- Мозг и память

Содержание
- 2. Две предыдущие лекции были посвящены в основном таким компонентам поведения и психической деятельности, как запускаемые извне
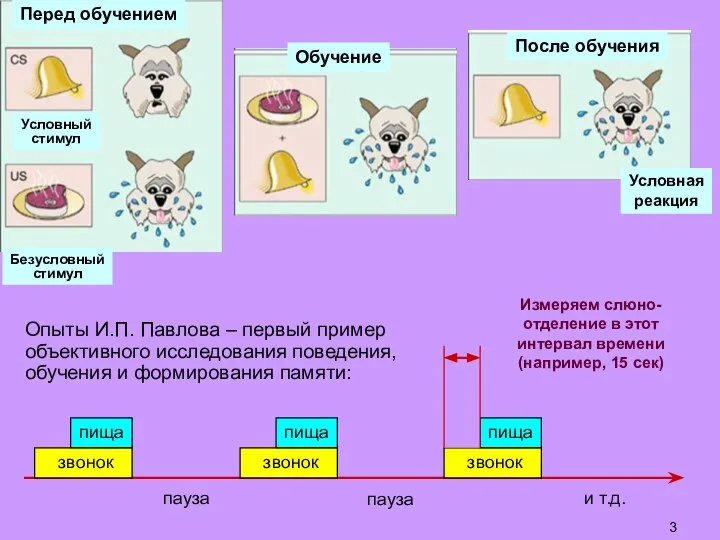
- 3. Опыты И.П. Павлова – первый пример объективного исследования поведения, обучения и формирования памяти: Перед обучением Обучение
- 4. звонок пища пауза пауза и т.д. звонок пища звонок пища Измеряем слюно- отделение в этот интервал
- 5. S-образная кривая обучения: свидетельствует о том, что исходно незначимый стимул стал значимым и теперь запускает реакцию
- 6. ПИЩА СЛЮНО- ОТДЕЛЕНИЕ ЗВОНОК безусловный рефлекс условный рефлекс: в коре больших полушарий образуется новый канал для
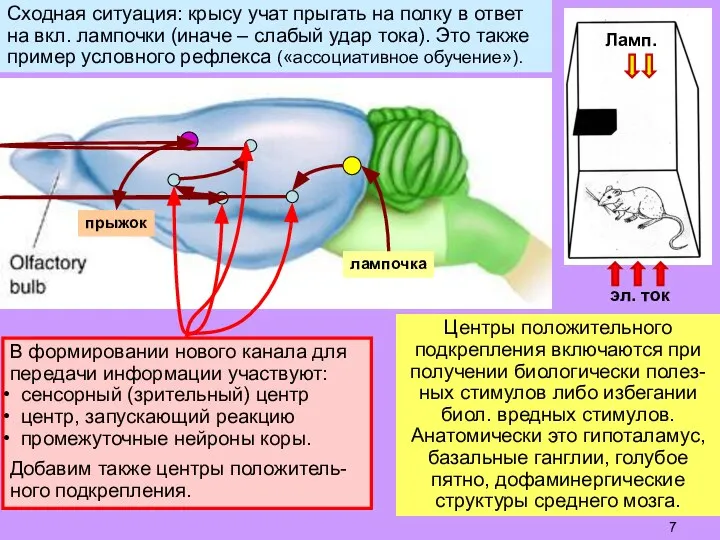
- 7. Сходная ситуация: крысу учат прыгать на полку в ответ на вкл. лампочки (иначе – слабый удар
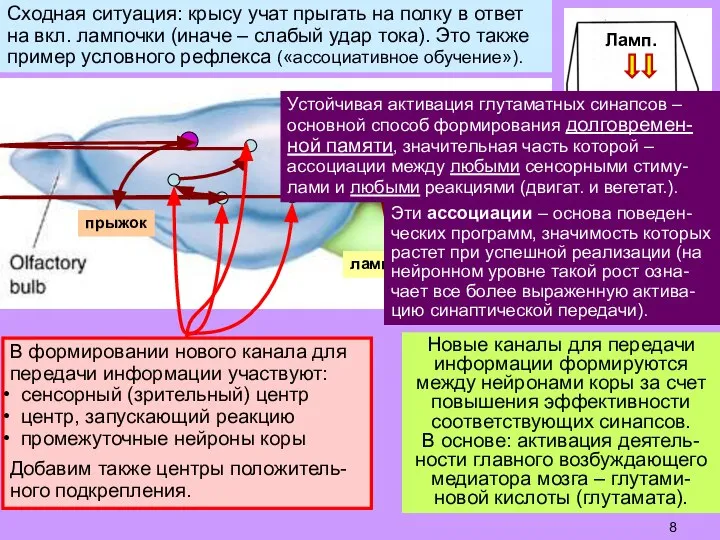
- 8. Новые каналы для передачи информации формируются между нейронами коры за счет повышения эффективности соответствующих синапсов. В
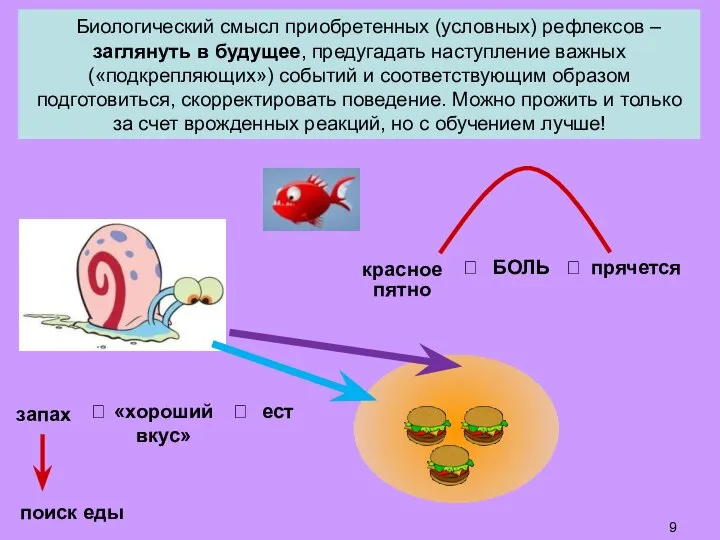
- 9. Биологический смысл приобретенных (условных) рефлексов – заглянуть в будущее, предугадать наступление важных («подкрепляющих») событий и соответствующим
- 10. В ходе эволюции организмы быстро умнеют – то есть все успешнее формируют приобретенные программы. Дополнительно подталкивает
- 11. Разнообразие типов памяти: кратковременная и долговременная; ассоциативная и неассоциативная (без положительного подкрепления). В любом случае –
