие пути обмена АК
2. ДЕЗАМИНИРОВАНИЕ –

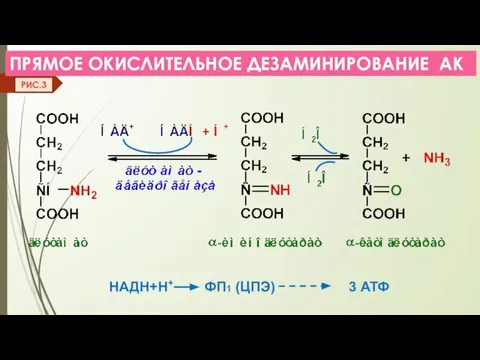
Слайд 13ПРЯМОЕ ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ АК
НАДН+Н+ ФП1 (ЦПЭ) 3 АТФ
РИС.3
ПРЯМОЕ ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ АК
НАДН+Н+ ФП1 (ЦПЭ) 3 АТФ
РИС.3
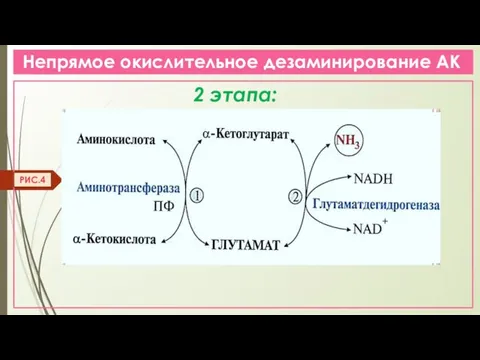
Слайд 14 Непрямое окислительное дезаминирование АК
2 этапа:
РИС.4
Непрямое окислительное дезаминирование АК
2 этапа:
РИС.4
Слайд 15 НЕПРЯМОЕ ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ
ТРАНСФЕРАЗА
α-
ПФ(В6)
РИС. 5
2
1
НЕПРЯМОЕ ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ
ТРАНСФЕРАЗА
α-
ПФ(В6)
РИС. 5
2
1

Слайд 16Биологическая РОЛЬ РЕАКЦИЙ ДЕЗАМИНИРОВАНИЯ АК
Реакции распада АК
Образование α-кетокислот, к-е могут поступать
Биологическая РОЛЬ РЕАКЦИЙ ДЕЗАМИНИРОВАНИЯ АК
Реакции распада АК
Образование α-кетокислот, к-е могут поступать

Слайд 17III. Метаболизм АК в тканях: общие пути обмена АК
3. ДЕКАРБОКСИЛИРОВАНИЕ –
III. Метаболизм АК в тканях: общие пути обмена АК
3. ДЕКАРБОКСИЛИРОВАНИЕ –

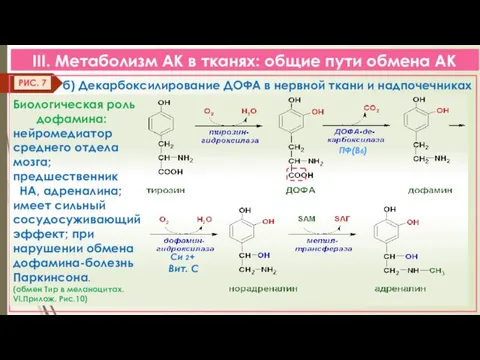
Слайд 18 III. Метаболизм АК в тканях: общие пути обмена АК
б) Декарбоксилирование
III. Метаболизм АК в тканях: общие пути обмена АК
б) Декарбоксилирование
Слайд 19III. Метаболизм АК в тканях: общие пути обмена АК
в) Декарбоксилирование Глу:
Биологическая
III. Метаболизм АК в тканях: общие пути обмена АК
в) Декарбоксилирование Глу:
Биологическая

