- Особенности обмена отдельных аминокислот

Содержание
- 2. Функции цистеина - участие в фолдинге белков за счет способности тиогруппы цистеина образовывать дисульфидные связи. При
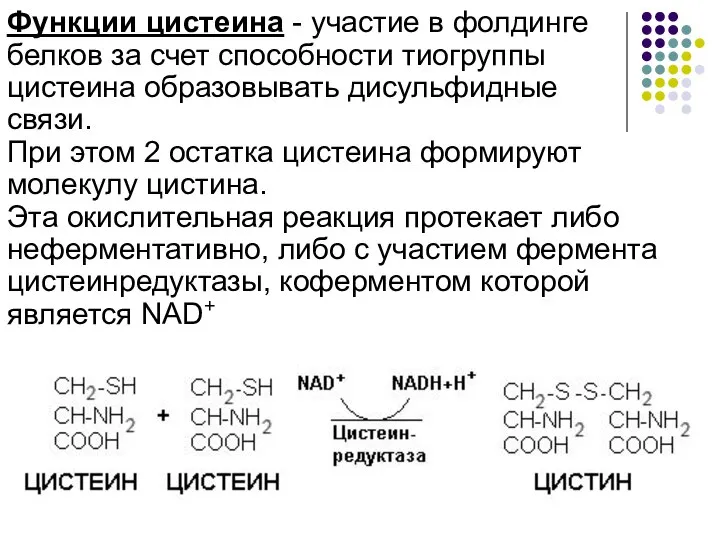
- 3. Нарушения обмена ЦИСТЕИНА Образование гомоцистина при нарушении использования гомоцистеина Гомоцистин накапливается в крови и в тканях,
- 4. Дисульфидные связи стабилизируют пространственную структуру полипептидной цепи или связывают между собой 2 цепи (например:А и В-цепи
- 5. Участие глутатиона в восстановлении цистина Осуществление антиоксидантной функции Формирование надвторичной структур Участие в фолдинге белка
- 6. СИНТЕЗ ТАУРИНА - важный путь использования цистеина, который осуществляется за счет декарбоксилирования производных цистеина - цистеиновой
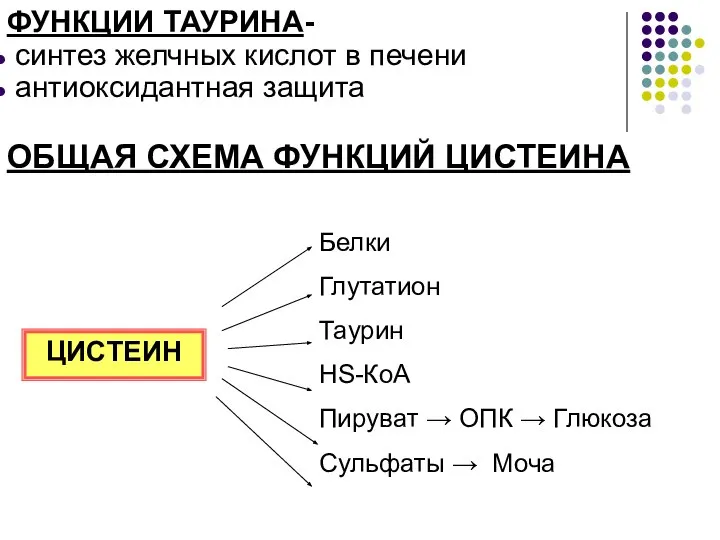
- 7. ФУНКЦИИ ТАУРИНА- синтез желчных кислот в печени антиоксидантная защита ОБЩАЯ СХЕМА ФУНКЦИЙ ЦИСТЕИНА ЦИСТЕИН Белки Глутатион
