Слайд 2Генетические основы методов создания селекционных популяций
На втором этапе селекционного процесса проводят отбор.

Под этим термином в селекции понимают, два мероприятия.
1. Сам отбор, т.е. оценку растений селекционного материала и выделение лучших из них в качестве родоначальников будущих потомств.
2. Применение разных методов получения от отобранных растений семенных или вегетативных потомств.
В процессе отбора на основе генофонда популяции исходного материала селекционер создает новые - сортовые популяции.
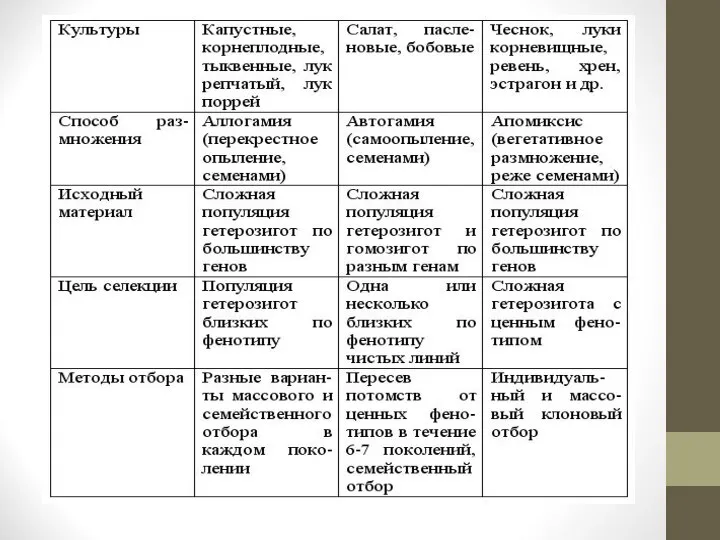
Структура сортовых популяций у разных видов зависит от способа размножения растений.
У перекрестноопыляемых растений они состоят из разнообразных по составу аллелей гетерозигот, имеющих близкое фенотипическое проявление генов.
Сортовые популяции самоопыляющихся растений представляют собой чистые линии или смесь очень близких чистых линий.
Сортовая популяция вегетативно размножаемых многолетних растений представлена клоном растения с очень сложной генетической природой, или смесью близких по фенотипу клонов.
Слайд 3Способ размножения растений определяет структуру создаваемой сортовой популяции и, в соответствии с

этим, существует три варианта выполнения второго этапа селекционного процесса:
1. При селекции перекрестников селекционер стремится, сохраняя гетерозиготность хозяйственно важных генов, достичь их фенотипической однородности.
При селекции самоопыляющихся растений, наоборот, необходимо возможно скорее достичь гомозиготности селектируемого материала, чтобы иметь возможность определить качество будущего сорта.
При селекции вегетативно размножаемых растений важно точно определить ценный генотип в исходном материале и после этого размножить его.
Три варианта выполнения второго этапа селекционного процесса, обусловленные способом размножения растений, можно кратко представить следующей таблицей.
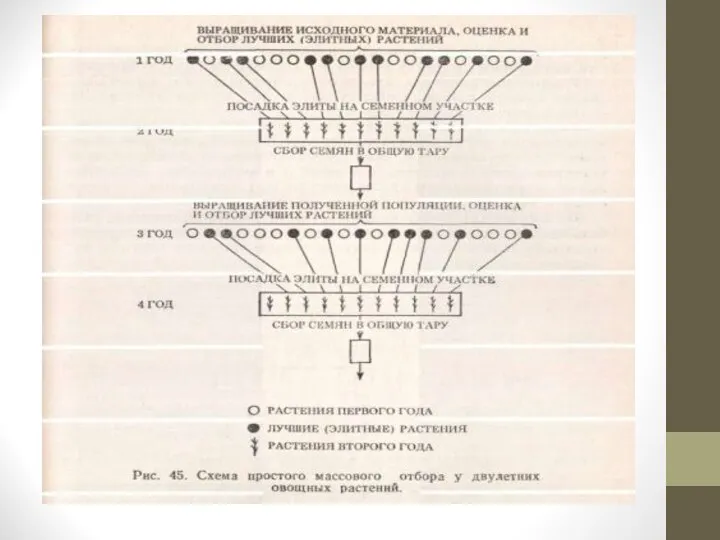
Слайд 5Древнейшим методом отбора является массовый.
При массовом отборе из исходной популяции отбирается большое

