- Обмен азота. Пути синтеза аминокислот. Лекция 3

Содержание
- 3. Как живые организмы получают азот?
- 4. ЦИКЛ АЗОТА В БИОСФЕРЕ СИНТЕЗ АМИНОКИСЛОТ, по учебнику Анисимова п. 5.1 и 5.2 по: Нельсон, Кокс
- 5. Для синтеза заменимых аминокислот человеку и животным подходит только NH3 (в воде NH4OH, NH4+)
- 6. Источники аммиака Фриц Габер, Карл Бош – Нобелевская премия 1918 и 1931 соответственно за создание технологии
- 7. Цикл азота в биосфере 1) диазотрофы 2) нитрификация (бактерии) 3) поглощение и ассимиляция NH4+ и NO3-
- 8. 1. Фиксация NΞN. НИТРОГЕНАЗА БАКТЕРИИ – диазотрофы: - свободноживущие (рр. Azotobacter, Clostridium, все фотосинтезирующие, ряд АРХЕЙ…)
- 9. Повсеместность азотфиксации 1895 - Наббе, Хилтнер – патент на микробное удобрение Nitragin (культуры азотфиксаторов для разных
- 10. Общая схема азотфиксации У Rhizobium для восстановления 1N2 может идти от 12 до 35 АТФ –
- 11. Легоглобины – защита от О2 Из люпина Миоглобин кашалота
- 12. “Из аммиака в аммонийную форму” - работает нитрогеназный комплекс (упрощенно называемый нитрогеназой). Комплекс состоит из 2х
- 13. Нитрогеназный комплекс Fe-белок (редуктаза динитрогеназы) – каждый из 2х - димер MoFe-белок (динитрогеназа) - тетрамер, имеет
- 14. Кластеры нитрогеназы
- 15. FeS-центры в Fe-белке Р-кластер (8Fe-7S) 4Fe-4S-кластер
- 16. MoFe-кофактор: что в центре? Атомы: cерые – Fe (7), желтые – S (9), черные – С,
- 17. Работа нитрогеназы N2 + 8H+ + 8e + 16АТФ→ 2NH3 + H2 + 16АДФ + 16Фн
- 18. Расшифровка к предыдущему слайду В молекуле Fe-белка (димере) 1 FeS-центр, поэтому Fe-белок может переносить только по
- 19. Ферредоксины (Фд, Fd) Ферредоксины (ferrum — железо; сокращённо обозначается «Фд») — группа небольших (6—12 кДа) растворимых
- 20. Дополнительный материал Направления исследований: 1) Разработка технологий для сельского хозяйства на базе изучения физиологии и биохимии
- 21. Дополнительный материал: Гены и белки азотфиксации Вершинина З.Р., Баймиев Ал.Х., Чемерис А.В. Симбиотические реакции корней облепихи,
- 22. ПУТИ СИНТЕЗА АМИНОКИСЛОТ Семейства аминокислот по происхождению
- 23. Из аммиака в аминокислоты
- 24. РАСТЕНИЕ: поглощение и переработка NH4+ из почвы (или от симбионтов) Дополнит. информация:2 типа транспортных систем: если
- 25. Прямое восстановительное аминирование – 1 путь синтеза а-к-т Фермент – глутаматдегидрогеназа (глутаматДГ) НАДН – никотинамид-аденин-динуклеотид в
- 26. Образование глутамина – 1а путь синтеза Фермент - глутаминсинтетаза
- 27. Переаминирование – 3 путь синтеза аминокислот Ферменты переаминирования – аминотрансферазы (например, аланин-амино-трансфераза (АлАт), аспартатаминотрансфераза (АсАт)
- 28. 3 путь синтеза аминокислот - ферментативные превращения 1) Семейства аминокислот 1 – 4 (на основе α-кетоглутаровой
- 29. Семейства аминокислот 1-4
- 30. Гистидин – особый путь ! У животных гис не синтезируется
- 31. Семейство 5 (ароматические) – по шикиматному пути 1) синтез дегидрохинной кислоты (дегидрохинната), у большинства видов -
- 32. 1) Основной путь синтеза дегидрохинната
- 33. Главные ступени синтеза аминокислот из шикимата ! У животных фен и три – не синтезируются, тир
- 34. Шикиматный путь (начиная с дегидрохинната)
- 35. ИТОГО: основные пути синтеза белковых аминокислот 1) прямое восстановительное аминирование 1а) – образование амидов 2) переаминирование
- 36. Ферментативные превращения в синтезе аминокислот 1) Семейства аминокислот 1 – 4 (на основе α-кетоглутаровой к-ты, ПВК,
- 37. Семейства аминокислот 1-4
- 38. Гистидин – особый путь ! У животных гис не синтезируется
- 39. Семейство 5 (ароматические) – по шикиматному пути 1) синтез дегидрохинной кислоты (дегидрохинната), у большинства видов -
- 40. 1) Основной путь синтеза дегидрохинната
- 41. Главные ступени синтеза аминокислот из шикимата ! У животных фен и три – не синтезируются, тир
- 42. Шикиматный путь (начиная с дегидрохинната)
- 43. Нитрификация: 1) автотрофная (бактерии сем. Nitrobacteriaceae, археи), при окислении аммиака получающие энергию в виде АТФ, которую
- 44. Автотрофная нитрификация у бактерий a) NH4+ в клетку – транспортный белок Сu-содержащая транслоказа б) образование нитрита
- 45. Денитрификация (из нитрата в N2)
- 46. Денитрификация. Анаммокс Недавнее открытие (1999г) – возможность окисления аммиака бактериями в анаэробных условиях NH4++NO2- = N2
- 47. Бактерии АНАММОКС – загадка на загадке... - до сих пор не выделены в чистую культуру (“кандидаты”
- 48. Селеноцистеин (Sec) Формилметионин
- 49. Синтез формилметионина – прямо на тРНК, в 2 стадии
- 50. Селеноцистеин образуется из серина прямо на тРНК
- 51. γ-аминомасляная кислота (ГАМК) основной нейромедиатор торможения, рецепторы ГАМК A и ГАМК C – ионные каналы, активация
- 52. Синтез нетипичных аминокислот ПОСЛЕ построения белка РЕАКЦИИ - Ацилирование (метилирование и др.) по –NH2-группе бокового радикала
- 53. Роль ɣ-карбоксиглутаминовой кислоты в улучшении связывания Са2+ Образуется ПОСЛЕ ТРАНСЛЯЦИИ (из ГЛУ, находящейся уже в составе
- 54. Mucuna (сем. Бобовые) – зудящие бобы Гриффония (Griffonia simplicifolia, сем. Бобовые) 3,4-дигидроксифенилаланин 5-гидрокситриптофан Ландыш, юкки ЗАЩИТА.
- 55. как в синтезе серотонина
- 56. Синтез азетидин-2-карбоновой кислоты Convallaria majalis (до 7% сухого веса листьев). Очень мало – в сахарной свекле.
- 57. Синтез β-циан(о)аланина - токсина β-циан-L-аланин синтаза КФ 4.4.1.9 (L-цистеин-сероводород-лиаза (добавляющая HCN), ее кофактор пиридоксальфосфат бобовых (Lupinus
- 59. Скачать презентацию
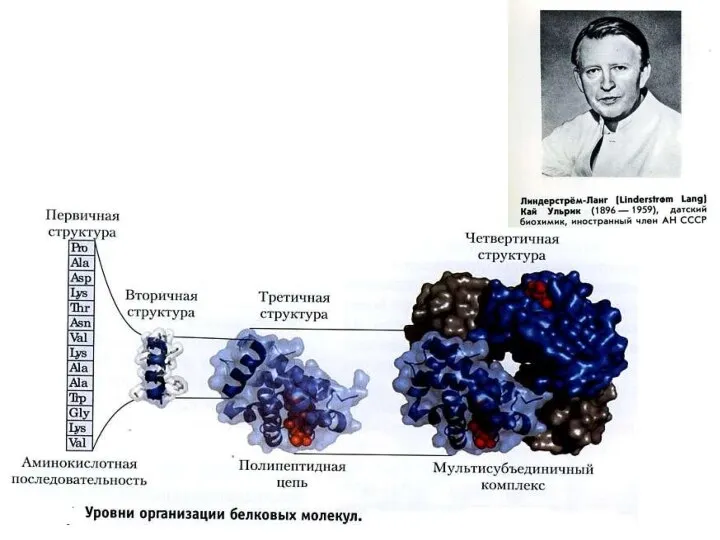
Слайд 3Как живые организмы получают азот?
Как живые организмы получают азот?