- 8. МЕТИОНИН - незаменимая АМК, однако она может регенерироваться из гомоцистеина. Следовательно, незаменим именно гомоцистеин, но единственным
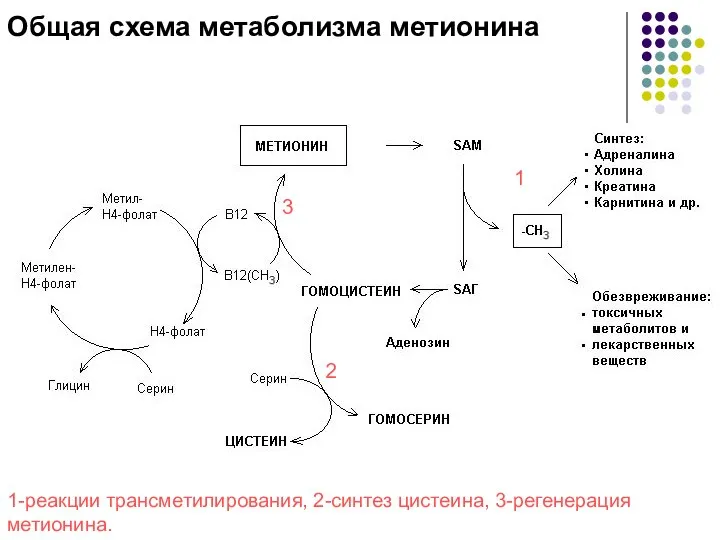
- 9. Общая схема метаболизма метионина 1-реакции трансметилирования, 2-синтез цистеина, 3-регенерация метионина. 1 3 2
- 10. Метаболизм ФЕНИЛАЛАНИНА 2 основных пути: включение в белки и превращение в тирозин Тирозин - заменимая АМК,
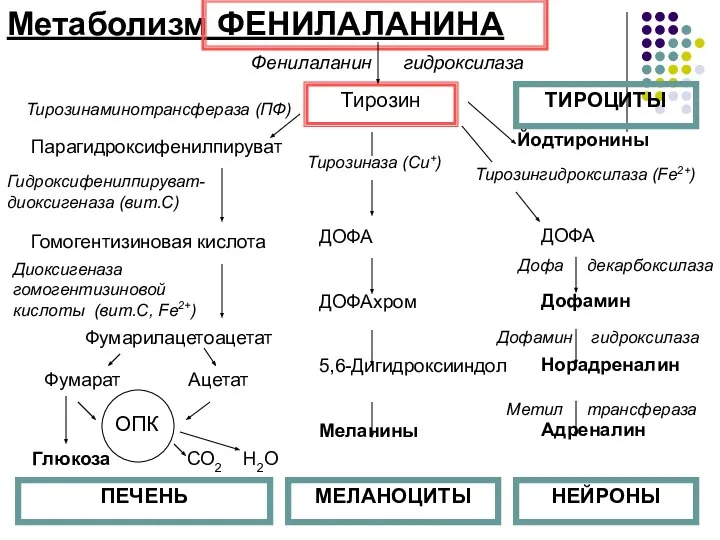
- 11. Метаболизм ФЕНИЛАЛАНИНА Тирозин Фенилаланин гидроксилаза Тирозинаминотрансфераза (ПФ) Парагидроксифенилпируват Гомогентизиновая кислота Гидроксифенилпируват-диоксигеназа (вит.С) Диоксигеназа гомогентизиновой кислоты (вит.С,
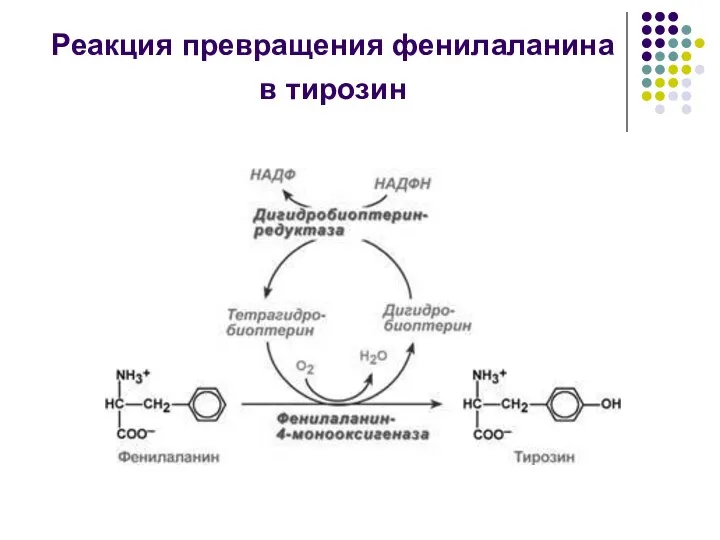
- 12. Реакция превращения фенилаланина в тирозин
- 13. Обмен ФЕНИЛАЛАНИНА и ТИРОЗИНА связан со значительным количеством реакций гидроксилирования, катализируемых оксигеназами (гидроксилазами), использующими молекулу О2
- 14. Схема превращение фенилаланина при фенилкетонурии При любых нарушениях превращения его в тирозин развивается фенилкетонурия.