количество сходных по комплексу признаков лучших растений: от нескольких сотен до нескольких тысяч.
Семена объединяются и высеваются на следующий год на одной делянке. Следовательно, будущий сорт —это потомство массы лучших отобранных растений.
Массовый отбор может быть однократным и многократным (непрерывным).
Массовый однократный отбор заключается в том, что из общей массы растений отбирают самые лучшие по комплексу определенных признаков.
Необходимость в таком отборе возникает, например, при массовом засорении семян того или иного сорта, когда сортовая примесь настолько значительна, что удалять ее с помощью сортовой прополки нецелесообразно. Однократный массовый отбор более эффективен у самоопыляющихся культур.
Массовый многократный отбор, как правило, применяется у перекрестноопыляющихся культур.
Сущность этого метода в следующем. В первый год высевается исходный материал, из которого будет проводиться отбор.
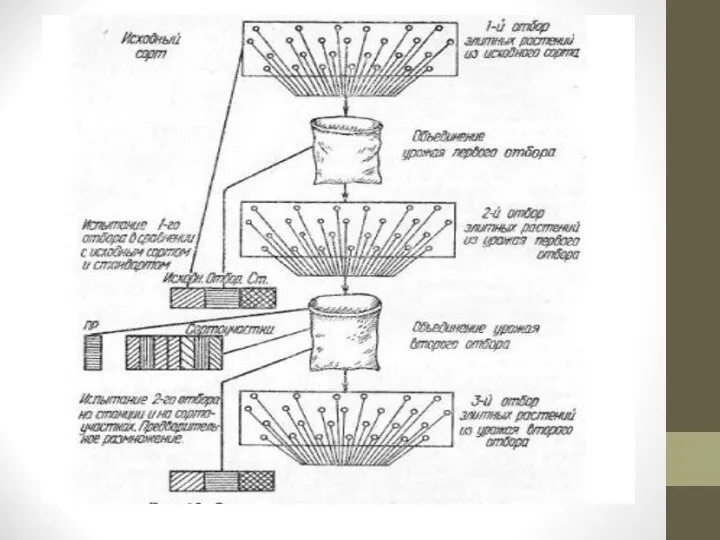
Слайд 7На участке отбираются элитные растения, в наибольшей степени отвечающие селекционной программе. Семена

с отобранных растений высеваются в будущем году на одном участке для повторного отбора. Часть семян урожая первого отбора высевается в сортоиспытании где результаты первого отбора сравнивают с исходной популяцией и лучшим стандартным сортом.
Собранные семена повторного массового отбора используются для посева с целью проведения третьего отбора и дальнейшего сортоиспытания. Если сортоиспытание дало положительные результаты, организуют предварительное размножение нового образца и испытание его на государственных сортоиспытательных участках.
Массовый многократный отбор (рис. 1) продолжают до тех пор, пока не будет получен новый сорт. Если для поддержания ценных признаков и свойств сорта необходим систематический отбор — его повторяют из года в год. Такой отбор называется непрерывным массовым отбором.
Этот метод используется также многими селекционными учреждениями для получение элитных семян перекрестноопыляющихся культур.
Слайд 9При использовании многократного массового отбора у вегетативно размножаемых растений полученный посадочный материал

в результате первого отбора поступает в селекционный питомник для последующего селекционного питомника и т. д. до тех пор, пока не будет получен новый улучшенный сорт.
Преимуществами массового отбора являются простота, доступность и возможность быстрого улучшения материала в значительном количестве.
Его можно использовать не только в научно-исследовательских учреждениях, но и в производственных условиях. Массовый отбор широко применяется в семеноводстве как метод сохранения однородности морфологических признаков и хозяйственно-биологических свойств выращиваемых сортов.
Одна из разновидностей его — негативный отбор, когда не отбирают лучшие растения, а удаляют из посева худшие. Негативный отбор также очень часто применяют в семеноводческой работе.
Слайд 10Массовый отбор дает хорошие и быстрые результаты в том случае, когда его

задачи соответствуют направлению естественного отбора.
Чем популяция экологически больше приспособлена к данным условиям возделывания в ходе естественного отбора, тем результативнее может быть массовый отбор.
Например, если в популяции имеются раннеспелые формы и этот признак в данной местности решающий для получения устойчивых урожаев, то массовый отбор на скороспелость может быть очень эффективным.
В селекции массовый отбор имеет большое значение. Все местные сорта народной селекции созданы этим методом.
Методом массового отбора получен на Шатиловской опытной станции сорт гречихи Богатырь. Созданный очень давно, он и в настоящее время является одним из лучших сортов этой культуры. Этим методом получены многие сорта ржи и почти все сорта многолетних трав.
Существенным недостатком массового отбора является то, что он не дает возможности индивидуально оценить потомство и выделить из популяции наиболее ценные в селекционном отношении формы.
Слайд 11Методы отбора, применяемые при селекции размножаемых семенами перекрестноопыляемых растений
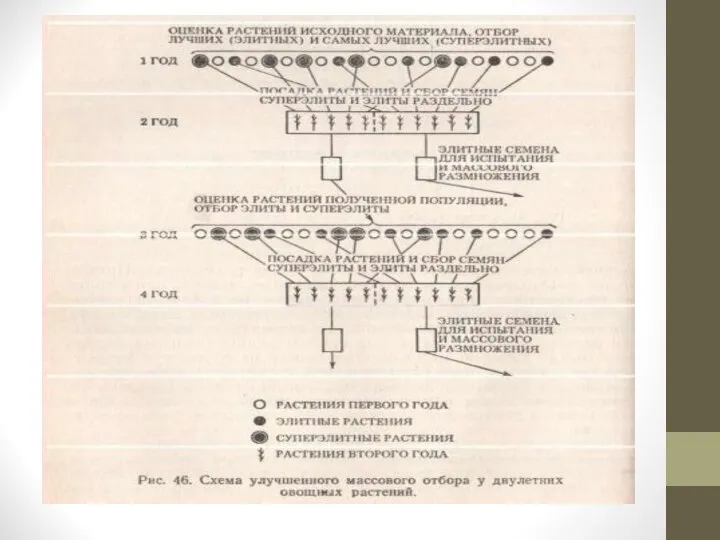
Применяют два варианта массового