Слайд 4ЦИКЛ АЗОТА В БИОСФЕРЕ
СИНТЕЗ АМИНОКИСЛОТ,
по учебнику Анисимова п. 5.1 и 5.2
СИНТЕЗ АМИНОКИСЛОТ,
по учебнику Анисимова п. 5.1 и 5.2

Слайд 5Для синтеза заменимых аминокислот человеку и животным подходит только NH3 (в воде
Для синтеза заменимых аминокислот человеку и животным подходит только NH3 (в воде

Слайд 6Источники аммиака
Фриц Габер, Карл Бош – Нобелевская премия 1918 и 1931 соответственно
Источники аммиака
Фриц Габер, Карл Бош – Нобелевская премия 1918 и 1931 соответственно

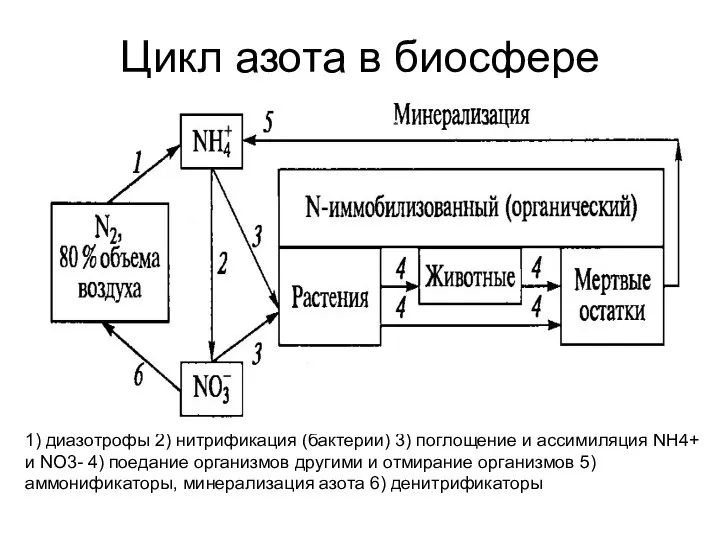
Слайд 7Цикл азота в биосфере
1) диазотрофы 2) нитрификация (бактерии) 3) поглощение и ассимиляция
Цикл азота в биосфере
1) диазотрофы 2) нитрификация (бактерии) 3) поглощение и ассимиляция
Слайд 81. Фиксация NΞN. НИТРОГЕНАЗА
БАКТЕРИИ – диазотрофы:
- свободноживущие (рр. Azotobacter, Clostridium, все фотосинтезирующие,
1. Фиксация NΞN. НИТРОГЕНАЗА
БАКТЕРИИ – диазотрофы:
- свободноживущие (рр. Azotobacter, Clostridium, все фотосинтезирующие,

Слайд 9Повсеместность азотфиксации
1895 - Наббе, Хилтнер – патент на микробное удобрение Nitragin (культуры
Повсеместность азотфиксации
1895 - Наббе, Хилтнер – патент на микробное удобрение Nitragin (культуры

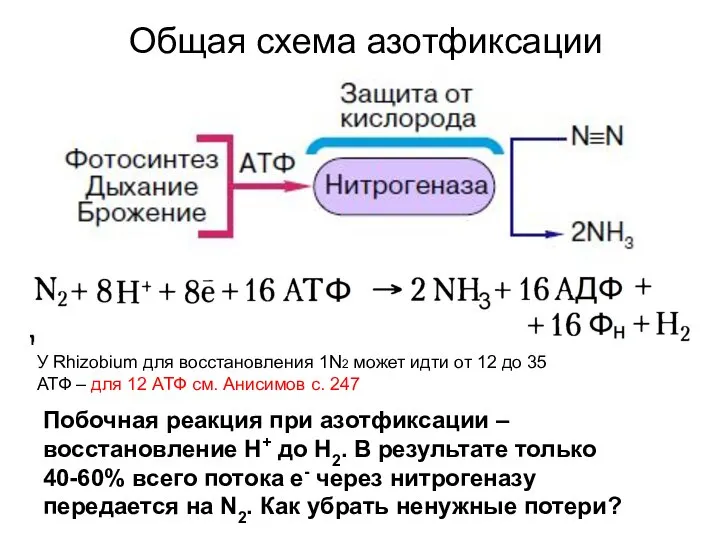
Слайд 10Общая схема азотфиксации
У Rhizobium для восстановления 1N2 может идти от 12 до
Общая схема азотфиксации
У Rhizobium для восстановления 1N2 может идти от 12 до
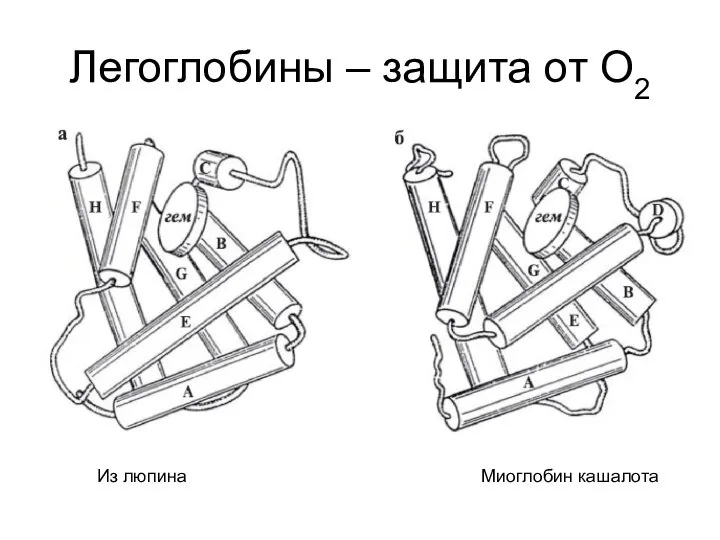
Слайд 11Легоглобины – защита от О2
Из люпина
Миоглобин кашалота
Легоглобины – защита от О2
Из люпина
Миоглобин кашалота
Слайд 12“Из аммиака в аммонийную форму” - работает нитрогеназный комплекс (упрощенно называемый нитрогеназой).
Комплекс
“Из аммиака в аммонийную форму” - работает нитрогеназный комплекс (упрощенно называемый нитрогеназой).
Комплекс