- 15. В патогенезе фенилкетонурии имеют значение многие обстоятельства, в частности: 1) значительное накопление в тканях и жидкостях
- 16. Обмен АРГИНИНА Аргинин - источник оксида азота (NO, ЭРФ) и орнитина (АМК, не входящей в состав
- 17. выступает как вторичный мессенджер и активирует цитозольную гуанилатциклазу, - является нейромедиатором, играет роль в регуляции сосудистого
- 18. (Вал, Лей, Илей) - не в печени, как у других АМК, а в мышцах, жировой ткани,
- 19. Катаболизм АМК с разветвленной цепью
- 20. Нарушение работы дегидрогеназного комплексе α-кислот лежит в основе заболеваний, связанных с нарушением метаболизма разветвленных аминокислот –
- 21. Обмен триптофана - незаменимая АМК. Триптофан под действием гемсодержащего фермента триптофан-2,3-диоксигеназы в присутствии молекулярного кислорода превращается
- 22. Обмен дикарбоновых АМК (глутаминовой и аспарагиновой) и их амидов (глутамина и аспарагина) Аспарагиновая кислота - участвует
- 24. Скачать презентацию
Слайд 3Нарушения обмена ЦИСТЕИНА
Образование гомоцистина при
нарушении использования гомоцистеина
Гомоцистин накапливается в крови и
Нарушения обмена ЦИСТЕИНА
Образование гомоцистина при
нарушении использования гомоцистеина
Гомоцистин накапливается в крови и
Слайд 4Дисульфидные связи стабилизируют пространственную структуру полипептидной
цепи или связывают между собой 2 цепи
Дисульфидные связи стабилизируют пространственную структуру полипептидной
цепи или связывают между собой 2 цепи

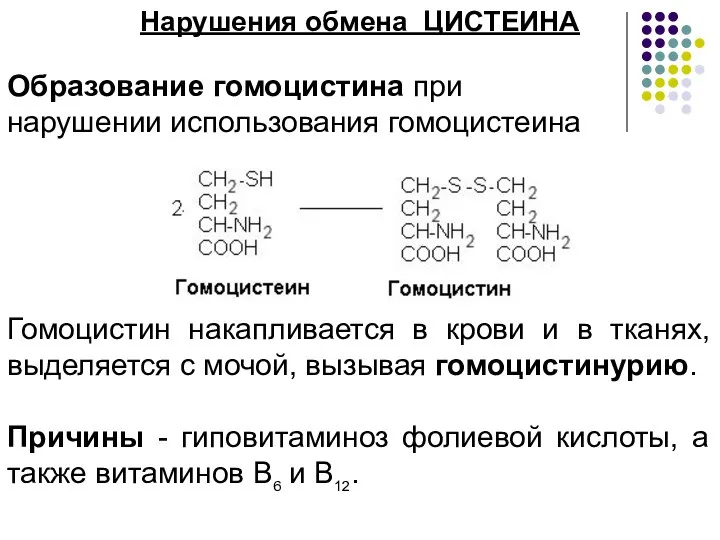
Слайд 5Участие глутатиона в восстановлении цистина
Осуществление антиоксидантной функции
Формирование надвторичной структур
Участие в фолдинге белка
Участие глутатиона в восстановлении цистина
Осуществление антиоксидантной функции
Формирование надвторичной структур
Участие в фолдинге белка
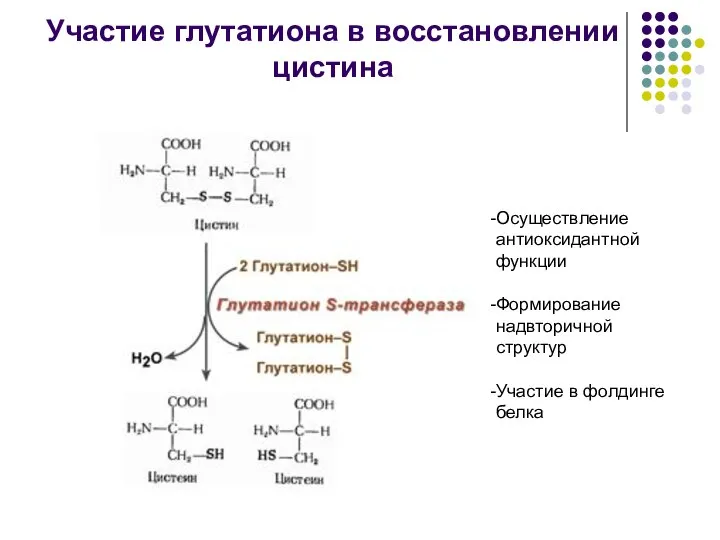
Слайд 6СИНТЕЗ ТАУРИНА - важный путь использования цистеина, который осуществляется за счет декарбоксилирования