Слайд 20III. Метаболизм АК в тканях: общие пути обмена АК
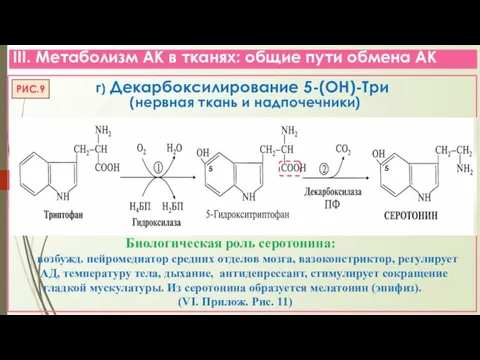
г) Декарбоксилирование 5-(ОН)-Три
III. Метаболизм АК в тканях: общие пути обмена АК
г) Декарбоксилирование 5-(ОН)-Три
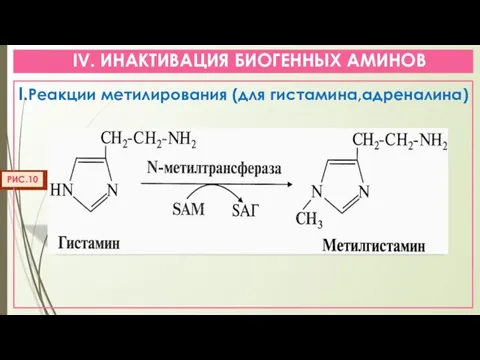
Слайд 21 IV. ИНАКТИВАЦИЯ БИОГЕННЫХ АМИНОВ
I.Реакции метилирования (для гистамина,адреналина)
РИС.10
IV. ИНАКТИВАЦИЯ БИОГЕННЫХ АМИНОВ
I.Реакции метилирования (для гистамина,адреналина)
РИС.10
Слайд 22 IV. ИНАКТИВАЦИЯ БИОГЕННЫХ АМИНОВ
2. Реакции окислительного дезаминирования
(для дофамина,
IV. ИНАКТИВАЦИЯ БИОГЕННЫХ АМИНОВ
2. Реакции окислительного дезаминирования
(для дофамина,

Слайд 23V. Метаболизм АК в тканях: индивидуальные пути обмена АК
РИС.12
Биологическая роль реакции:
V. Метаболизм АК в тканях: индивидуальные пути обмена АК
РИС.12
Биологическая роль реакции:

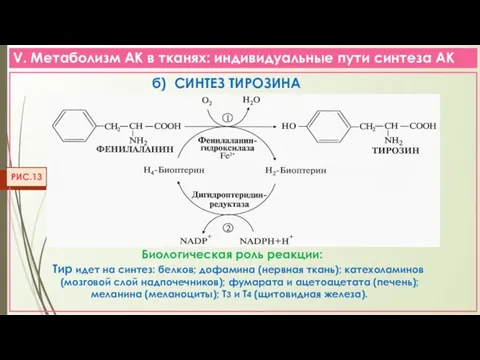
Слайд 24V. Метаболизм АК в тканях: индивидуальные пути синтеза АК
РИС.13
б) СИНТЕЗ ТИРОЗИНА
V. Метаболизм АК в тканях: индивидуальные пути синтеза АК
РИС.13
б) СИНТЕЗ ТИРОЗИНА
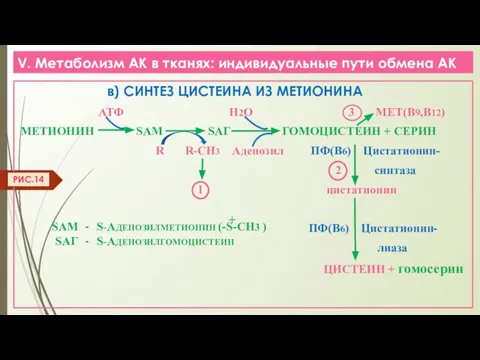
Слайд 25V. Метаболизм АК в тканях: индивидуальные пути обмена АК
в) СИНТЕЗ ЦИСТЕИНА
V. Метаболизм АК в тканях: индивидуальные пути обмена АК
в) СИНТЕЗ ЦИСТЕИНА
Слайд 26V. Метаболизм АК в тканях: индивидуальные пути обмена АК
1 ТРАНСМЕТИЛИРОВАНИЕ СУБСТРАТОВ
V. Метаболизм АК в тканях: индивидуальные пути обмена АК
1 ТРАНСМЕТИЛИРОВАНИЕ СУБСТРАТОВ

Слайд 27 Б/РОЛЬ ЦИСТЕИНА (Цис)
1) Цистеин содержат многие ферменты (ДГ, ЩФ), гормоны
Б/РОЛЬ ЦИСТЕИНА (Цис)
1) Цистеин содержат многие ферменты (ДГ, ЩФ), гормоны

Слайд 28 VI. ПРИЛОЖЕНИЕ: ГОМОЦИСТИНУРИЯ
В основе заболевания гомоцистинурия (МКБ 10: Е72.1)
VI. ПРИЛОЖЕНИЕ: ГОМОЦИСТИНУРИЯ
В основе заболевания гомоцистинурия (МКБ 10: Е72.1)

Слайд 29 VI. ПРИЛОЖЕНИЕ: ГОМОЦИСТИНУРИЯ
Повышенное содержание метаболитов метионина в крови и
VI. ПРИЛОЖЕНИЕ: ГОМОЦИСТИНУРИЯ
Повышенное содержание метаболитов метионина в крови и