- 12. Синаптичес- кая активность: межклеточная передача сигнала в контактах-синапсах с помощью особых веществ (медиаторов), вызыва-ющих реакцию следующей
- 14. Основные части синапса: * окончание отростка нейрона * синаптические пузырьки (везикулы) с медиатором * пресинаптическая мембрана
- 15. Выброс (экзоцитоз) медиатора в синаптическую щель происходит после появления ПД, который вызывает открывание электрочувствительных Са2+-каналов (примерно
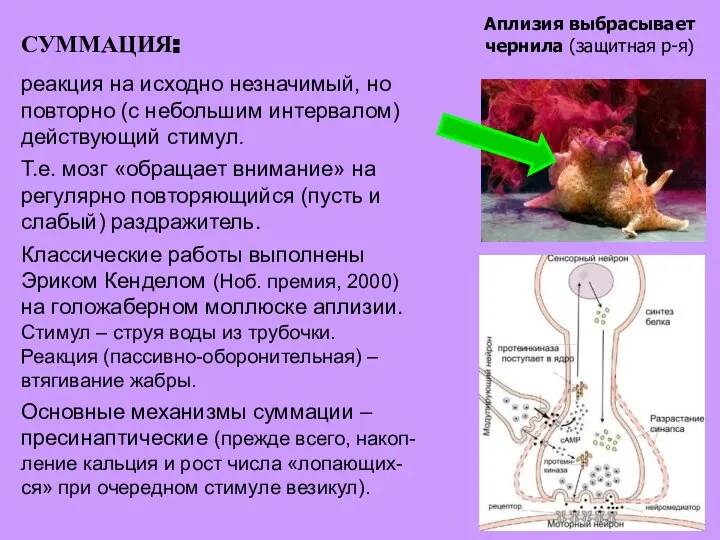
- 16. СУММАЦИЯ: реакция на исходно незначимый, но повторно (с небольшим интервалом) действующий стимул. Т.е. мозг «обращает внимание»
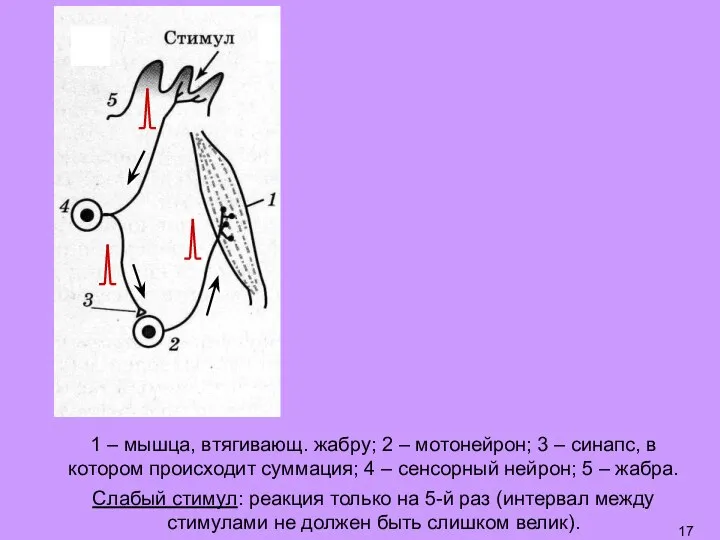
- 17. 1 – мышца, втягивающ. жабру; 2 – мотонейрон; 3 – синапс, в котором происходит суммация; 4
- 18. 1 – мышца, втягивающ. жабру; 2 – мотонейрон; 3 – синапс, в котором происходит суммация; 4
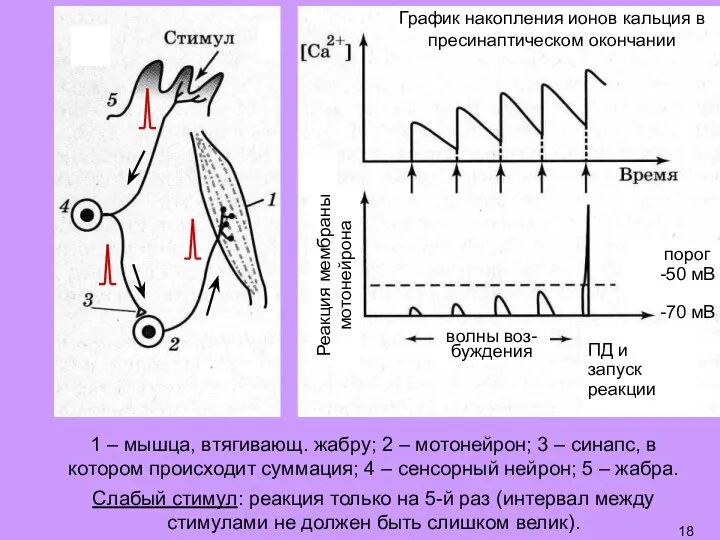
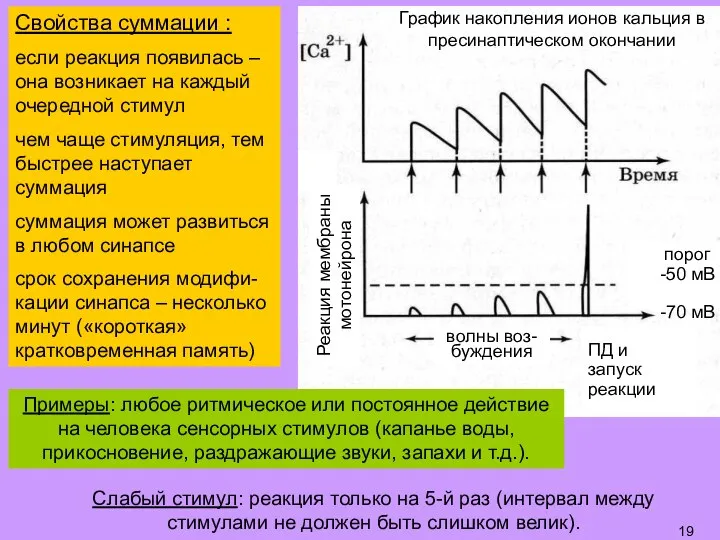
- 19. Слабый стимул: реакция только на 5-й раз (интервал между стимулами не должен быть слишком велик). График
- 20. Долговременная потенциация (ДолгП): пусть реакция на слабый стимул отсутствует далее – подаем сильный стимул, запускающий реакцию
- 21. Уникальная особенность NMDA-рецепторов (в отличие от других рецепторов к Glu) состоит в том, что их канал
- 22. Наиболее очевидный способ удаления Mg2+: ПД, запущенный с помощью не-NMDA-рецепторов к Glu, находящихся на той же
- 23. 1 – не-NMDA-рецептор; 2 – NMDA-рецептор и ион Mg2+ 3 – везикула с Glu 4 –
- 24. Подобного рода синапсы, способные практически мгновенно увеличить эффективность работы, характерны для коры больш. полушарий и, осо-бенно,