отбора: простой и улучшенный, и четыре варианта семейственного отбора: без изоляции, с изоляцией, метод парных скрещиваний и метод половинок.
Простой массовый отбор. При простом массовом отборе из популяции исходного селекционного материала выделяют лучшие по фенотипу растения.
Если оценка товарного органа осуществляется до цветения, то маточные растения высаживают на отдельном изолированном участке, с целью их взаимного переопыления.
В зависимости от генетической природы исходного материала генотипы выделенных фенотипически ценных растений могут содержать как гетерозиготные, так и гомозиготные локусы, контролирующие ценные признаки.
Доля гомозигот в исходном материале представленном сортовыми популяциями перекрестно опыляемых растений незначительна. Чаще всего это касается генов, контролирующих качественные признаки.
Слайд 12В исходном материале ценные фенотипы преимущественно обусловлены гетерозиготностью их генотипов, или сверхдоминированием

аллелей.
При этом у разных генотипов могут быть самые разнообразные сочетания гетерозигот по разным аллелям разных локусов и разнообразные взаимодействия аллелей разных локусов.
При массовом отборе в результате взаимного переопыления сложного комплекса генотипов образуется новая сложная популяция следующего поколения.
В ней могут появиться как более ценные генотипы в результате новых удачных сочетаний положительных аллелей, так и не представляющие ценности в результате сочетания менее ценных аллелей.
В целом новая популяция будет более продвинута в направлении отбора в сравнении с популяцией исходного материала, если в качестве него были использованы сортовые популяции существующих местных или селекционных сортов.
Она будет резко отличаться сочетанием аллелей от исходной, если в качестве нее было использовано второе гибридное потомство.
Слайд 13При применении массового отбора к популяциям селекционных районированных, выращиваемых в других регионах

или зарубежных сортов выявленные в них перспективные фенотипы могут быть результатом, как особого сочетания ценных аллелей генов, так и модификационной изменчивости вследствие более благоприятных условий среды.
Поэтому при первом применении массового отбора следует выяснить степень наследуемости отбираемого признака.
Для этой цели используют такой показатель, как коэффициент наследуемости. Различают наследуемость в широком и в узком смысле слова.
Наследуемость в широком смысле обозначается символом Н2 и показывает долю генотипической изменчивости в фенотипической
Наследуемость в узком смысле– символом h2 и показывает долю изменчивости, контролируемую аддитивным действием генов, в общей фенотипической.
Слайд 14Коэффициент наследуемости в широком смысле определяется в виде отношения селекционного эффекта к

селекционному дифференциалу: Н2 = SE / SD.
Селекционный дифференциал (SD) вычисляют в год проведения первого отбора как разницу между средним значением признака у отобранных растений и средней величиной признака в исходной популяции.
Селекционный эффект (SE) – разница между средней величиной признака у полученного потомства и исходной популяции. Его определяют в следующем поколении.
Вычисленный таким способом коэффициент наследуемости при работе с перекрестниками отражает не только аддитивные эффекты полигенов, но и возникшие в созданной популяции внутри и меж генные взаимодействия аллелей.
Коэффициент наследуемости выше 0,6 считают достаточно надежным для эффективного проведения отбора.
Слайд 15У разных сортов реакция на новые провокационные условия окружающей среды бывает выражена

в разной степени: у одних реагируют все особи популяции, у других – только большая или меньшая часть.
Для селекционера представляют интерес генотипы, обеспечивающие получение товарного органа в условиях нового комплекса факторов среды.
Выделение новых генотипов из популяции и взаимное их опыление позволит создать новую популяцию, насыщенную аллелями с широкой нормой реакции.
В этой ситуации массовый отбор позволяет не только выделить из популяции генотипы с ценными сочетаниями аллелей генов, но и создать, а затем отобрать новые более эффективные их комбинации
Слайд 16Массовый отбор на олигогенный признак демонстрирует свою эффективность, если он контролируется рецессивными

аллелями генов.
Обнаружение в сортовой популяции особей, отличающихся от основной массы каким-то рецессивным признаком, наиболее вероятное событие. Как правило, это бывают спонтанные мутации, возникшие в каком-то из ранних поколений.
В силу своей рецессивности, они скрытно распространились в популяции в гетерозиготном состоянии и проявились в результате скрещивания гетерозигот.
Взаимное опыление нескольких отобранных особей с ценным признаком позволит получить однородное потомство.
При массовым отборе особей с доминантными признаками, они могут быть как гомозиготами, так и гетерозиготами по контролирующим их генам.
Их взаимное опыление снова даст потомство, содержащее доминантные гомозиготы, гетерозиготы и рецессивные гомозиготы. При удалении рецессивных гомозигот из селекционного материала с каждым поколением доля доминантных аллелей будет возрастать, но процесс идет медленно и зависит от доли гетерозигот среди особей, выделенных из исходной популяции.
Практически массовый отбор при отборе олигогенных доминантных признаков обладает низкой эффективностью.
Слайд 17Многие количественные признаки определяются не только генами ядра, но и цитоплазмы.
Генный