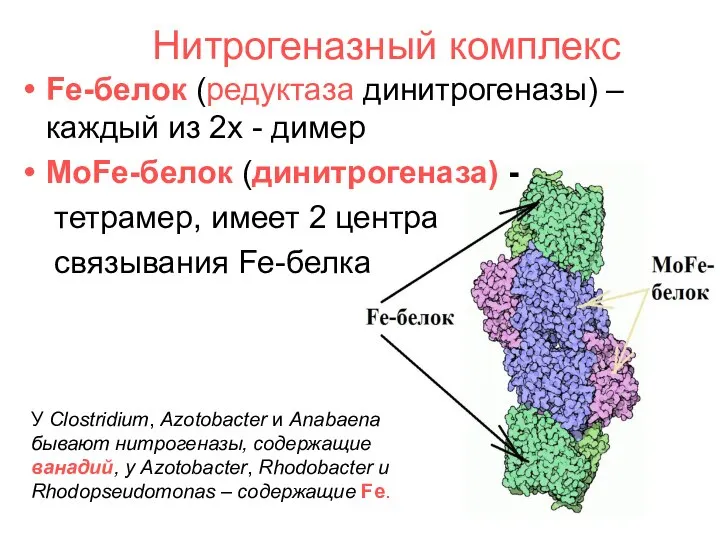
Слайд 13Нитрогеназный комплекс
Fe-белок (редуктаза динитрогеназы) – каждый из 2х - димер
MoFe-белок (динитрогеназа) -
Нитрогеназный комплекс
Fe-белок (редуктаза динитрогеназы) – каждый из 2х - димер
MoFe-белок (динитрогеназа) -
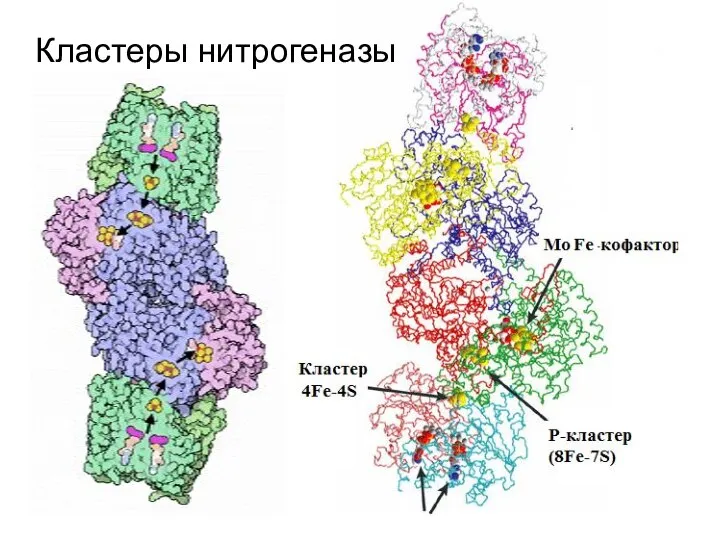
Слайд 14Кластеры нитрогеназы
Кластеры нитрогеназы
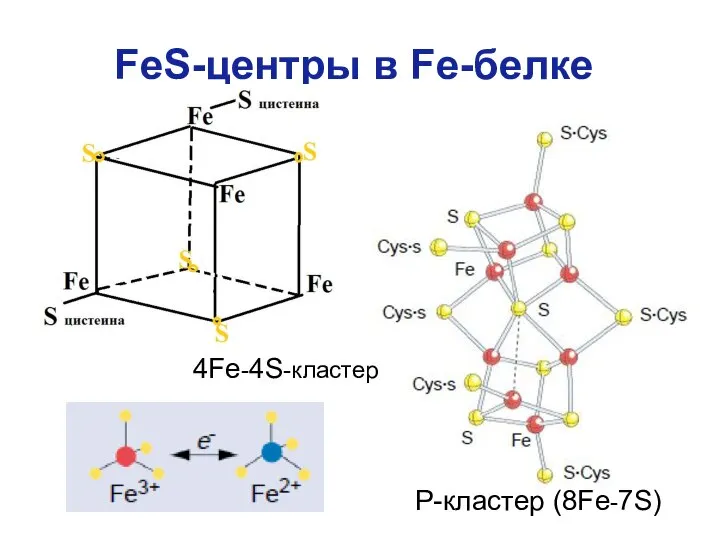
Слайд 15FeS-центры в Fe-белке
Р-кластер (8Fe-7S)
4Fe-4S-кластер
FeS-центры в Fe-белке
Р-кластер (8Fe-7S)
4Fe-4S-кластер
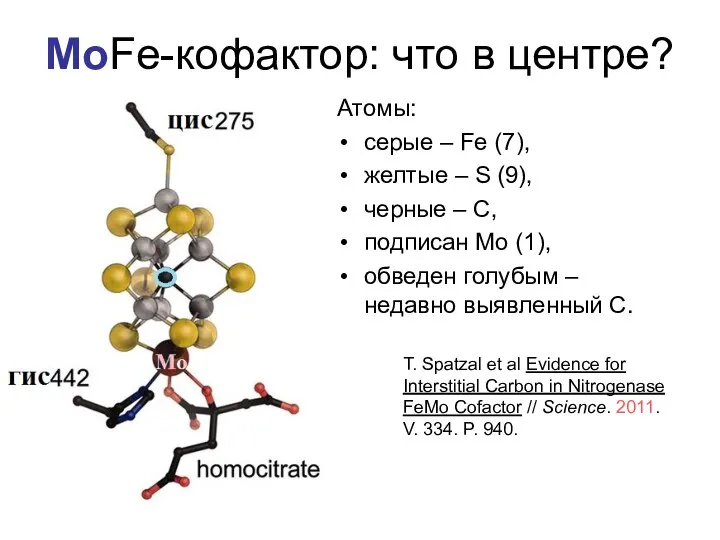
Слайд 16MoFe-кофактор: что в центре?
Атомы:
cерые – Fe (7),
желтые – S (9),
черные
MoFe-кофактор: что в центре?
Атомы:
cерые – Fe (7),
желтые – S (9),
черные
Слайд 17Работа нитрогеназы
N2 + 8H+ + 8e + 16АТФ→ 2NH3 + H2 + 16АДФ + 16Фн
Работа нитрогеназы
N2 + 8H+ + 8e + 16АТФ→ 2NH3 + H2 + 16АДФ + 16Фн
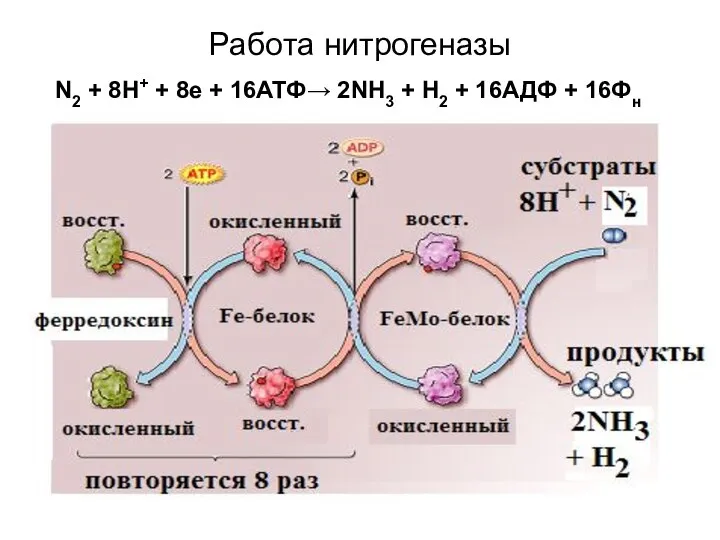
Слайд 18Расшифровка к предыдущему слайду
В молекуле Fe-белка (димере) 1 FeS-центр, поэтому Fe-белок может
Расшифровка к предыдущему слайду
В молекуле Fe-белка (димере) 1 FeS-центр, поэтому Fe-белок может

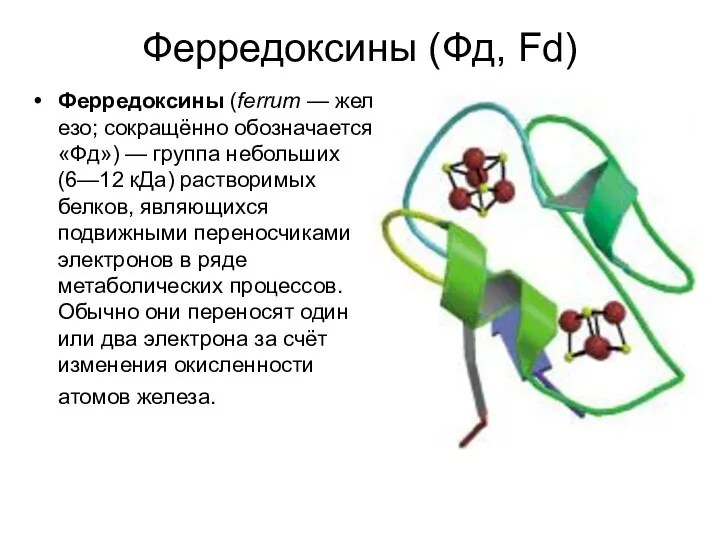
Слайд 19Ферредоксины (Фд, Fd)
Ферредоксины (ferrum — железо; сокращённо обозначается «Фд») — группа небольших (6—12 кДа) растворимых белков, являющихся
Ферредоксины (Фд, Fd)
Ферредоксины (ferrum — железо; сокращённо обозначается «Фд») — группа небольших (6—12 кДа) растворимых белков, являющихся
Слайд 20Дополнительный материал
Направления исследований:
1) Разработка технологий для сельского хозяйства на базе изучения
Дополнительный материал
Направления исследований:
1) Разработка технологий для сельского хозяйства на базе изучения

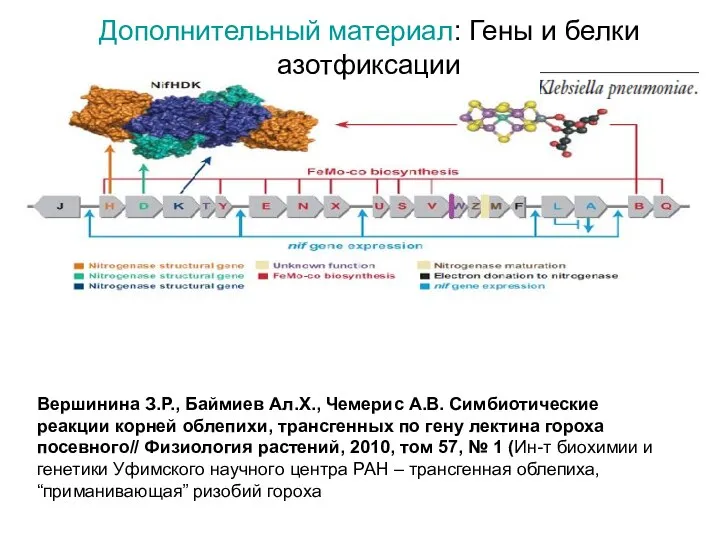
Слайд 21Дополнительный материал: Гены и белки азотфиксации
Вершинина З.Р., Баймиев Ал.Х., Чемерис А.В. Симбиотические
Дополнительный материал: Гены и белки азотфиксации
Вершинина З.Р., Баймиев Ал.Х., Чемерис А.В. Симбиотические
Слайд 22ПУТИ СИНТЕЗА АМИНОКИСЛОТ Семейства аминокислот по происхождению
ПУТИ СИНТЕЗА АМИНОКИСЛОТ Семейства аминокислот по происхождению