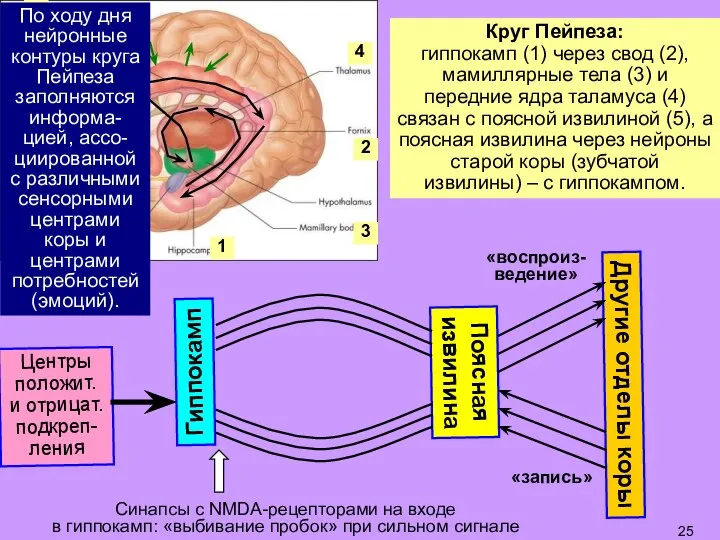
- 25. По ходу дня нейронные контуры круга Пейпеза заполняются информа-цией, ассо-циированной с различными сенсорными центрами коры и
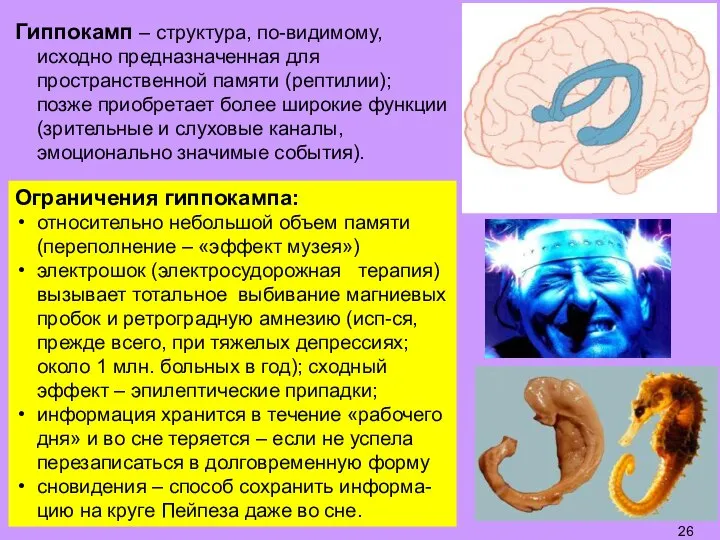
- 26. Гиппокамп – структура, по-видимому, исходно предназначенная для пространственной памяти (рептилии); позже приобретает более широкие функции (зрительные
- 27. Импринтинг = запечатление. Конрад Лоренц, серые гуси. «Впечатывание в мозг» определен-ной ключевой информации в строго определенный
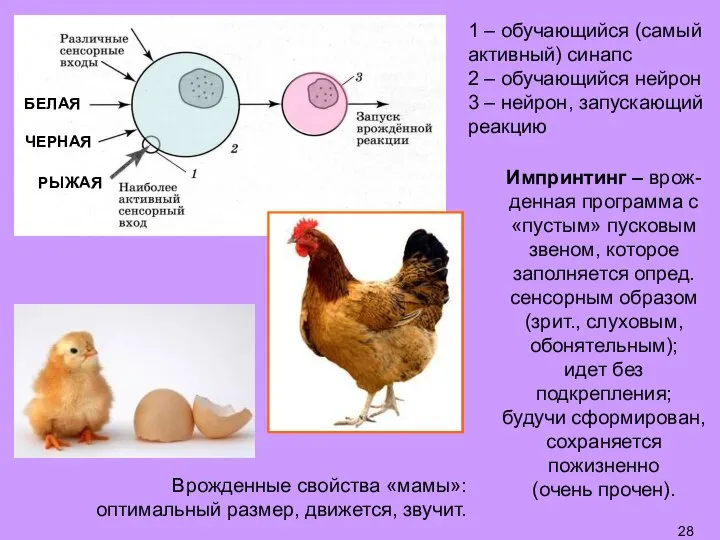
- 28. 1 – обучающийся (самый активный) синапс 2 – обучающийся нейрон 3 – нейрон, запускающий реакцию БЕЛАЯ
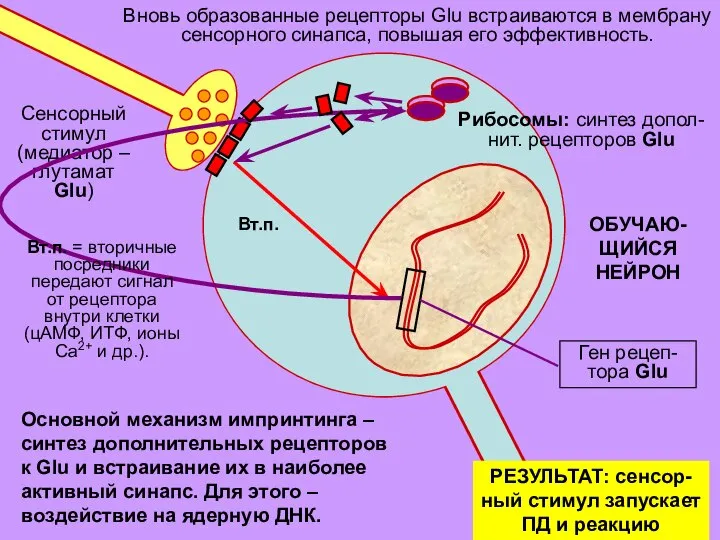
- 29. ОБУЧАЮ- ЩИЙСЯ НЕЙРОН Вновь образованные рецепторы Glu встраиваются в мембрану сенсорного синапса, повышая его эффективность. РЕЗУЛЬТАТ:
- 30. Импринтинг – модель формиро-вания долговременной памяти. Лосята – импринтинг образа матери; лосихи – импринтинг запаха детеныша.
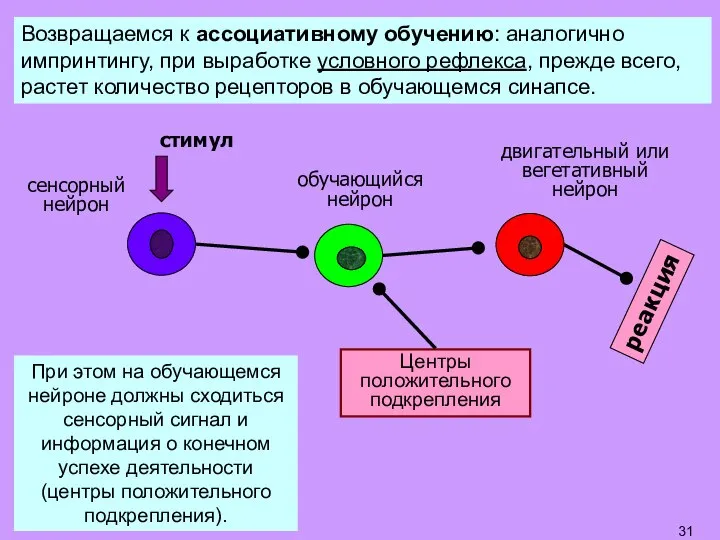
- 31. Возвращаемся к ассоциативному обучению: аналогично импринтингу, при выработке условного рефлекса, прежде всего, растет количество рецепторов в
- 32. ОБУЧАЮЩИЙСЯ НЕЙРОН Вновь образованные рецепторы Glu встраиваются в мембрану сенсорного синапса. Вт.п. – вторичные посредники (цАМФ,
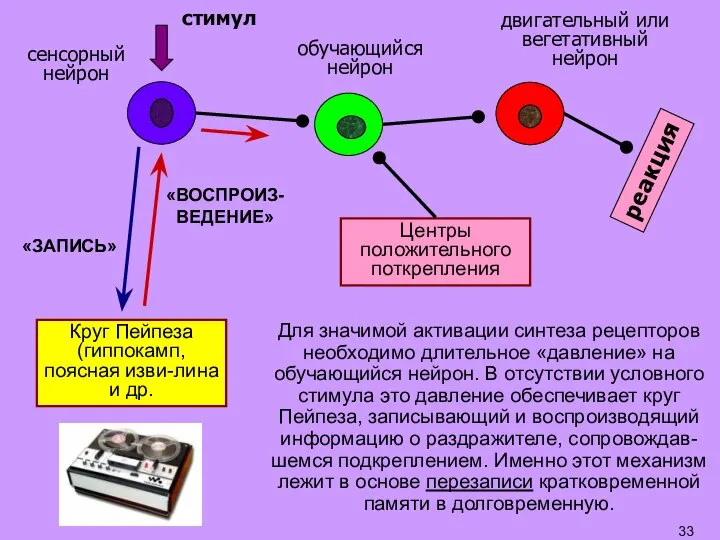
- 33. Для значимой активации синтеза рецепторов необходимо длительное «давление» на обучающийся нейрон. В отсутствии условного стимула это
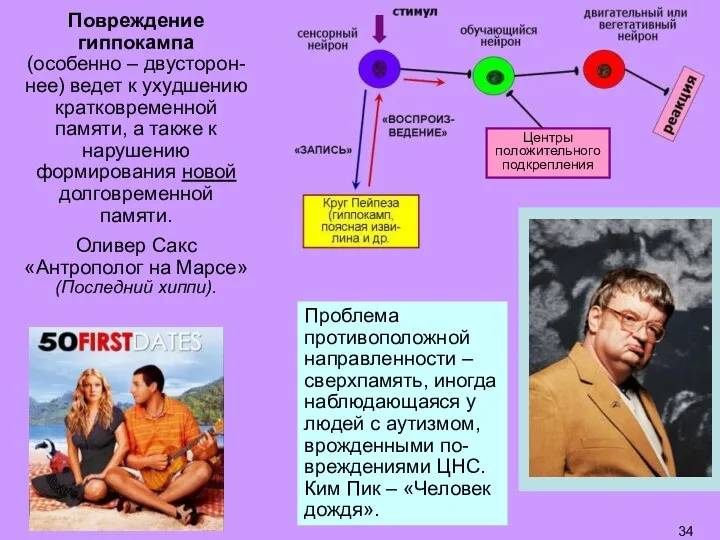