комплекс цитоплазмы может определять признак самостоятельно или во взаимодействии с генами ядра.
В первом случае включение в группу отбираемых родоначальников особей с цитоплазматическим контролем признака позволит быстро создать новую ценную сортовую популяцию
Во втором – процесс создания будет замедлен частичным разрушением существовавших комбинаций аллелей генов ядра и созданием новых, не всегда достаточно эффективных.
Эффективность массового отбора в значительной мере зависит от его интенсивности.
Интенсивность отбора, или доля выделяемых из исходной популяции родоначальников для получения популяции следующего поколения, определяется в первую очередь их наличием в исходной популяции, а не тем какую долю популяции они составляют.
Говорить об интенсивности отбора имеет смысл тогда, когда в разной мере ценные генотипы составляют значительную часть исходной популяции. Тогда высокая интенсивность отбора позволяет исключить из выделенного материала менее ценные генотипы.
Слайд 19Массовый улучшенный отбор отличается от простого массового отбора только большим числом оцениваемых

признаков или более тщательной оценкой особей.
В целом массовые отборы отличаются простотой выполнения, менее трудоемки в сравнении с другими методами, но вместе с тем и менее эффективны.
Их чаще применяют при выделении ценных фенотипов из исходной популяции и в последних поколениях второго этапа селекционного процесса при размножении созданных, близких к будущему сорту популяций.
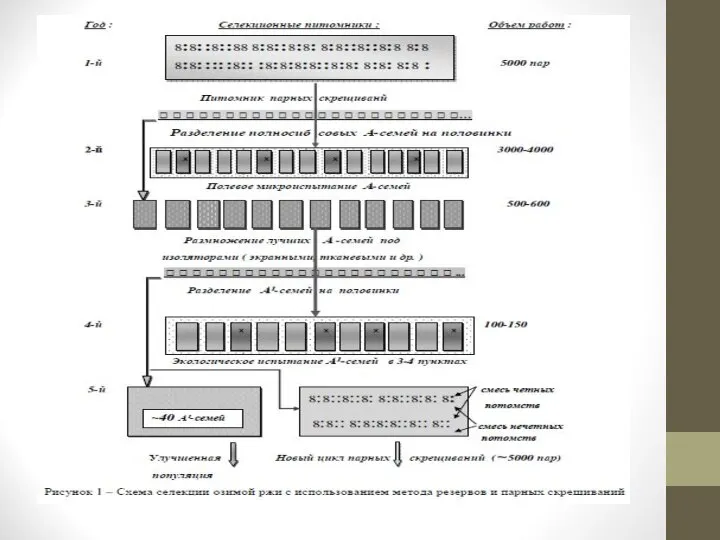
Семейственный отбор. Эффективность селекционного процесса с перекрестно опыляемыми растениями значительно возрастает при применении разных вариантов семейственного отбора.
При семейственном отборе семена с выделенных ценных растений собирают раздельно и полученные потомства (семьи) выращивают на отдельных делянках.
По результатам оценки растений получают оценку каждой семьи отдельно. Таким образом, материалом для отбора служат семьи. Из лучших семей выделяют маточные растения, с которых раздельно собирают семена и из них выращивают семьи следующего поколения и так далее.
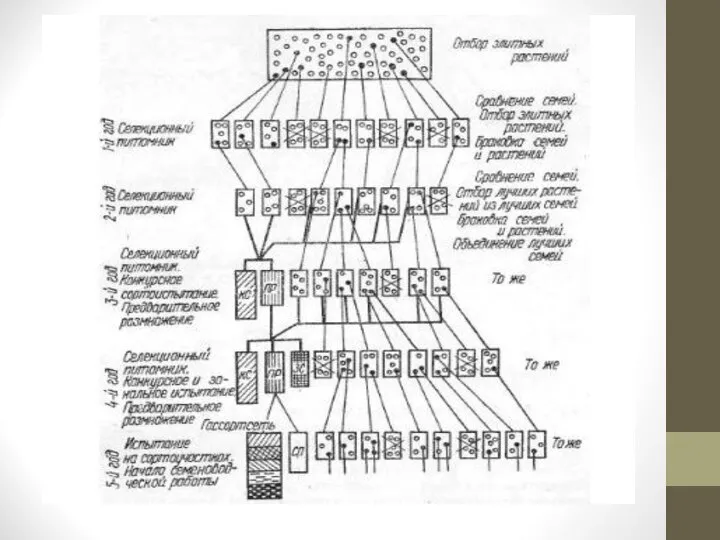
Слайд 20При семейственном отборе у перекрестноопыляющихся растений из исходной популяции отбирают элитные растения

с нужными хозяйственно полезными признаками и свойствами.
Урожай их семян на следующий год высевают раздельно по семьям в селекционном питомнике.
Семьи сравнивают между собой, худшие бракуют, из лучших снова отбирают элитные растения для посевов в селекционном питомнике следующего года.
Здесь также отбирают элитные растения, бракуют худшие растения и семьи.
Лучшие семьи объединяют и используют для конкурсного сортоиспытания (КС) и предварительного размножения (ПР).
При получении хороших результатов в конкурсном сортоиспытании новый сорт передают для посева на государственные сортоиспытательные участки, одновременно организуя его семеноводство (рис. 2).
Слайд 22У перекрестноопыляющихся культур каждое новое потомство получается в результате переопыления, т. е.