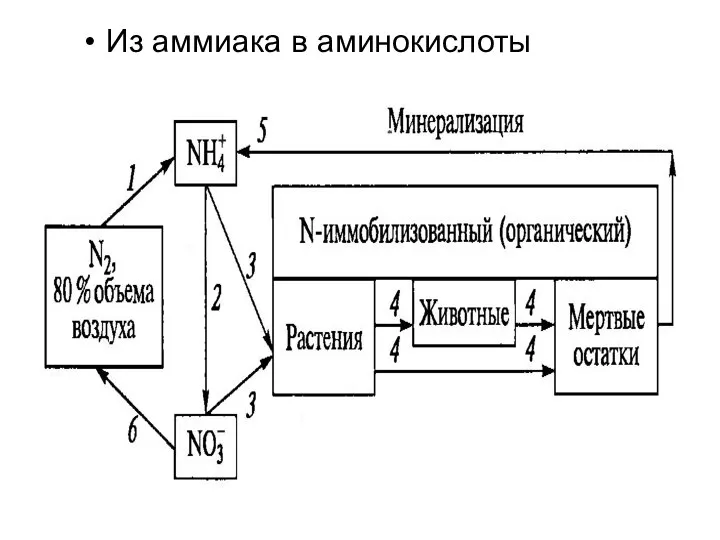
Слайд 23Из аммиака в аминокислоты
Из аммиака в аминокислоты
Слайд 24 РАСТЕНИЕ: поглощение и переработка NH4+ из почвы (или от симбионтов)
Дополнит. информация:2
РАСТЕНИЕ: поглощение и переработка NH4+ из почвы (или от симбионтов)
Дополнит. информация:2

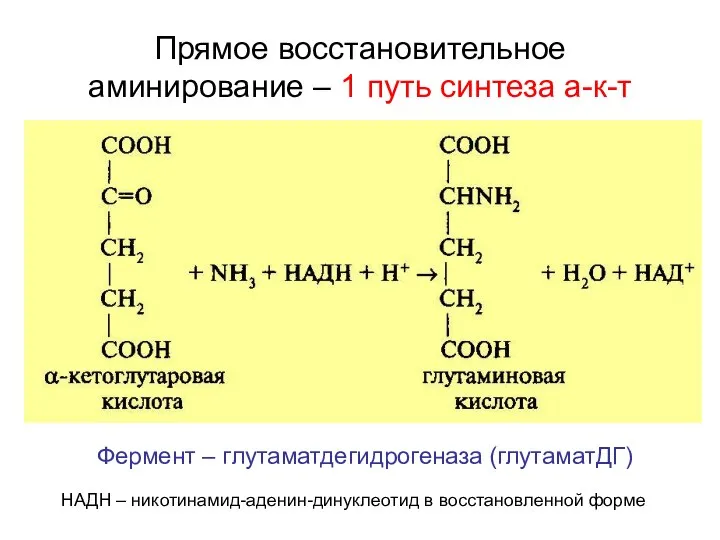
Слайд 25Прямое восстановительное аминирование – 1 путь синтеза а-к-т
Фермент – глутаматдегидрогеназа (глутаматДГ)
НАДН –
Прямое восстановительное аминирование – 1 путь синтеза а-к-т
Фермент – глутаматдегидрогеназа (глутаматДГ)
НАДН –
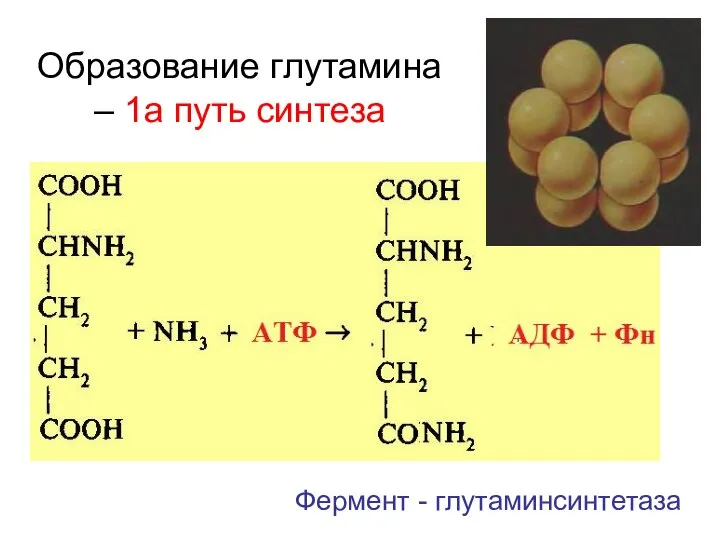
Слайд 26Образование глутамина – 1а путь синтеза
Фермент - глутаминсинтетаза
Образование глутамина – 1а путь синтеза
Фермент - глутаминсинтетаза
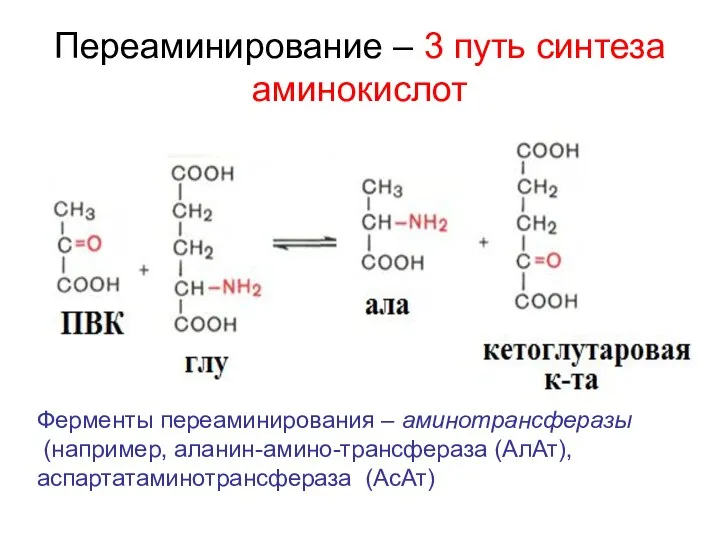
Слайд 27Переаминирование – 3 путь синтеза аминокислот
Ферменты переаминирования – аминотрансферазы
(например, аланин-амино-трансфераза (АлАт),
Переаминирование – 3 путь синтеза аминокислот
Ферменты переаминирования – аминотрансферазы
(например, аланин-амино-трансфераза (АлАт),
Слайд 283 путь синтеза аминокислот -
ферментативные превращения
1) Семейства аминокислот 1 – 4 (на
3 путь синтеза аминокислот -
ферментативные превращения
1) Семейства аминокислот 1 – 4 (на