СИНТЕЗ ТАУРИНА - важный путь использования цистеина, который осуществляется за счет декарбоксилирования
Слайд 7ФУНКЦИИ ТАУРИНА-
синтез желчных кислот в печени
антиоксидантная защита
ОБЩАЯ СХЕМА ФУНКЦИЙ
ФУНКЦИИ ТАУРИНА-
синтез желчных кислот в печени
антиоксидантная защита
ОБЩАЯ СХЕМА ФУНКЦИЙ
Слайд 8МЕТИОНИН - незаменимая АМК,
однако она может регенерироваться
из гомоцистеина.
Следовательно, незаменим именно
МЕТИОНИН - незаменимая АМК,
однако она может регенерироваться
из гомоцистеина.
Следовательно, незаменим именно

Слайд 9Общая схема метаболизма метионина
1-реакции трансметилирования, 2-синтез цистеина, 3-регенерация метионина.
1
3
2
Общая схема метаболизма метионина
1-реакции трансметилирования, 2-синтез цистеина, 3-регенерация метионина.
1
3
2
Слайд 10Метаболизм ФЕНИЛАЛАНИНА
2 основных пути: включение в белки
и превращение в тирозин
Тирозин -
Метаболизм ФЕНИЛАЛАНИНА
2 основных пути: включение в белки
и превращение в тирозин
Тирозин -

Слайд 11Метаболизм ФЕНИЛАЛАНИНА
Тирозин
Фенилаланин гидроксилаза
Тирозинаминотрансфераза (ПФ)
Парагидроксифенилпируват
Гомогентизиновая кислота
Гидроксифенилпируват-диоксигеназа (вит.С)
Диоксигеназа гомогентизиновой кислоты (вит.С, Fe2+)
Фумарилацетоацетат
Метаболизм ФЕНИЛАЛАНИНА
Тирозин
Фенилаланин гидроксилаза
Тирозинаминотрансфераза (ПФ)
Парагидроксифенилпируват
Гомогентизиновая кислота
Гидроксифенилпируват-диоксигеназа (вит.С)
Диоксигеназа гомогентизиновой кислоты (вит.С, Fe2+)
Фумарилацетоацетат
Слайд 12Реакция превращения фенилаланина в тирозин
Реакция превращения фенилаланина в тирозин
Слайд 13Обмен ФЕНИЛАЛАНИНА и ТИРОЗИНА
связан со значительным количеством реакций гидроксилирования, катализируемых оксигеназами
Обмен ФЕНИЛАЛАНИНА и ТИРОЗИНА
связан со значительным количеством реакций гидроксилирования, катализируемых оксигеназами

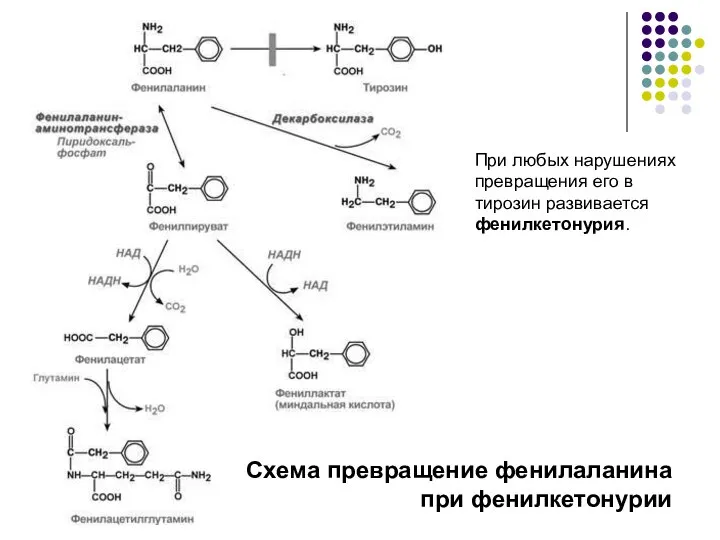