оно формируется на основе материнской и отцовской наследственности.
Часто при таком переопылении получается плохое потомство того или иного элитного растения не потому, что оно было плохим (его неправильно отобрали), а потому, что оно переопылилось с плохим растением.
Следовательно, в потомстве появились отрицательные отцовские признаки. Поэтому для перекрестноопыляющихся культур важно не только отбирать хорошие элитные растения, но и в пересевах удалять плохие, чтобы исключить их участие в опылении.
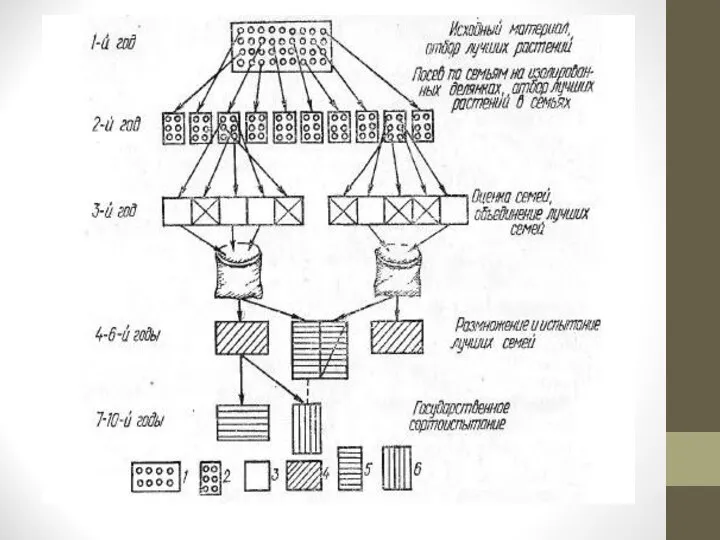
Исходя из этого, в селекционной работе используют два основных варианта многократного индивидуального отбора: индивидуально-семейственный и семейственно-групповой.
Индивидуально-семейственный отбор проводится по следующей схеме (рис. 3).
Слайд 24Семена каждого элитного растения высевают семьями изолированно одна от другой на отдельных

площадях.
Расстояние между семьями и другими посевами одной и той же культуры должно исключать возможность переопыления между ними.
В таких условиях переопыление происходит только в пределах семьи.
Чтобы избежать ухудшения потомства от переопыления с плохими растениями, их удаляют из каждой семьи до цветения.
В каждой семье проводится повторный отбор элитных растений, за исключением семей, выбракованных из-за болезни плохого общего развития и т. д.
Семена отобранных растений снова высевают семьями, также изолированно одна от другой, и снова в пределах каждой семьи проводят отбор и высев по семьям на изолированных участках.
Слайд 25Каждая семья состоит из довольно большого числа растений, выращиваемых в иных условиях,

чем материнское растение.
Если оно оказалось среди избранных вследствие модификационной изменчивости в результате выращивания в несколько лучших условиях, то его потомство не покажет преимущества перед другими и будет выбраковано.
Для селекции представляют интерес семьи с высокими показателями и однородностью по ценным признакам.
Большое разнообразие растений в семье указывает на гетерозиготность родоначальника семьи и преимущественный контроль анализируемого признака полигенами со сверхдоминантными эффектами.
Сохранить такой генетический контроль в следующих поколениях будет трудно вследствие смены аллельного состава гетерозигот при перекрестном опылении.
Выравненность семей напротив указывает на то, что анализируемый признак у родоначальника данной семьи контролировался преимущественно благоприятным сочетанием доминантных аллелей разных полигенов и гетерозиготностью по близким по активности аллелям.
Смена гетерозиготности в следующих поколениях в этом случае не приведет к сильному разнообразию растений и позволит в дальнейшем создать достаточно однородную сортовую популяцию.
Слайд 26В селекции перекрестно опыляемых растений применяют несколько вариантов семейственного отбора.
Их выбор