- 34. Повреждение гиппокампа (особенно – двусторон-нее) ведет к ухудшению кратковременной памяти, а также к нарушению формирования новой
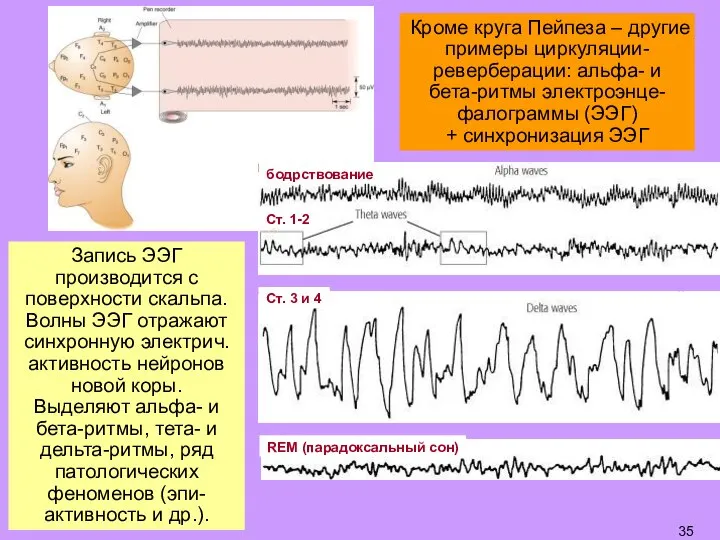
- 35. Запись ЭЭГ производится с поверхности скальпа. Волны ЭЭГ отражают синхронную электрич. активность нейронов новой коры. Выделяют
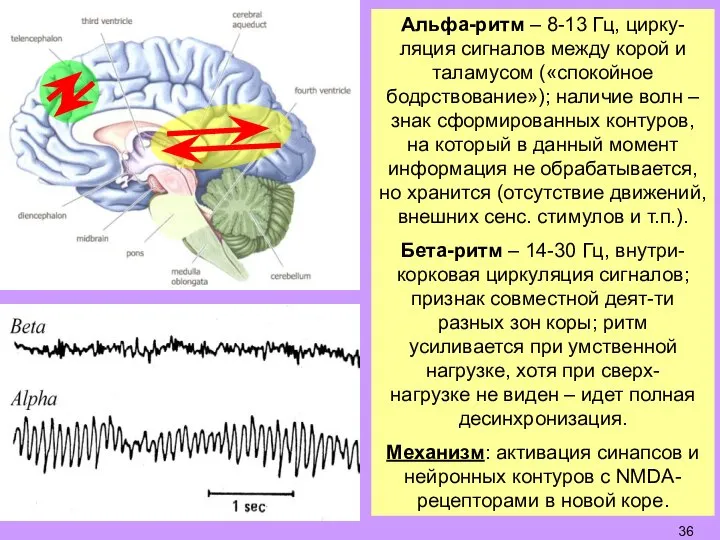
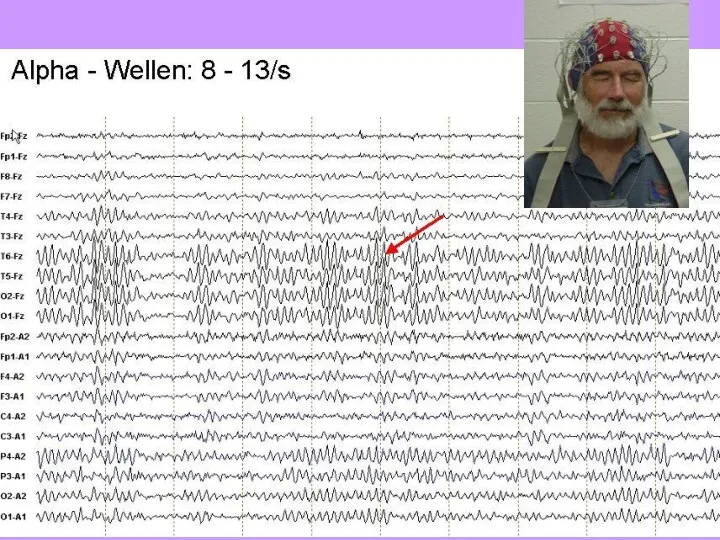
- 36. Альфа-ритм – 8-13 Гц, цирку-ляция сигналов между корой и таламусом («спокойное бодрствование»); наличие волн – знак
- 38. Ритмы ЭЭГ четко отражают уровень бодрствования: чем он ниже, тем меньше частота волн и выше амплитуда.
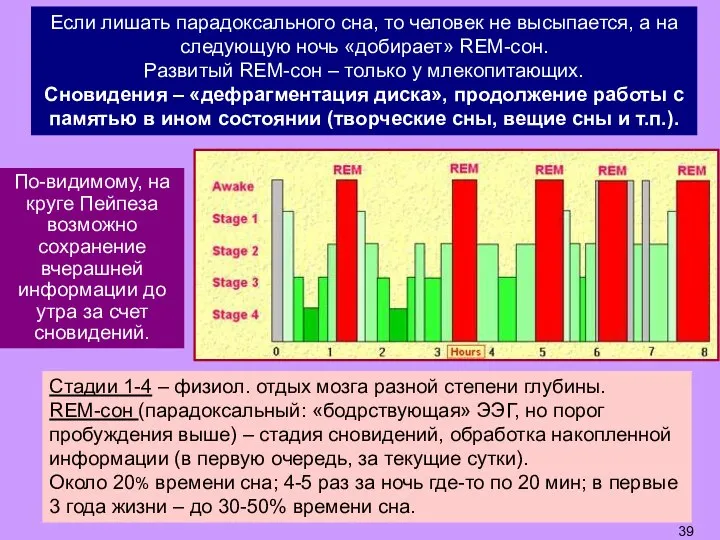
- 39. Стадии 1-4 – физиол. отдых мозга разной степени глубины. REM-сон (парадоксальный: «бодрствующая» ЭЭГ, но порог пробуждения
- 40. Если лишать парадоксального сна, то человек не высыпается, а на следующую ночь «добирает» REM-сон. Развитый REM-сон
- 41. Итак, память – это сетевое свойство нейронов коры больших полушарий. При этом мы пока говорили о
- 42. Итак, память – это сетевое свойство нейронов коры больших полушарий При этом мы пока говорили о
- 43. НООТРОПЫ. Выделяются в особую группу лекарственных препаратов. Их объединяет способность стимулировать высшие психические функции (память, мышление),
- 44. Не путать ноотропы с веществами, влияющими на работу синапсов – особенно, если имеется позитивный эмоцио-нальный компонент.
- 45. Не путать ноотропы с веществами, влияющими на работу синапсов – особенно, если имеется позитивный эмоцио-нальный компонент.