Слайд 14Схема превращение фенилаланина
при фенилкетонурии
При любых нарушениях превращения его в тирозин
Схема превращение фенилаланина
при фенилкетонурии
При любых нарушениях превращения его в тирозин
Слайд 15В патогенезе фенилкетонурии имеют значение многие
обстоятельства, в частности:
1) значительное накопление в тканях и
В патогенезе фенилкетонурии имеют значение многие
обстоятельства, в частности:
1) значительное накопление в тканях и

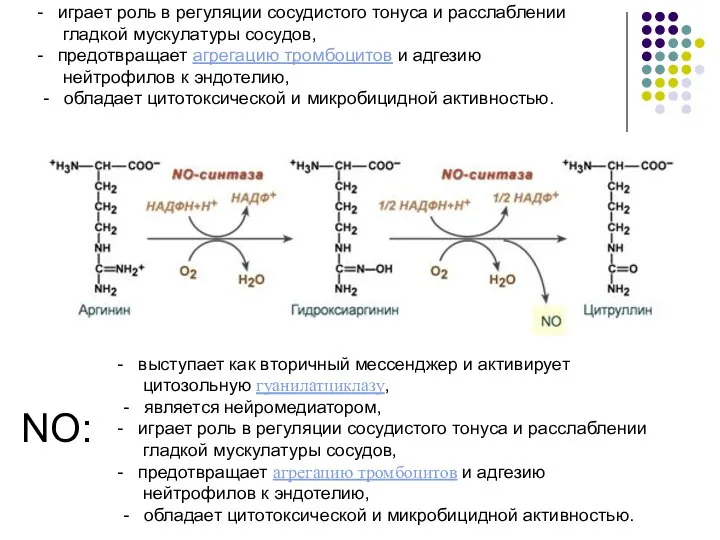
Слайд 16Обмен АРГИНИНА
Аргинин - источник оксида азота (NO, ЭРФ) и орнитина (АМК, не
Обмен АРГИНИНА
Аргинин - источник оксида азота (NO, ЭРФ) и орнитина (АМК, не

Слайд 17 выступает как вторичный мессенджер и активирует
цитозольную гуанилатциклазу,
- является нейромедиатором,
играет
выступает как вторичный мессенджер и активирует
цитозольную гуанилатциклазу,
- является нейромедиатором,
играет
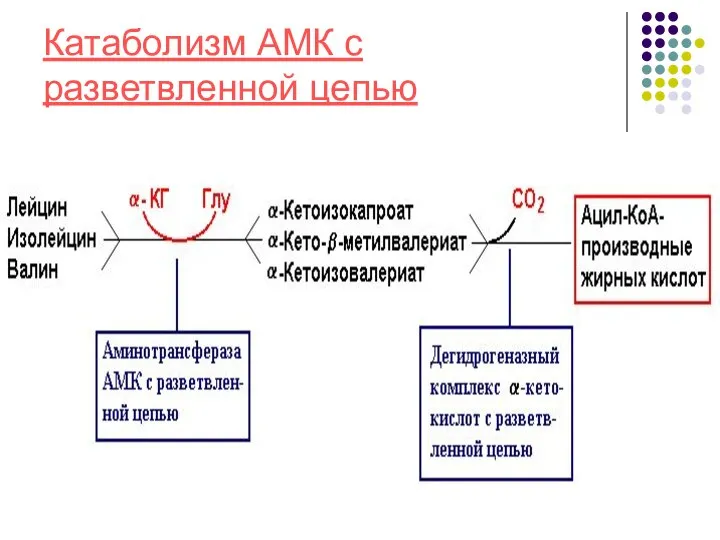
Слайд 18(Вал, Лей, Илей) - не в печени, как у других АМК, а
(Вал, Лей, Илей) - не в печени, как у других АМК, а

Слайд 19Катаболизм АМК с разветвленной цепью
Катаболизм АМК с разветвленной цепью
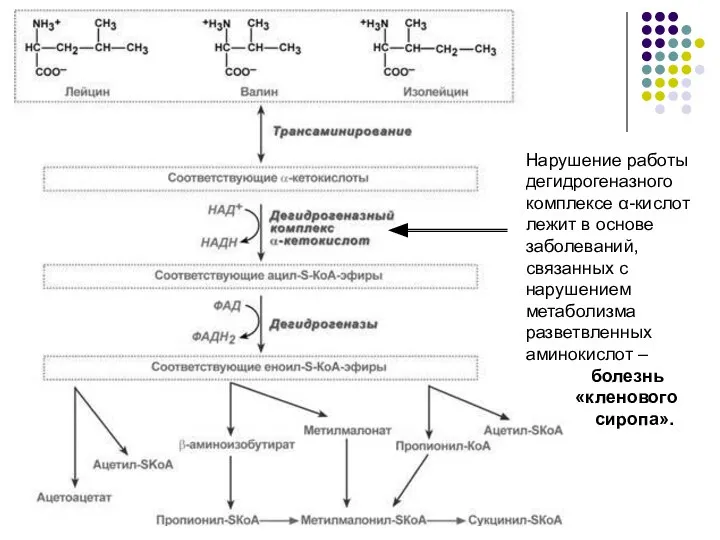
Слайд 20Нарушение работы дегидрогеназного комплексе α-кислот лежит в основе заболеваний, связанных с нарушением
Нарушение работы дегидрогеназного комплексе α-кислот лежит в основе заболеваний, связанных с нарушением
Слайд 21Обмен триптофана - незаменимая АМК.
Триптофан под действием гемсодержащего фермента триптофан-2,3-диоксигеназы в присутствии
Обмен триптофана - незаменимая АМК.
Триптофан под действием гемсодержащего фермента триптофан-2,3-диоксигеназы в присутствии

Слайд 22Обмен дикарбоновых АМК
(глутаминовой и аспарагиновой)
и их амидов (глутамина и аспарагина)
Аспарагиновая
Обмен дикарбоновых АМК
(глутаминовой и аспарагиновой)
и их амидов (глутамина и аспарагина)
Аспарагиновая

 Хемилюминесценция
Хемилюминесценция Теплица. Выращивание овощей