определяется, как желанием селекционера создать новую популяцию, в той или иной мере отличную от исходного материала, так и биологическими особенностями растений.
Их реакцией на инбридинг, возможностью оценки ценных признаков до или после цветения и другими причинами.
В работе с перекрестно опыляемыми растениями применяют: семейственный отбор без изоляции, семейственный отбор с изоляцией, метод парных скрещиваний и метод половинок.
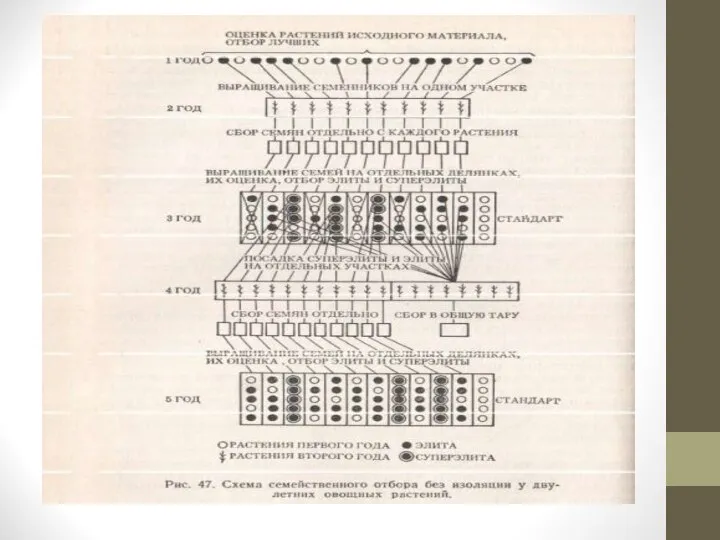
Семейственный отбор без изоляции.
При работе с растениями оценка которых и пересадка возможна до цветения, выделенные из исходного материала, лучшие по хозяйственным признакам экземпляры пересаживают на изолированный участок, где они взаимно переопыляются.
При работе с однолетними растениями, пересадка которых затруднена или невозможна, ценные растения оставляют на месте их выращивания, а остальной исходный материал бракуют.
Слайд 28Семена с каждого растения собирают отдельно.
В следующем поколении каждую семью выращивают

на отдельной делянке. Из лучших семей выделяют лучшие растения и используют их в качестве родоначальников будущих семей для следующего этапа семейственного отбора.
Так повторяют несколько раз до получения высококачественных и достаточно выровненных семей.
Каждая семья представляет собой маленькую популяцию, особи которой имеют в каждом локусе полигенного комплекса один аллель, полученный от ценного метеринского организма, и другой аллель от других, также ценных растений, участвующих в его опылении.
Различия генотипов особей в пределах семьи определяются как разнообразием по составу аллелей мужских, так и женских гамет, сформировавшихся в половых органах гетерозиготного растения. В целом выбор в лучшей семье растений - родоначальников семей следующего цикла семейственного отбора позволит выбрать наиболее удачные сочетания ценных аллелей.
Взаимным перекрестным опылением лучших растений лучшей семьи или нескольких близких семей получают семена оригинального селекционного материала, который в дальнейшем используется для продолжения работы по улучшению сорта и первичного семеноводства.
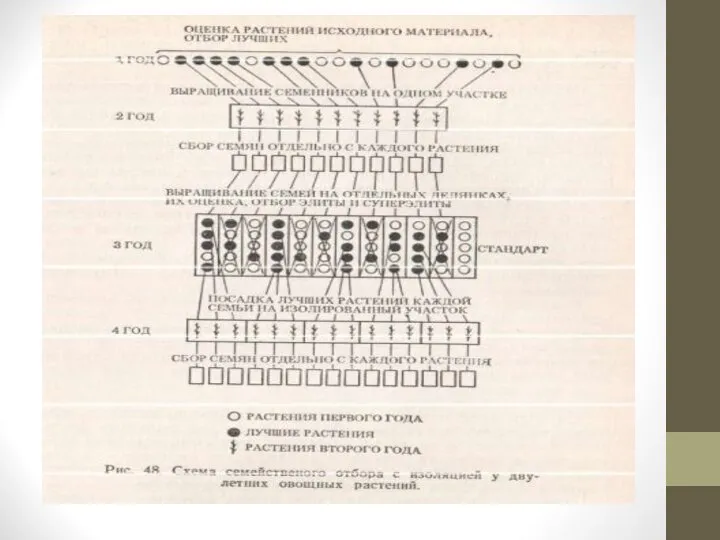
Слайд 30Семейственный отбор с изоляцией.
Выделенные на основании оценки исходного материала лучшие растения

высаживают вместе на изолированном участке для получения семян в результате взаимного переопыления.
Семена с каждого растения собирают раздельно, на следующий год семьи выращивают и оценивают на отдельных делянках.
Лучшие растения из лучших семей высаживают на отдельных изолированных участках, для их взаимного переопыления в пределах каждой семьи.
Семена собирают с лучших растений раздельно, учитывая их принадлежность к определенной семье. Полученные семьи выращивают на отдельных делянках; цикл отбора повторяется.
Таким образом, при семейственном отборе без изоляции на всех этапах отбора в создании генотипов растений следующих поколений участвуют растения опылители разных семей, а при семейственном отборе с изоляцией, начиная со второго этапа, в опылении участвуют только растения данной семьи.
Слайд 31Применение семейственного отбора с изоляцией наиболее оправдано когда в исходном материале присутствуют