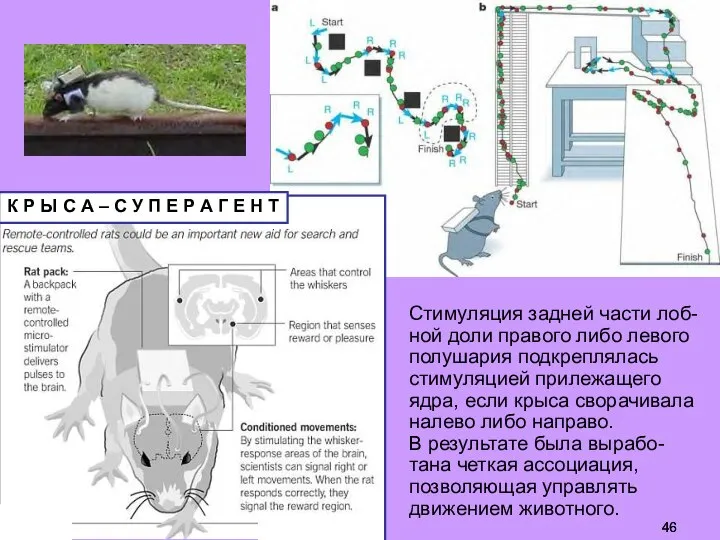
- 46. Стимуляция задней части лоб-ной доли правого либо левого полушария подкреплялась стимуляцией прилежащего ядра, если крыса сворачивала
- 47. Все это – «положительное обучение», приводящее к формир-ю новых каналов для передачи информации. Мозг умеет также
- 48. Спасибо за внимание! В следующий раз – зачет. От Александровой Ирины до Нестеровой Галины – к
- 50. Скачать презентацию

Слайд 3Опыты И.П. Павлова – первый пример объективного исследования поведения, обучения и формирования
Опыты И.П. Павлова – первый пример объективного исследования поведения, обучения и формирования
Слайд 4 звонок
пища
пауза
пауза
и т.д.
звонок
пища
звонок
пища
Измеряем слюно- отделение в этот
звонок
пища
пауза
пауза
и т.д.
звонок
пища
звонок
пища
Измеряем слюно- отделение в этот
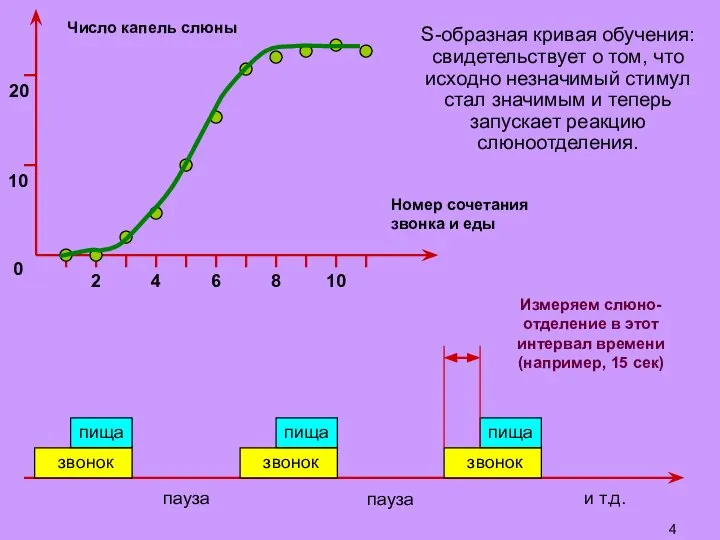
Слайд 5S-образная кривая обучения:
свидетельствует о том, что исходно незначимый стимул стал значимым и
S-образная кривая обучения:
свидетельствует о том, что исходно незначимый стимул стал значимым и
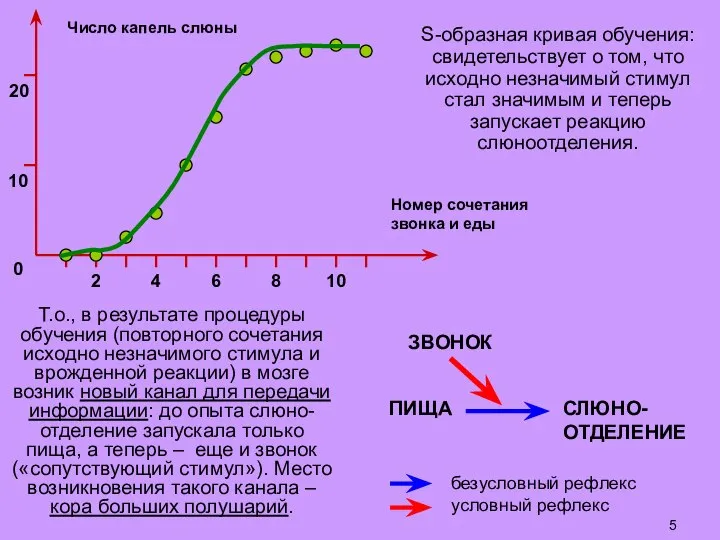
Слайд 6ПИЩА
СЛЮНО-
ОТДЕЛЕНИЕ
ЗВОНОК
безусловный рефлекс
условный рефлекс: в коре
больших полушарий
образуется новый канал
для передачи информации
между слуховой
и вкусовой
(островковой)
корой;
ПИЩА
СЛЮНО-
ОТДЕЛЕНИЕ
ЗВОНОК
безусловный рефлекс
условный рефлекс: в коре
больших полушарий
образуется новый канал
для передачи информации
между слуховой
и вкусовой
(островковой)
корой;
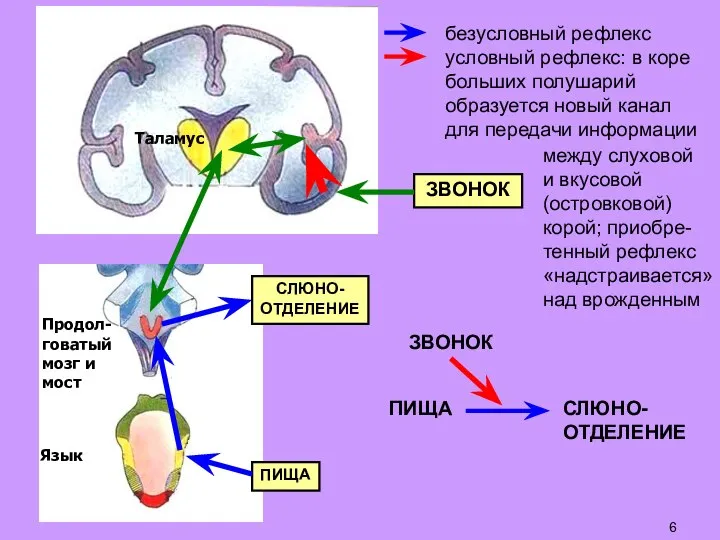
Слайд 7Сходная ситуация: крысу учат прыгать на полку в ответ
на вкл. лампочки (иначе
Сходная ситуация: крысу учат прыгать на полку в ответ
на вкл. лампочки (иначе
Слайд 8Новые каналы для передачи информации формируются между нейронами коры за счет повышения
Новые каналы для передачи информации формируются между нейронами коры за счет повышения
Слайд 9 Биологический смысл приобретенных (условных) рефлексов – заглянуть в будущее, предугадать наступление
Биологический смысл приобретенных (условных) рефлексов – заглянуть в будущее, предугадать наступление
Слайд 10 В ходе эволюции организмы быстро умнеют – то есть все успешнее
В ходе эволюции организмы быстро умнеют – то есть все успешнее

Слайд 11 Разнообразие типов памяти:
кратковременная и долговременная; ассоциативная и неассоциативная
(без положительного подкрепления).
В
Разнообразие типов памяти:
кратковременная и долговременная; ассоциативная и неассоциативная
(без положительного подкрепления).