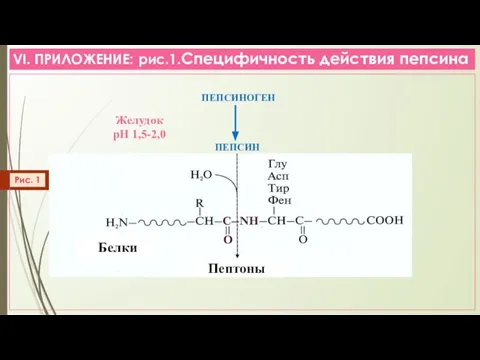
Слайд 30VI. ПРИЛОЖЕНИЕ: рис.1.Специфичность действия пепсина
ПЕПСИНОГЕН
ПЕПСИН
Рис. 1
Пептоны
Белки
Желудок
рН 1,5-2,0
VI. ПРИЛОЖЕНИЕ: рис.1.Специфичность действия пепсина
ПЕПСИНОГЕН
ПЕПСИН
Рис. 1
Пептоны
Белки
Желудок
рН 1,5-2,0
Слайд 31 Пептоны
Рис. 2
Олигопептиды + аминокислоты
Про
Сер
Аром. АК
VI. ПРИЛОЖЕНИЕ: рис. 2. Специфичность действия панкреатических
Пептоны
Рис. 2
Олигопептиды + аминокислоты
Про
Сер
Аром. АК
VI. ПРИЛОЖЕНИЕ: рис. 2. Специфичность действия панкреатических

Слайд 32 VI. ПРИЛОЖЕНИЕ: рис. 3
М.Г. Крицман
(1905-1971)
Окончила Одесский медицинский институт. Работала
VI. ПРИЛОЖЕНИЕ: рис. 3
М.Г. Крицман
(1905-1971)
Окончила Одесский медицинский институт. Работала

Слайд 33 VI. ПРИЛОЖЕНИЕ: рис. 4
РИС.4
VI. ПРИЛОЖЕНИЕ: рис. 4
РИС.4

Слайд 34VI. ПРИЛОЖЕНИЕ: рис.5. Виды реакций дезаминирования
РИС.5
VI. ПРИЛОЖЕНИЕ: рис.5. Виды реакций дезаминирования
РИС.5

Слайд 35VI. ПРИЛОЖЕНИЕ: рис.6.
:
Неокислительное дезаминирование Сер,Тре (элиминирующее)
РИС. 6
VI. ПРИЛОЖЕНИЕ: рис.6.
:
Неокислительное дезаминирование Сер,Тре (элиминирующее)
РИС. 6

Слайд 36 VI. ПРИЛОЖЕНИЕ: рис.7
Неокислительное дезаминирование ЦИС
(цистатионин - γ
VI. ПРИЛОЖЕНИЕ: рис.7
Неокислительное дезаминирование ЦИС
(цистатионин - γ

Слайд 37 VI.ПРИЛОЖЕНИЕ: рис.8
Непрямое дезаминирование АК
с участием цикла ИМФ –
VI.ПРИЛОЖЕНИЕ: рис.8
Непрямое дезаминирование АК
с участием цикла ИМФ –

Слайд 38VI. ПРИЛОЖЕНИЕ: рис.9. Декарбоксилирование серусодержащих АК
РИС. 9
VI. ПРИЛОЖЕНИЕ: рис.9. Декарбоксилирование серусодержащих АК
РИС. 9

Слайд 39
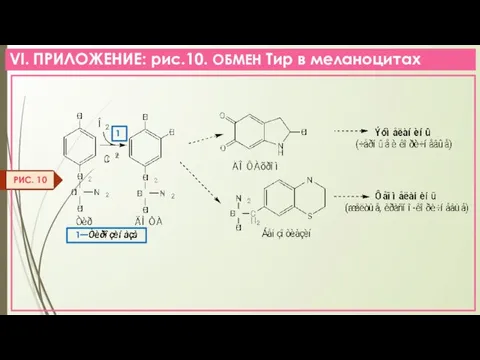
VI. ПРИЛОЖЕНИЕ: рис.10. ОБМЕН Тир в меланоцитах
РИС. 10
1
1
VI. ПРИЛОЖЕНИЕ: рис.10. ОБМЕН Тир в меланоцитах
РИС. 10
1
1
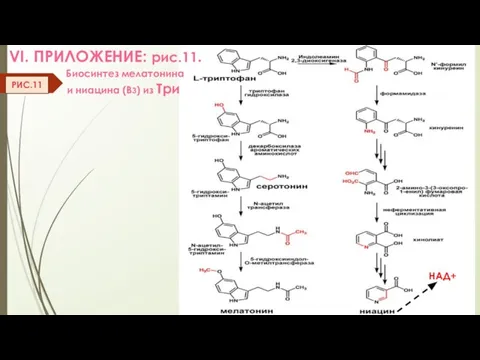
Слайд 40VI. ПРИЛОЖЕНИЕ: рис.11.
Биосинтез мелатонина
и ниацина (В3) из Три
РИС.11
НАД+
VI. ПРИЛОЖЕНИЕ: рис.11.
Биосинтез мелатонина
и ниацина (В3) из Три
РИС.11
НАД+
Слайд 41 СПИСОК ЛИТЕРАТУРЫ
ЛК 6
Основная литература:
1. а) Биохимия: учебник/ под
СПИСОК ЛИТЕРАТУРЫ
ЛК 6
Основная литература:
1. а) Биохимия: учебник/ под