Слайд 29Семейства аминокислот 1-4
Семейства аминокислот 1-4
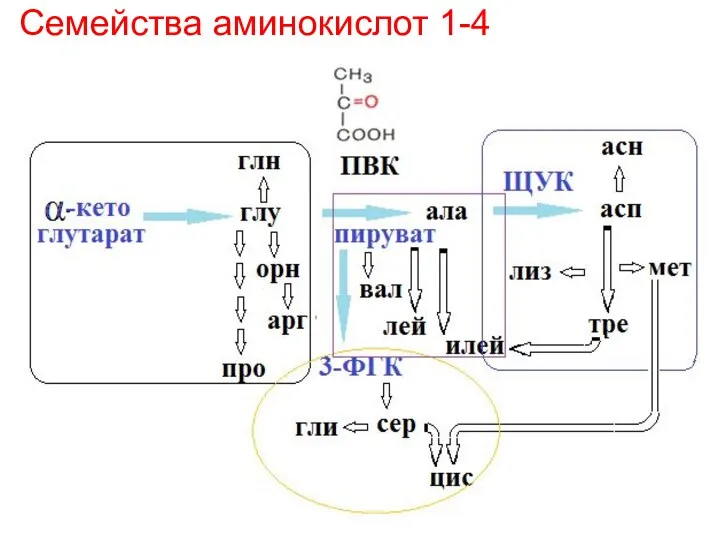
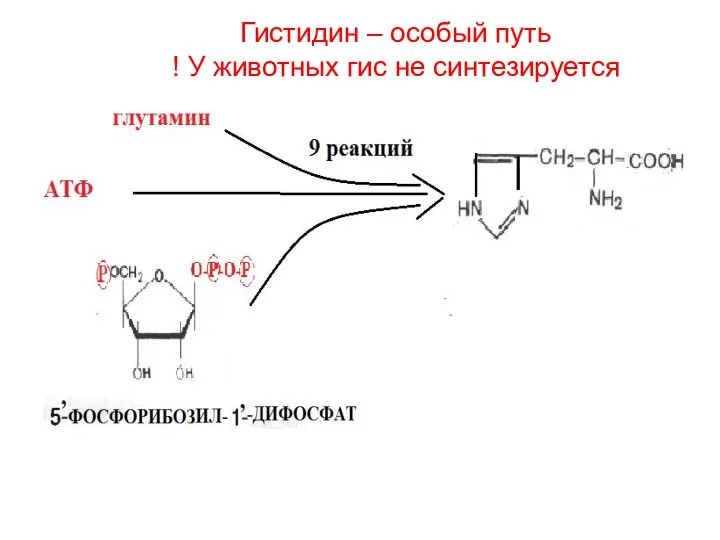
Слайд 30Гистидин – особый путь
! У животных гис не синтезируется
Гистидин – особый путь
! У животных гис не синтезируется
Слайд 31Семейство 5 (ароматические) – по шикиматному пути
1) синтез дегидрохинной кислоты (дегидрохинната), у
Семейство 5 (ароматические) – по шикиматному пути
1) синтез дегидрохинной кислоты (дегидрохинната), у

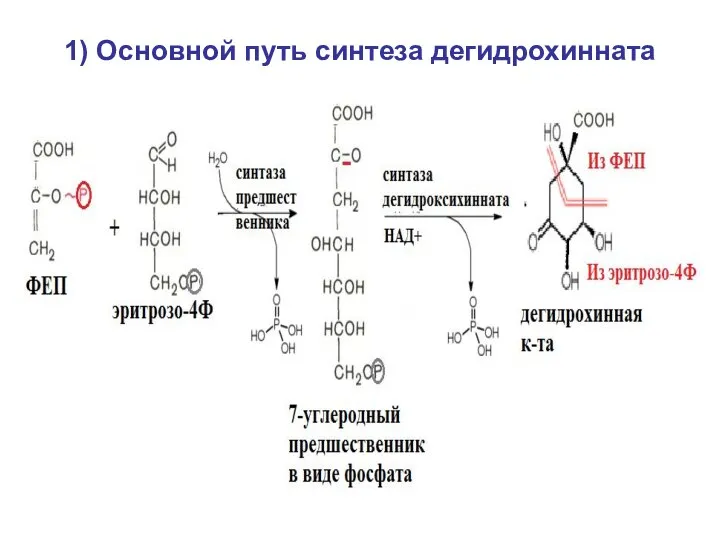
Слайд 321) Основной путь синтеза дегидрохинната
1) Основной путь синтеза дегидрохинната
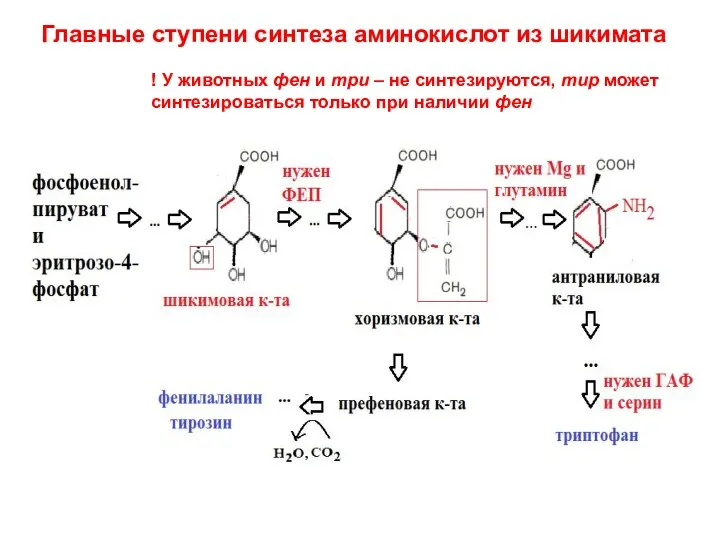
Слайд 33Главные ступени синтеза аминокислот из шикимата
! У животных фен и три –
Главные ступени синтеза аминокислот из шикимата
! У животных фен и три –
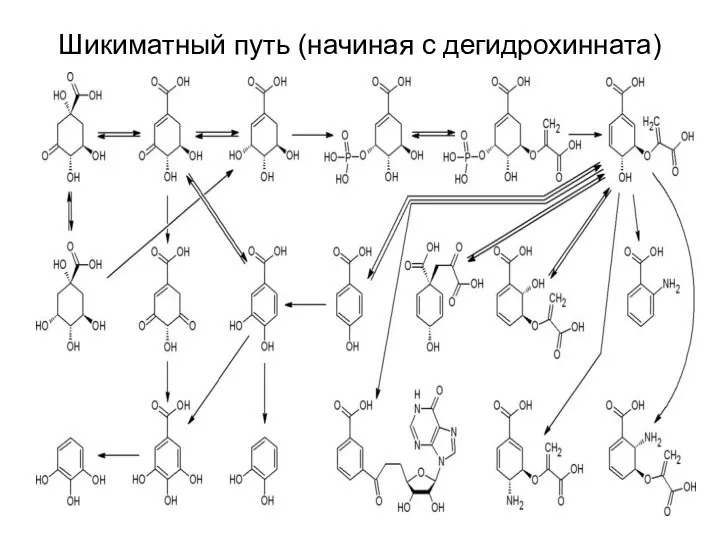
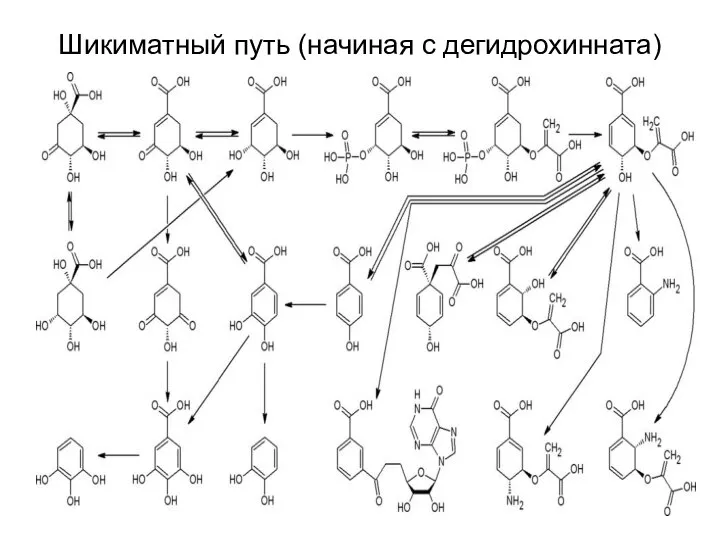
Слайд 34Шикиматный путь (начиная с дегидрохинната)
Шикиматный путь (начиная с дегидрохинната)
Слайд 35ИТОГО: основные пути синтеза белковых аминокислот
1) прямое восстановительное аминирование
1а) –
ИТОГО: основные пути синтеза белковых аминокислот
1) прямое восстановительное аминирование
1а) –

Слайд 36Ферментативные превращения в синтезе аминокислот
1) Семейства аминокислот 1 – 4 (на основе
Ферментативные превращения в синтезе аминокислот
1) Семейства аминокислот 1 – 4 (на основе