В
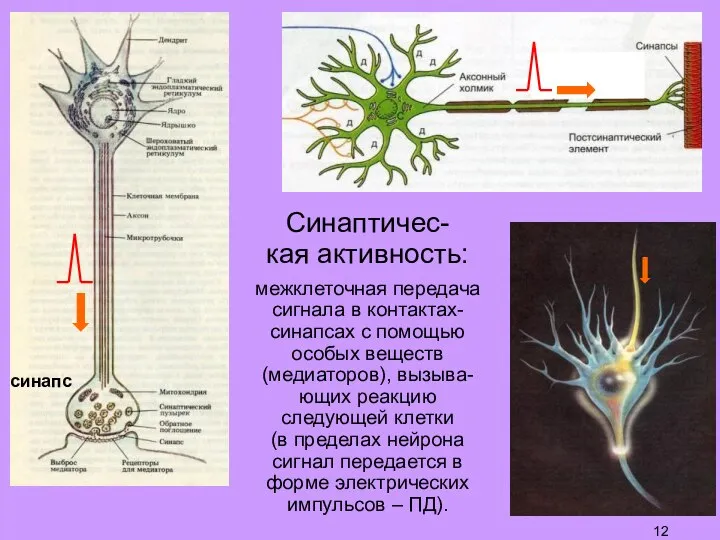
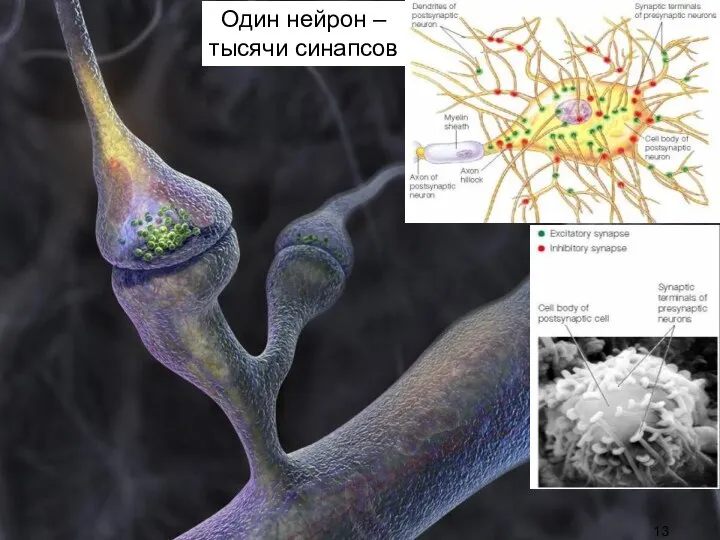
Слайд 12Синаптичес-
кая активность:
межклеточная передача
сигнала в контактах-синапсах с помощью особых веществ (медиаторов), вызыва-ющих
Синаптичес-
кая активность:
межклеточная передача
сигнала в контактах-синапсах с помощью особых веществ (медиаторов), вызыва-ющих
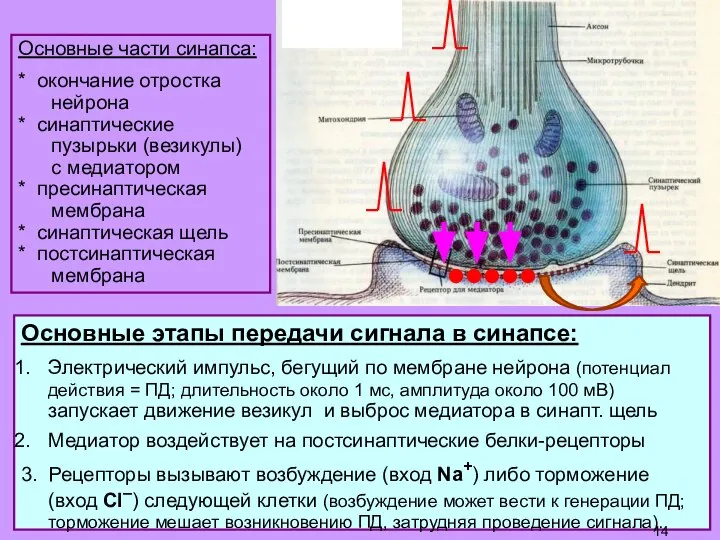
Слайд 14Основные части синапса:
* окончание отростка
нейрона
* синаптические
пузырьки (везикулы)
с медиатором
*
Основные части синапса:
* окончание отростка
нейрона
* синаптические
пузырьки (везикулы)
с медиатором
*
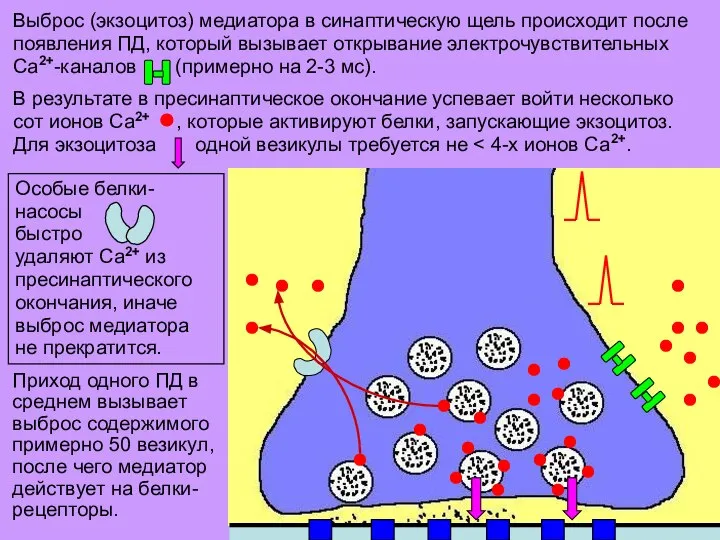
Слайд 15Выброс (экзоцитоз) медиатора в синаптическую щель происходит после появления ПД, который вызывает
Выброс (экзоцитоз) медиатора в синаптическую щель происходит после появления ПД, который вызывает
Слайд 16СУММАЦИЯ:
реакция на исходно незначимый, но повторно (с небольшим интервалом) действующий стимул.
Т.е. мозг
СУММАЦИЯ:
реакция на исходно незначимый, но повторно (с небольшим интервалом) действующий стимул.