Слайд 42 VII. СПИСОК ЛИТЕРАТУРЫ
ЛК 6
Дополнительная литература:
Биологическая химия: учебник. Т.Т.
VII. СПИСОК ЛИТЕРАТУРЫ
ЛК 6
Дополнительная литература:
Биологическая химия: учебник. Т.Т.

 Презентация на тему Половое размножение
Презентация на тему Половое размножение  Отруйні гриби
Отруйні гриби Структура квітки
Структура квітки Зачем человеку растения
Зачем человеку растения Простые и сложные методы окрашивания микроорганизмов: окраска по Граму, окраска спор, капсул, жгутиков
Простые и сложные методы окрашивания микроорганизмов: окраска по Граму, окраска спор, капсул, жгутиков Многообразие земноводных
Многообразие земноводных Опыты с растениями. Банан. 7 класс
Опыты с растениями. Банан. 7 класс Клуб любителей кошек
Клуб любителей кошек Членистоногие обитающие в Республике Крым
Членистоногие обитающие в Республике Крым Презентация на тему Цитология
Презентация на тему Цитология  Презентация на тему Рациональное питание залог здоровья
Презентация на тему Рациональное питание залог здоровья  pril
pril Нервная система человека
Нервная система человека Ткани
Ткани Виороиды. Инфекционные молекулы РНК, способные вызывать заболевания высших растений
Виороиды. Инфекционные молекулы РНК, способные вызывать заболевания высших растений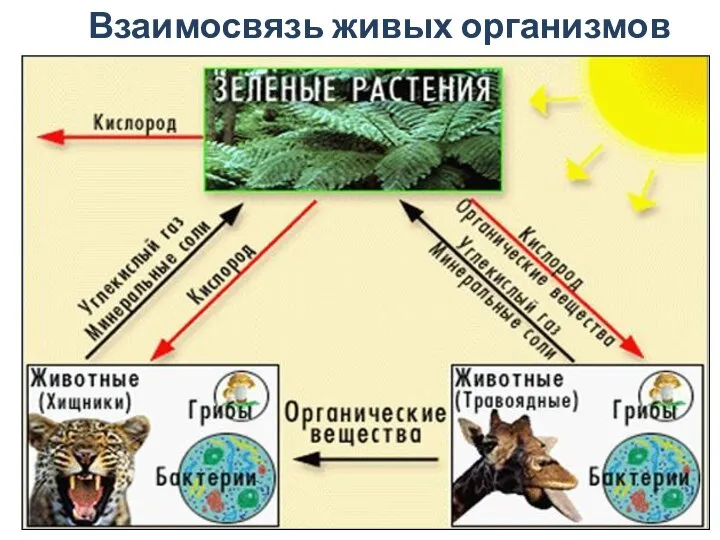 Биосфера. Часть 3. Взаимосвязь живых организмов
Биосфера. Часть 3. Взаимосвязь живых организмов Техника размораживания спермы перед осеменением
Техника размораживания спермы перед осеменением Царство Грибы
Царство Грибы Синица. Внешний вид
Синица. Внешний вид Zheludochno-kishechny_trakt_Prezentatsia_3
Zheludochno-kishechny_trakt_Prezentatsia_3 Красная книга Карелии
Красная книга Карелии Царство грибы (часть 1)
Царство грибы (часть 1) Гигиена сенсорных систем
Гигиена сенсорных систем Анализаторы и органы чувств
Анализаторы и органы чувств Анатомия человека. Понятие о костях
Анатомия человека. Понятие о костях Тема 1
Тема 1 Низшие растения. Водоросли
Низшие растения. Водоросли Презентация на тему МИКРООРГАНИЗМЫ-ХУДОЖНИКИ
Презентация на тему МИКРООРГАНИЗМЫ-ХУДОЖНИКИ