Слайд 37Семейства аминокислот 1-4
Семейства аминокислот 1-4
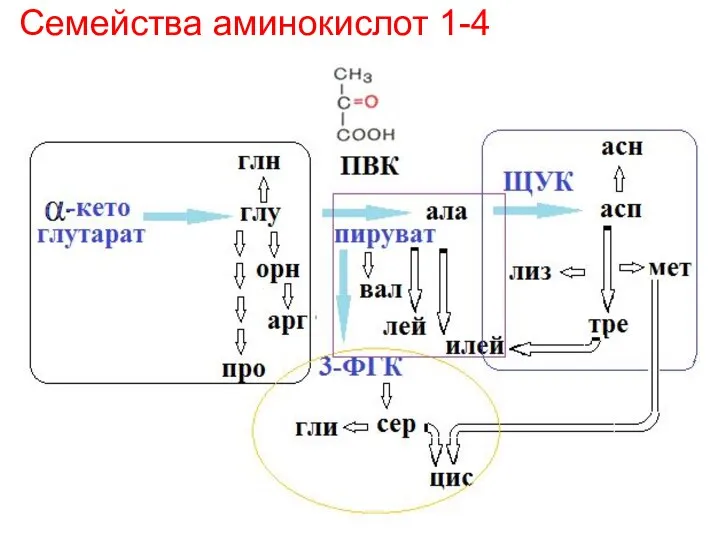
Слайд 38Гистидин – особый путь
! У животных гис не синтезируется
Гистидин – особый путь
! У животных гис не синтезируется
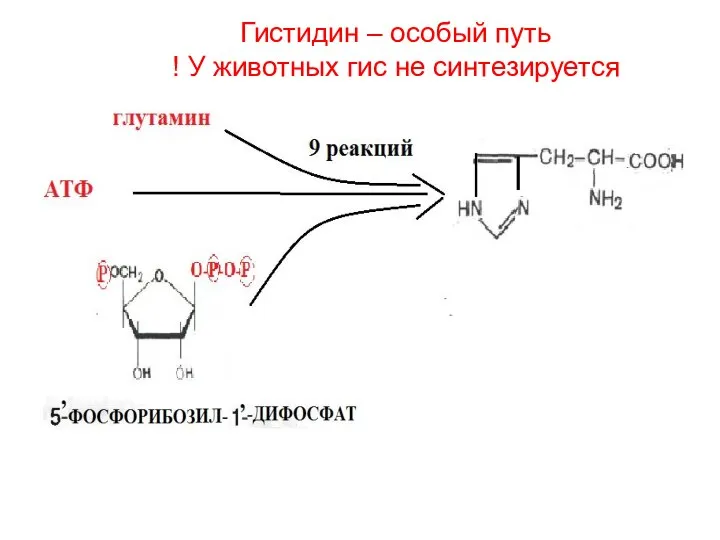
Слайд 39Семейство 5 (ароматические) – по шикиматному пути
1) синтез дегидрохинной кислоты (дегидрохинната), у
Семейство 5 (ароматические) – по шикиматному пути
1) синтез дегидрохинной кислоты (дегидрохинната), у

Слайд 401) Основной путь синтеза дегидрохинната
1) Основной путь синтеза дегидрохинната
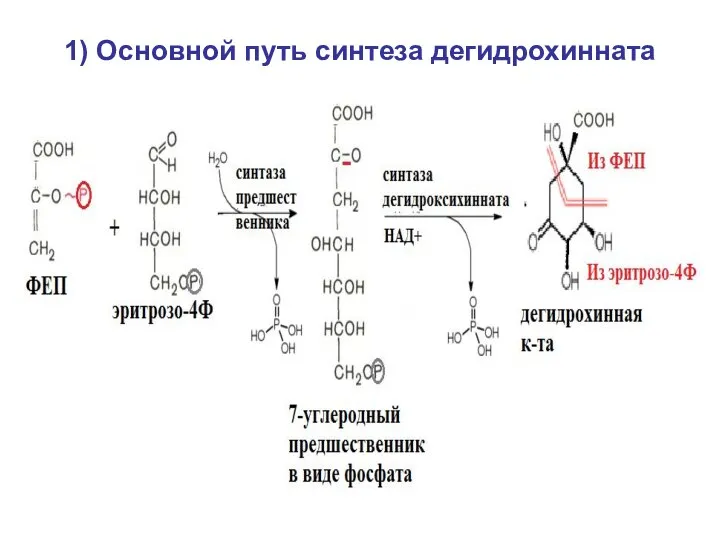
Слайд 41Главные ступени синтеза аминокислот из шикимата
! У животных фен и три –
Главные ступени синтеза аминокислот из шикимата
! У животных фен и три –
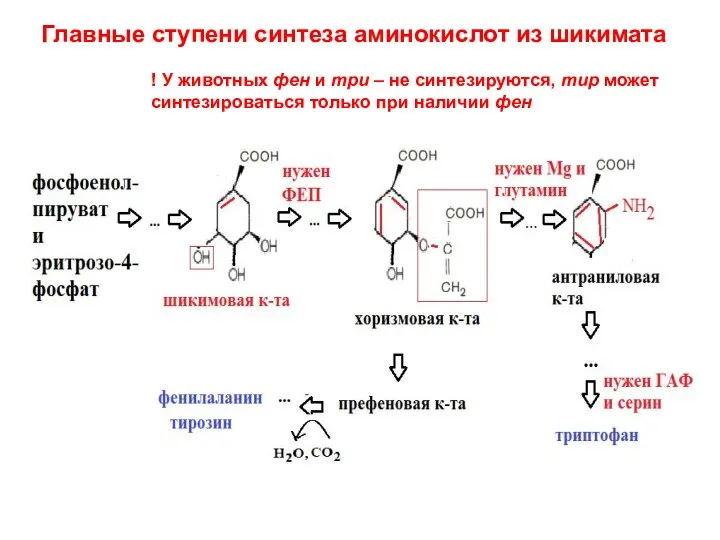
Слайд 42Шикиматный путь (начиная с дегидрохинната)
Шикиматный путь (начиная с дегидрохинната)
Слайд 43Нитрификация:
1) автотрофная (бактерии сем. Nitrobacteriaceae, археи), при окислении аммиака получающие энергию в
Нитрификация:
1) автотрофная (бактерии сем. Nitrobacteriaceae, археи), при окислении аммиака получающие энергию в

Слайд 44Автотрофная нитрификация у бактерий
a) NH4+ в клетку – транспортный белок Сu-содержащая транслоказа
б)
Автотрофная нитрификация у бактерий
a) NH4+ в клетку – транспортный белок Сu-содержащая транслоказа
б)