сильно различающиеся по хозяйственным признакам ценные растения, когда появляется возможность вести селекцию в нескольких направлениях.
Крайней формой семейственного отбора с изоляцией является получение семян от выделенных в исходном материале растений с помощью инбридинга.
В этом случае разнообразие полученных семей будет определено только различиями в составе аллелей генотипов маточных растений.
В результате генетическая ценность той или иной семьи будет выявлена наиболее полно и точно.
При повторном применении инбридинга в полученных потомствах примерно, в полтора раза возрастет доля растений, гомозиготных по каждому локусу.
При работе с перекрестно опыляемыми растениями это не всегда желательно, так как гетерозиготность по большинству локусов полигенов – основное условие жизненности сорта перекрестно опыляемых растений.
Слайд 32Свободное взаимное опыление лучших растений в пределах инбредной семьи или объединение с

другой близкой семьей – наиболее оптимальный выход из положения.
Близкородственное скрещивание растений в пределах семьи, а тем более инбридинг у некоторых овощных растений быстро приводит к появлению инбредной депрессии – снижению жизненности растений вследствие перехода в гомозиготное состояние неблагоприятных рецессивных аллелей полигенов, что затрудняет оценку семей.
Чтобы устранить отрицательное действие близкородственного скрещивания, был предложен метод парных скрещиваний.
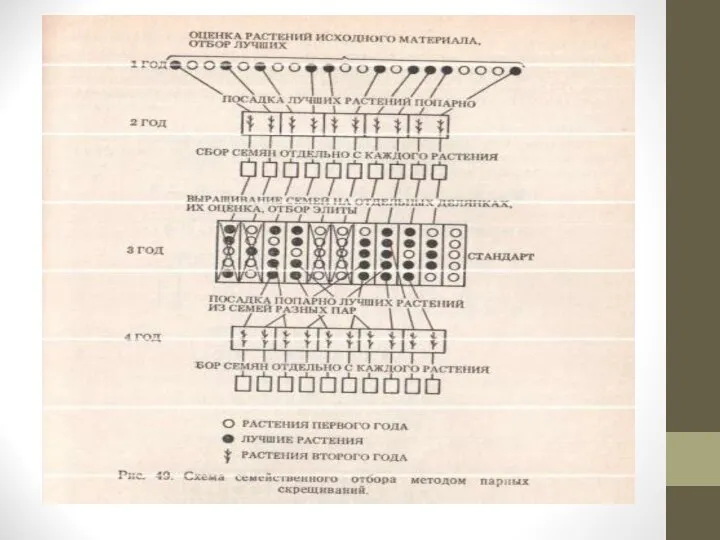
Метод парных скрещиваний. При применении метода парных скрещиваний выделенные из исходного материала ценные и близкие по фенотипу растения распределяют по парам.
Каждую пару высаживают под отдельный изолятор и обеспечивают взаимное опыление растений с помощью насекомых или вручную.
Слайд 34При работе с растениями, обладающими высокой семенной продуктивностью, достаточно поместить в один

изолятор побеги двух растущих рядом особей.
При применении этого метода в селекции однолетних, не переносящих пересадку растений, особи одной пары могут находиться в удалении друг от друга. В этом случае их скрещивание приходится осуществлять вручную, перенося пыльцу с одного растения на другое и обратно.
Семена с каждого растения или находящегося под изолятором побега собирают раздельно. Семьи выращивают и оценивают на отдельных делянках.
В следующий цикл парных скрещиваний подбирают растения из разных, но близких по хозяйственным признакам пар.
Из двух пар первого цикла составляют две пары второго цикла.
Парные скрещивания повторяют несколько поколений до достижения выравненности растений семей по ценным хозяйственным признакам. Их выравненность обеспечивается тем, что в создании генотипов особей участвует генофонд только четырех близких по фенотипу растений. Затем близкие семьи объединяют в сортовую популяцию.
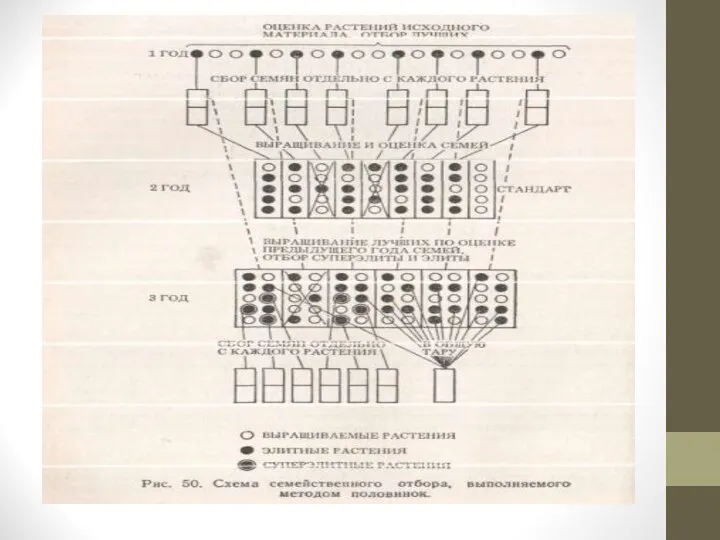
Слайд 36Метод половинок применяют при проведении семейственного отбора у перекрестноопыляемых растений, у которых