Теплица. Выращивание овощей Уголок живой природы. Экологическое занятие
Уголок живой природы. Экологическое занятие Широкопалый речной рак
Широкопалый речной рак В3 витамині суда, алкогольде, сірке қышқылында ерітілген майлы зат болып табылады
В3 витамині суда, алкогольде, сірке қышқылында ерітілген майлы зат болып табылады Что происходит с растениями зимой?
Что происходит с растениями зимой? Эфирные масла – как антисептик!
Эфирные масла – как антисептик! Строение плесневого гриба мукора
Строение плесневого гриба мукора Презентация на тему ЭВОЛЮЦИЯ ЧЕЛОВЕКА
Презентация на тему ЭВОЛЮЦИЯ ЧЕЛОВЕКА  Мышечная система
Мышечная система ПРОСТЕЙШИЕ
ПРОСТЕЙШИЕ Изучение и изготовление муравейника
Изучение и изготовление муравейника Chromosomal diseases associated with changes in the number of chromosomes in humans
Chromosomal diseases associated with changes in the number of chromosomes in humans Характеристика класса Marattiopsida
Характеристика класса Marattiopsida Почему звенит звонок
Почему звенит звонок Физиологическая роль натрия и калия в организме человека
Физиологическая роль натрия и калия в организме человека Семейство крестоцветные
Семейство крестоцветные Презентация на тему Космическая роль растений
Презентация на тему Космическая роль растений  Животные леса
Животные леса Роль корней в жизни и в питании растений
Роль корней в жизни и в питании растений Функции опорно-двигательной системы. 8 класс
Функции опорно-двигательной системы. 8 класс Презентация на тему Мы и наше здоровье. Организм человека
Презентация на тему Мы и наше здоровье. Организм человека  Углеводы (сахариды)
Углеводы (сахариды) Орган слуха и равновесия ухо
Орган слуха и равновесия ухо Кровоснабжение органов брюшной полости
Кровоснабжение органов брюшной полости Диаграмма цветка
Диаграмма цветка Растения в нашем уголке природы
Растения в нашем уголке природы Примитивные организмы
Примитивные организмы