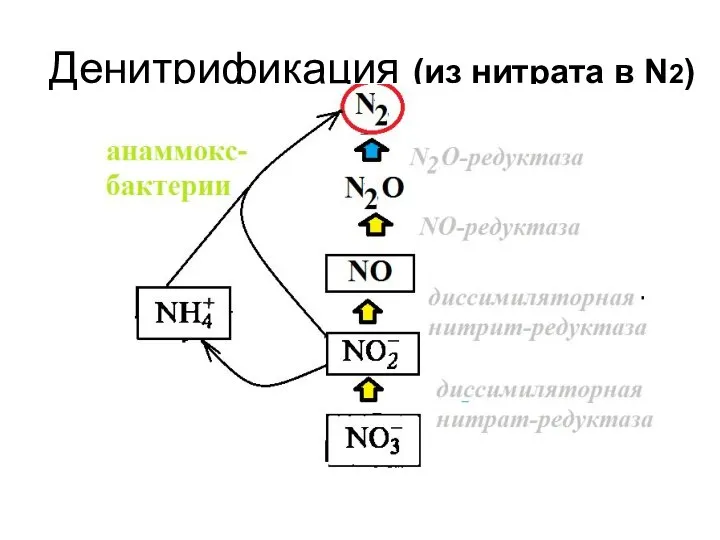
Слайд 45Денитрификация (из нитрата в N2)
Денитрификация (из нитрата в N2)
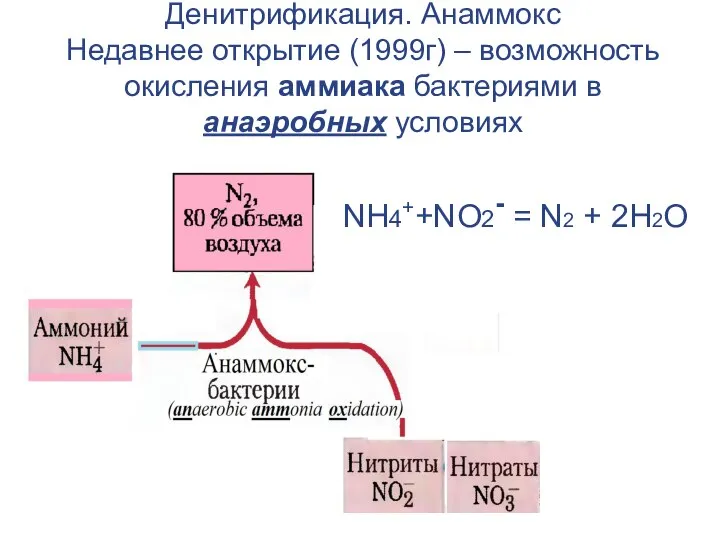
Слайд 46Денитрификация. Анаммокс
Недавнее открытие (1999г) – возможность окисления аммиака бактериями в анаэробных условиях
NH4++NO2-
Денитрификация. Анаммокс
Недавнее открытие (1999г) – возможность окисления аммиака бактериями в анаэробных условиях
NH4++NO2-
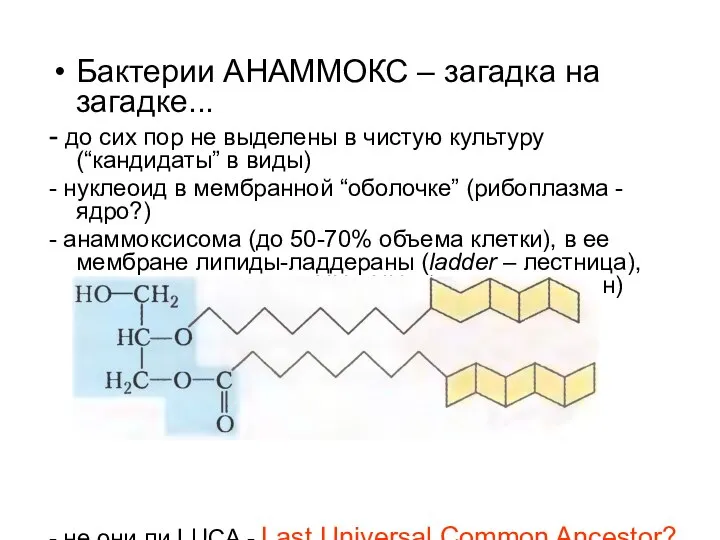
Слайд 47Бактерии АНАММОКС – загадка на загадке...
- до сих пор не выделены в
Бактерии АНАММОКС – загадка на загадке...
- до сих пор не выделены в
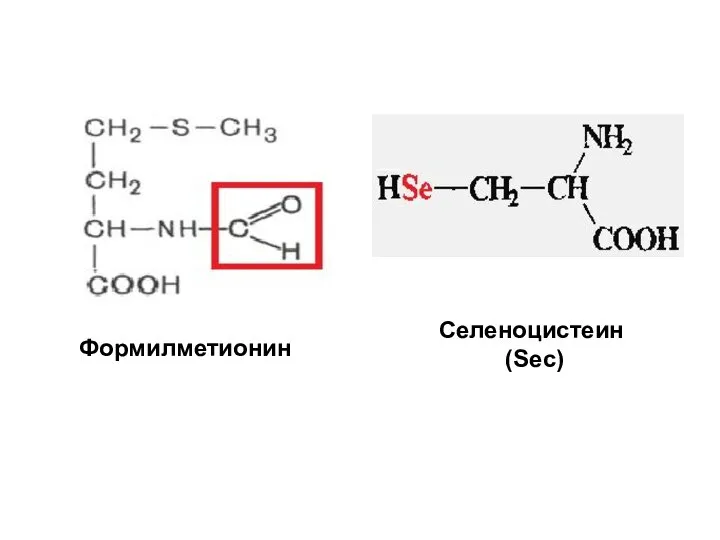
Слайд 48Селеноцистеин
(Sec)
Формилметионин
Селеноцистеин
(Sec)
Формилметионин
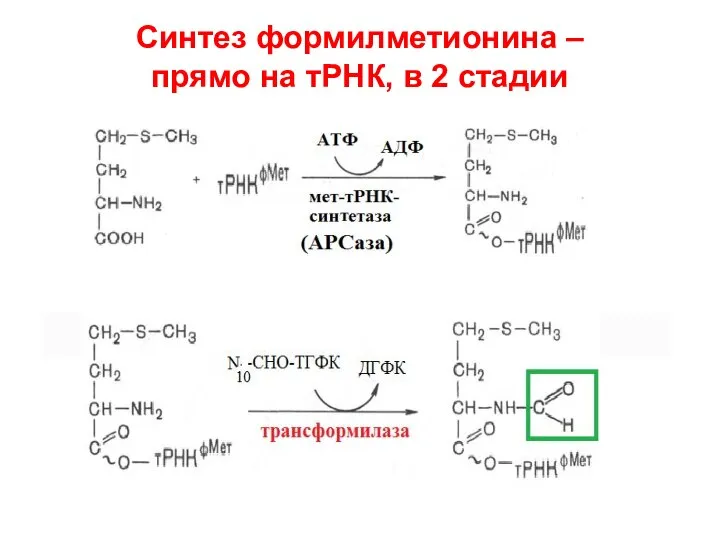
Слайд 49Синтез формилметионина –
прямо на тРНК, в 2 стадии
Синтез формилметионина –
прямо на тРНК, в 2 стадии
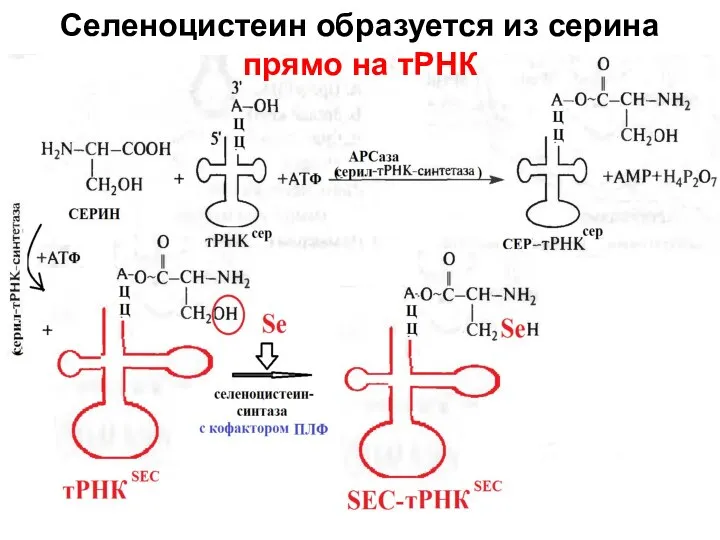
Слайд 50Селеноцистеин образуется из серина прямо на тРНК
Селеноцистеин образуется из серина прямо на тРНК
Слайд 51γ-аминомасляная кислота (ГАМК)
основной нейромедиатор торможения, рецепторы ГАМК A и ГАМК C – ионные
γ-аминомасляная кислота (ГАМК)
основной нейромедиатор торможения, рецепторы ГАМК A и ГАМК C – ионные

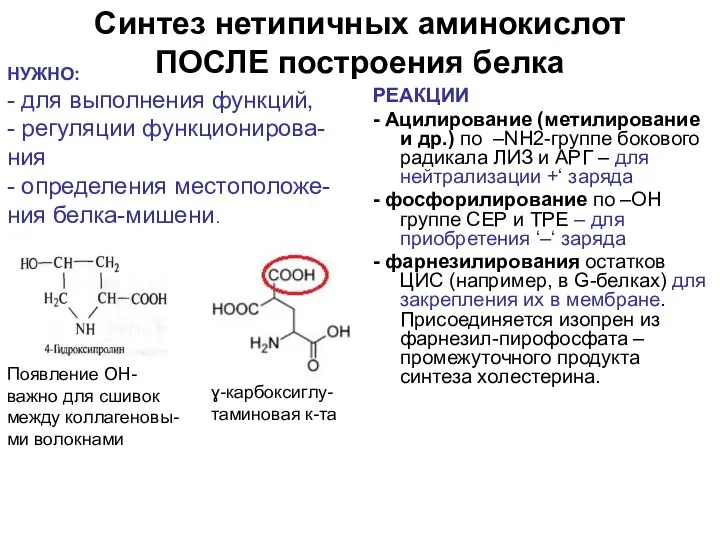
Слайд 52Синтез нетипичных аминокислот ПОСЛЕ построения белка
РЕАКЦИИ
- Ацилирование (метилирование и др.) по –NH2-группе
Синтез нетипичных аминокислот ПОСЛЕ построения белка
РЕАКЦИИ
- Ацилирование (метилирование и др.) по –NH2-группе
Слайд 53
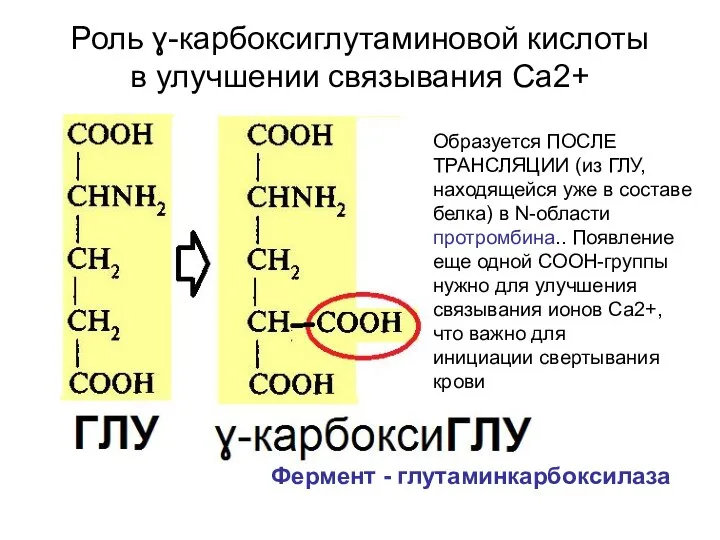
Роль ɣ-карбоксиглутаминовой кислоты
в улучшении связывания Са2+
Образуется ПОСЛЕ ТРАНСЛЯЦИИ (из ГЛУ,
Роль ɣ-карбоксиглутаминовой кислоты
в улучшении связывания Са2+
Образуется ПОСЛЕ ТРАНСЛЯЦИИ (из ГЛУ,
Слайд 54Mucuna (сем. Бобовые) – зудящие бобы
Гриффония (Griffonia simplicifolia, сем. Бобовые)
3,4-дигидроксифенилаланин
5-гидрокситриптофан
Ландыш, юкки
ЗАЩИТА.
Mucuna (сем. Бобовые) – зудящие бобы
Гриффония (Griffonia simplicifolia, сем. Бобовые)
3,4-дигидроксифенилаланин
5-гидрокситриптофан
Ландыш, юкки
ЗАЩИТА.