оценку товарных органов проводят после или во время цветения, когда взаимное опыление хороших и плохих особей уже произошло.
Например, как это наблюдается у растений семейства тыквенные, у сахарной кукурузы, сладкого перца и других культур при выделении лучших растений из исходного материала без применения инбридинга.
Собранные с них семена образуются в результате опыления не только пыльцой лучших, но и менее ценных растений, которые, как правило, имеют большее количество мужских цветков. В этих условиях в семьях, полученных от лучших растений, могут преобладать мало ценные особи.
При применении метода половинок от лучших растений исходного материала получают семена обычным способом, иногда с помощью инбридинга или, опыляя ценное растение пыльцой других ценных растений. Для селекционного испытания семей используют небольшую часть полученных семян.
На основании полученных оценок на следующий год высевают оставшиеся семена только лучших семей и среди них выделяют родоначальников семей следующего этапа испытания и отбора.
Слайд 38Собранные с них семена сформировались в результате оплодотворения яйцеклеток ценных растений мужскими

гаметами также ценных растений.
Так поступают в течение нескольких генераций, пока не будут получены достаточно выровненные семьи. Близкие по хозяйственным признакам семьи объединяют в сортовую популяцию.
Преимущества метода половинок заключаются в его специфичной применимости к определенным перекрестно опыляемым растениям, у которых товарными органами являются плоды или семена.
Его недостаток – вдвое большая длительность выполнения, так как оценку семей и выделение родоначальников семей следующего поколения производят в разные годы.
По способам получения семян от родоначальников семей он аналогичен другим вариантам семейственного отбора.
Так, при получении семян от свободного опыления он близок к семейственному отбору без изоляции, при применении инбридинга – к семейственному отбору с изоляцией, а при опылении – пыльцой лучших растений – к парному методу.
Слайд 39Следует учитывать, что при применении жестких методов семейственного отбора: семейственного отбора с

изоляцией, парного метода и особенно инбридинга в сортовую популяцию желательно включать не менее 3-4 семей.
Из перечисленных выше методов в селекции перекрестноопыляемых растений наиболее часто применялись массовые отборы, как наиболее легко выполнимые, позволяющие получать достаточно большие количества семян и показывающие достаточно точно достоинства и недостатки создаваемых селекционных популяций.
Семейственные отборы позволяют более радикально изменять состав селекционного материала, но они более трудоемки, их обычно применяют, когда массовый отбор не дает нужного эффекта.
Обычно, в разных поколениях второго этапа селекционного процесса применяют как массовый, так и тот или иной вариант семейственного отбора.

 Постэмбриональное развитие организмов. Прямое и непрямое развитие
Постэмбриональное развитие организмов. Прямое и непрямое развитие Наследование групп крови
Наследование групп крови Презентация на тему Класс Хрящевые рыбы
Презентация на тему Класс Хрящевые рыбы  Решение задач с биологическим содержанием. История создания клеточной теории
Решение задач с биологическим содержанием. История создания клеточной теории Влияние физических упражнений на кровеносную, дыхательную и нервную системы
Влияние физических упражнений на кровеносную, дыхательную и нервную системы Прощание с ботаникой
Прощание с ботаникой Как заглянуть в митохондрию с помощью наночастиц
Как заглянуть в митохондрию с помощью наночастиц Классические методы селекции
Классические методы селекции Организм многоклеточного животного. Тип Кишечнополостные. 7 класс
Организм многоклеточного животного. Тип Кишечнополостные. 7 класс Презентация на тему Грибы - паразиты 6 класс
Презентация на тему Грибы - паразиты 6 класс  Мини-ландшафт в бутылке или флорариум
Мини-ландшафт в бутылке или флорариум Растения пустыни
Растения пустыни HydraMax. Оптимальная Гидратация
HydraMax. Оптимальная Гидратация Презентация на тему ПРАКТИЧЕСКОЕ ПРИМЕНЕНИЕ ГЕННОЙ ИНЖЕНЕРИИ
Презентация на тему ПРАКТИЧЕСКОЕ ПРИМЕНЕНИЕ ГЕННОЙ ИНЖЕНЕРИИ  Презентация на тему ОРГАНЫ ДЫХАНИЯ
Презентация на тему ОРГАНЫ ДЫХАНИЯ  Агатис южный
Агатис южный Ферменты
Ферменты Ткани
Ткани _Как работают в лаборатории
_Как работают в лаборатории Ядовитые растения
Ядовитые растения Глаз - оптическая система!
Глаз - оптическая система! Изменения строения и жизнедеятельности организмов. Основные направления эволюции
Изменения строения и жизнедеятельности организмов. Основные направления эволюции Обобщающий урок по теме: Жизнь растений
Обобщающий урок по теме: Жизнь растений Методы внутривидового типирования микроорганизмов
Методы внутривидового типирования микроорганизмов Биохимия мышц
Биохимия мышц Презентация на тему Вид. Критерии вида
Презентация на тему Вид. Критерии вида  Нарушение обмена витаминов
Нарушение обмена витаминов Современный этап эволюции человека
Современный этап эволюции человека