Т.е. мозг
Слайд 171 – мышца, втягивающ. жабру; 2 – мотонейрон; 3 – синапс, в
1 – мышца, втягивающ. жабру; 2 – мотонейрон; 3 – синапс, в
Слайд 181 – мышца, втягивающ. жабру; 2 – мотонейрон; 3 – синапс, в
1 – мышца, втягивающ. жабру; 2 – мотонейрон; 3 – синапс, в
Слайд 19
Слабый стимул: реакция только на 5-й раз (интервал между стимулами не должен
Слабый стимул: реакция только на 5-й раз (интервал между стимулами не должен
Слайд 20Долговременная потенциация (ДолгП):
пусть реакция на слабый стимул отсутствует
далее – подаем сильный стимул,
Долговременная потенциация (ДолгП):
пусть реакция на слабый стимул отсутствует
далее – подаем сильный стимул,
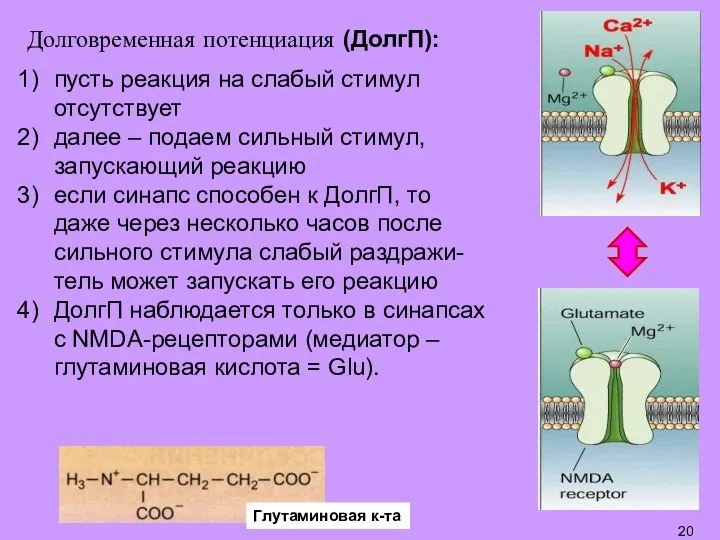
Слайд 21Уникальная особенность NMDA-рецепторов (в отличие от других рецепторов к Glu) состоит в
Уникальная особенность NMDA-рецепторов (в отличие от других рецепторов к Glu) состоит в

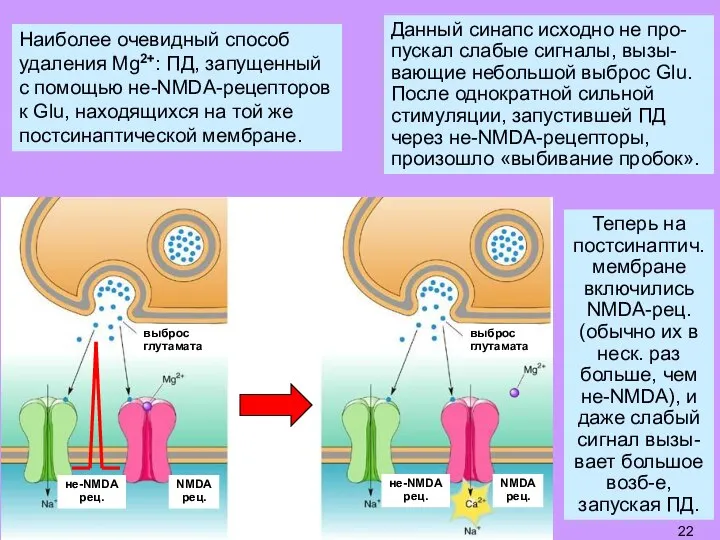
Слайд 22Наиболее очевидный способ
удаления Mg2+: ПД, запущенный с помощью не-NMDA-рецепторов к Glu, находящихся
Наиболее очевидный способ
удаления Mg2+: ПД, запущенный с помощью не-NMDA-рецепторов к Glu, находящихся
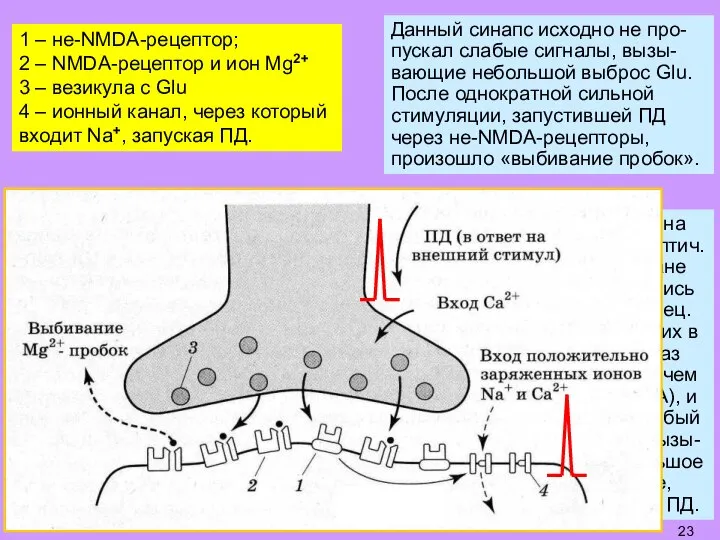
Слайд 231 – не-NMDA-рецептор;
2 – NMDA-рецептор и ион Mg2+
3 – везикула с
1 – не-NMDA-рецептор;
2 – NMDA-рецептор и ион Mg2+
3 – везикула с
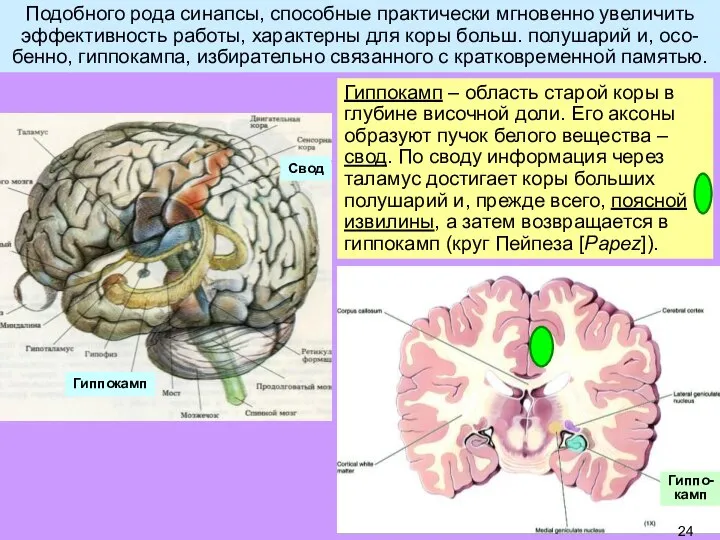
Слайд 24Подобного рода синапсы, способные практически мгновенно увеличить эффективность работы, характерны для коры
Подобного рода синапсы, способные практически мгновенно увеличить эффективность работы, характерны для коры
Слайд 25По ходу дня нейронные контуры круга Пейпеза заполняются информа-цией, ассо-циированной с различными
По ходу дня нейронные контуры круга Пейпеза заполняются информа-цией, ассо-циированной с различными
Слайд 26Гиппокамп – структура, по-видимому, исходно предназначенная для пространственной памяти (рептилии); позже приобретает
Гиппокамп – структура, по-видимому, исходно предназначенная для пространственной памяти (рептилии); позже приобретает
Слайд 27Импринтинг = запечатление.
Конрад Лоренц, серые гуси.
«Впечатывание в мозг» определен-ной ключевой информации в
Импринтинг = запечатление.
Конрад Лоренц, серые гуси.
«Впечатывание в мозг» определен-ной ключевой информации в

Слайд 281 – обучающийся (самый активный) синапс
2 – обучающийся нейрон
3 – нейрон, запускающий
1 – обучающийся (самый активный) синапс
2 – обучающийся нейрон
3 – нейрон, запускающий
Слайд 29ОБУЧАЮ-
ЩИЙСЯ
НЕЙРОН
Вновь образованные рецепторы Glu встраиваются в мембрану
сенсорного синапса, повышая его эффективность.
РЕЗУЛЬТАТ: сенсор-
ный
ОБУЧАЮ-
ЩИЙСЯ
НЕЙРОН
Вновь образованные рецепторы Glu встраиваются в мембрану
сенсорного синапса, повышая его эффективность.
РЕЗУЛЬТАТ: сенсор-
ный
Слайд 30Импринтинг – модель формиро-вания долговременной памяти.
Лосята – импринтинг образа матери; лосихи –
Импринтинг – модель формиро-вания долговременной памяти.