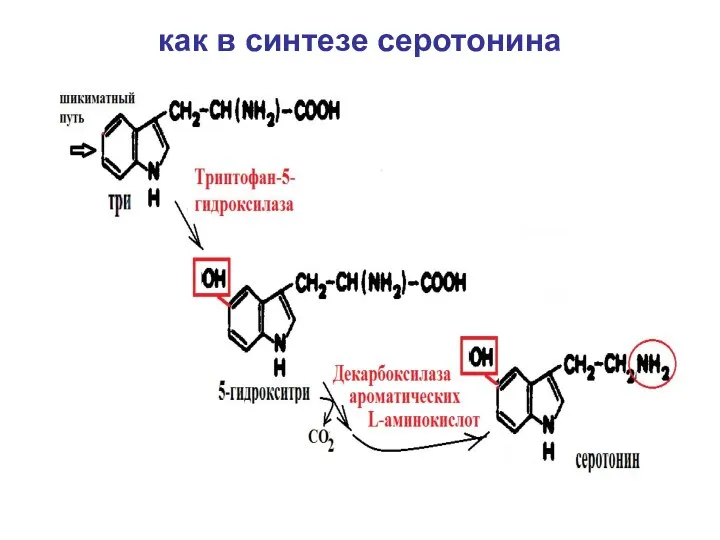
Слайд 55как в синтезе серотонина
как в синтезе серотонина
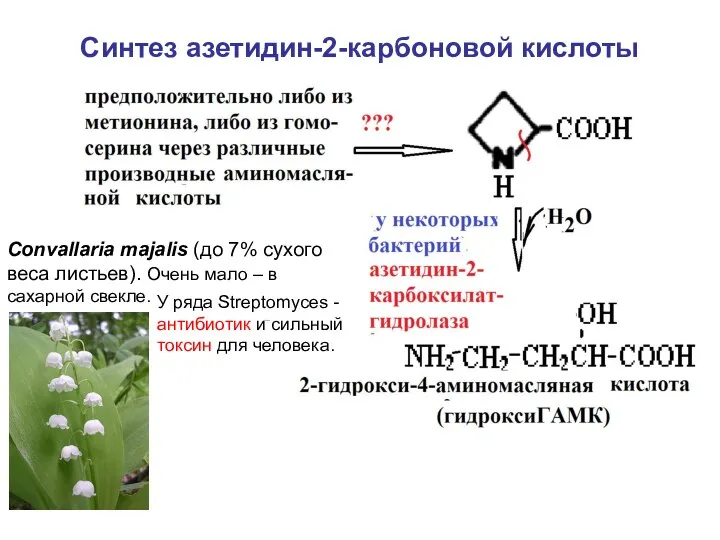
Слайд 56Синтез азетидин-2-карбоновой кислоты
Convallaria majalis (до 7% сухого веса листьев). Очень мало –
Синтез азетидин-2-карбоновой кислоты
Convallaria majalis (до 7% сухого веса листьев). Очень мало –
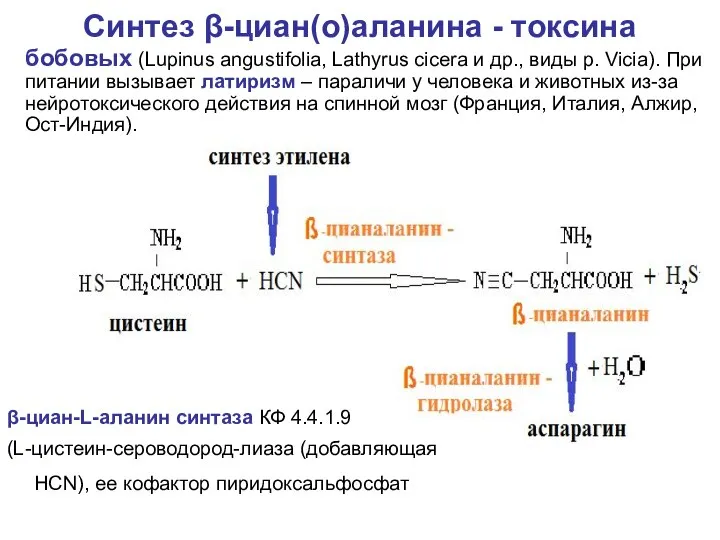
Слайд 57Синтез β-циан(о)аланина - токсина
β-циан-L-аланин синтаза КФ 4.4.1.9
(L-цистеин-сероводород-лиаза (добавляющая HCN), ее кофактор
Синтез β-циан(о)аланина - токсина
β-циан-L-аланин синтаза КФ 4.4.1.9
(L-цистеин-сероводород-лиаза (добавляющая HCN), ее кофактор
 Генетика. Закономерности наследования
Генетика. Закономерности наследования Биотехнологии. Биоинженерия
Биотехнологии. Биоинженерия soedineniya_kostey
soedineniya_kostey Обобщающий урок по теме: Жизнь растений
Обобщающий урок по теме: Жизнь растений Процесс пищеварения в желудке человека
Процесс пищеварения в желудке человека Презентация на тему "Энциклопедия слова - земляника" - презентации по Биологии
Презентация на тему "Энциклопедия слова - земляника" - презентации по Биологии Домики, которые построила природа
Домики, которые построила природа Человек – разумное существо
Человек – разумное существо Популяционная динамика бортевой пчелы (Apis mellifera mellifera L.) в заповеднике Шульган-Таш за полвека наблюдений
Популяционная динамика бортевой пчелы (Apis mellifera mellifera L.) в заповеднике Шульган-Таш за полвека наблюдений Кто такие птицы?
Кто такие птицы? Череп. Осевой скелет
Череп. Осевой скелет Анатомия волос
Анатомия волос Різноманітність аеробіонтів
Різноманітність аеробіонтів Атипус стенной
Атипус стенной Головной мозг. Нейрон
Головной мозг. Нейрон Презентация на тему Гипертония
Презентация на тему Гипертония  Кузнечик. Анатомия
Кузнечик. Анатомия Папоротники. Виды папоротника, размножение
Папоротники. Виды папоротника, размножение Токсины растений. Часть 2
Токсины растений. Часть 2 Презентация на тему Лишайники – симбиотические организмы
Презентация на тему Лишайники – симбиотические организмы  Морфология микроорганизмов
Морфология микроорганизмов Биомеханика кошки
Биомеханика кошки Классификация живых организмов. Основные систематические категории
Классификация живых организмов. Основные систематические категории Интересные факты про органы чувств человека
Интересные факты про органы чувств человека Тип Членистоногие
Тип Членистоногие 3 закона Г.И. Менделя
3 закона Г.И. Менделя Класс Пресмыкающиеся, или Рептилии
Класс Пресмыкающиеся, или Рептилии Розвиток пташенят від яйця до дорослої особини
Розвиток пташенят від яйця до дорослої особини