Лосята – импринтинг образа матери; лосихи –

Слайд 31Возвращаемся к ассоциативному обучению: аналогично импринтингу, при выработке условного рефлекса, прежде всего,
Возвращаемся к ассоциативному обучению: аналогично импринтингу, при выработке условного рефлекса, прежде всего,
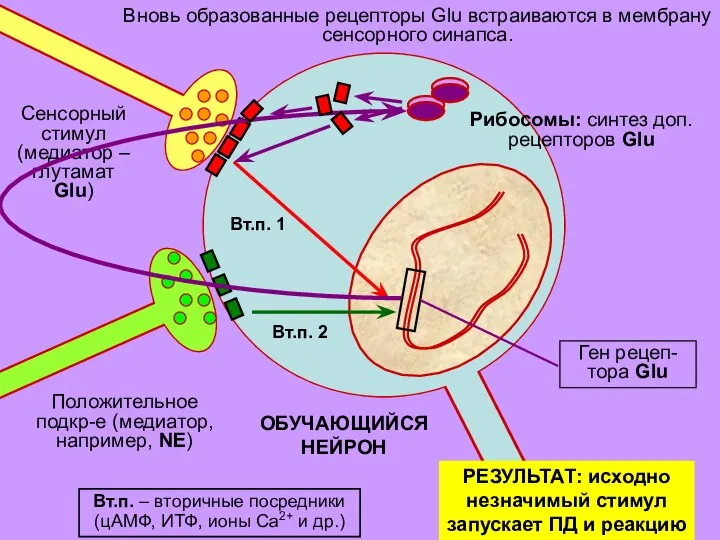
Слайд 32ОБУЧАЮЩИЙСЯ
НЕЙРОН
Вновь образованные рецепторы Glu встраиваются в мембрану
сенсорного синапса.
Вт.п. – вторичные посредники (цАМФ,
ОБУЧАЮЩИЙСЯ
НЕЙРОН
Вновь образованные рецепторы Glu встраиваются в мембрану
сенсорного синапса.
Вт.п. – вторичные посредники (цАМФ,
Слайд 33Для значимой активации синтеза рецепторов необходимо длительное «давление» на обучающийся нейрон. В
Для значимой активации синтеза рецепторов необходимо длительное «давление» на обучающийся нейрон. В
Слайд 34Повреждение гиппокампа
(особенно – двусторон-нее) ведет к ухудшению кратковременной памяти, а также
Повреждение гиппокампа
(особенно – двусторон-нее) ведет к ухудшению кратковременной памяти, а также
Слайд 35Запись ЭЭГ производится с поверхности скальпа. Волны ЭЭГ отражают синхронную электрич. активность
Запись ЭЭГ производится с поверхности скальпа. Волны ЭЭГ отражают синхронную электрич. активность
Слайд 36Альфа-ритм – 8-13 Гц, цирку-ляция сигналов между корой и таламусом («спокойное бодрствование»);
Альфа-ритм – 8-13 Гц, цирку-ляция сигналов между корой и таламусом («спокойное бодрствование»);
Слайд 38Ритмы ЭЭГ четко отражают уровень бодрствования: чем он ниже, тем меньше частота
Ритмы ЭЭГ четко отражают уровень бодрствования: чем он ниже, тем меньше частота
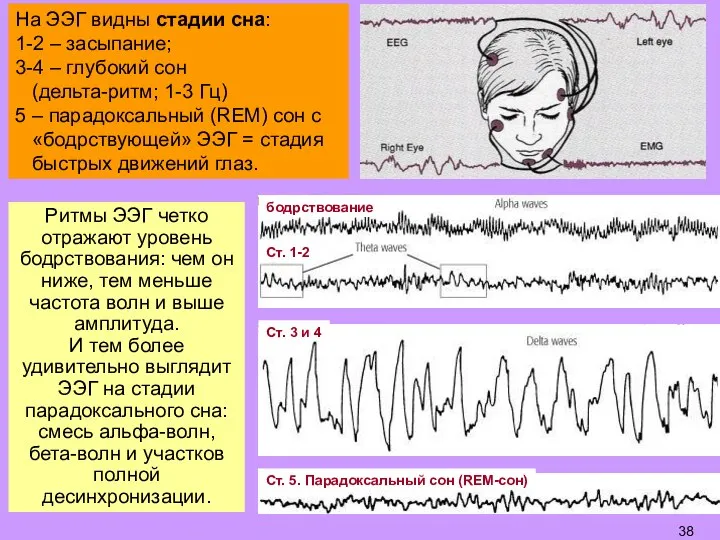
Слайд 39Стадии 1-4 – физиол. отдых мозга разной степени глубины.
REM-сон (парадоксальный: «бодрствующая»
Стадии 1-4 – физиол. отдых мозга разной степени глубины.
REM-сон (парадоксальный: «бодрствующая»
Слайд 40Если лишать парадоксального сна, то человек не высыпается, а на следующую ночь
Если лишать парадоксального сна, то человек не высыпается, а на следующую ночь

Слайд 41Итак, память – это сетевое свойство нейронов коры больших полушарий.
При этом мы
Итак, память – это сетевое свойство нейронов коры больших полушарий.
При этом мы

Слайд 42Итак, память – это сетевое свойство нейронов коры больших полушарий
При этом
Итак, память – это сетевое свойство нейронов коры больших полушарий
При этом
Слайд 43НООТРОПЫ.
Выделяются в особую группу лекарственных препаратов. Их объединяет способность стимулировать высшие психические
НООТРОПЫ.
Выделяются в особую группу лекарственных препаратов. Их объединяет способность стимулировать высшие психические
Слайд 44Не путать ноотропы с веществами, влияющими на работу синапсов – особенно, если
Не путать ноотропы с веществами, влияющими на работу синапсов – особенно, если
Слайд 45Не путать ноотропы с веществами, влияющими на работу синапсов – особенно, если
Не путать ноотропы с веществами, влияющими на работу синапсов – особенно, если

Слайд 46Стимуляция задней части лоб-ной доли правого либо левого полушария подкреплялась стимуляцией прилежащего
Стимуляция задней части лоб-ной доли правого либо левого полушария подкреплялась стимуляцией прилежащего
Слайд 47Все это – «положительное обучение», приводящее к
формир-ю новых каналов для передачи
Все это – «положительное обучение», приводящее к
формир-ю новых каналов для передачи

Слайд 48Спасибо за внимание!
В следующий раз – зачет.
От Александровой Ирины до Нестеровой
Спасибо за внимание!
В следующий раз – зачет.
От Александровой Ирины до Нестеровой

 День науки
День науки Haustiere in unserem Alltag
Haustiere in unserem Alltag Влияние внешних и внутренних факторов на транспирацию
Влияние внешних и внутренних факторов на транспирацию Своя игра. Генетика
Своя игра. Генетика Композиция из семян растений (технология)
Композиция из семян растений (технология) Многообразие Простейших. Одноклеточные. 1
Многообразие Простейших. Одноклеточные. 1 Пуансеттия - рождественская звезда
Пуансеттия - рождественская звезда Генетизация жизни
Генетизация жизни Царство растения. Основные отделы растений и их характеристики
Царство растения. Основные отделы растений и их характеристики Птицы Свердловской области
Птицы Свердловской области Жизнедеятельность организмов
Жизнедеятельность организмов Рост растений
Рост растений Желудочковая система головного мозга
Желудочковая система головного мозга Головной мозг
Головной мозг Р. Броун, его вклад в развитие физики
Р. Броун, его вклад в развитие физики Взаимоотношения организма и среды. Невидимые нити природы
Взаимоотношения организма и среды. Невидимые нити природы Разнообразие природы Ленинградской области
Разнообразие природы Ленинградской области Кожа
Кожа Skābes dabā
Skābes dabā Вредители хвои и листьев
Вредители хвои и листьев Насекомые. Тип питания
Насекомые. Тип питания Высшие споровые растения: мхи
Высшие споровые растения: мхи Лекция 3
Лекция 3 Происхождение и эволюция Млекопитающих в свете современных представлений
Происхождение и эволюция Млекопитающих в свете современных представлений Департамент декоративного кролиководства
Департамент декоративного кролиководства Анатомия. Строение органов
Анатомия. Строение органов Организм человека как биологическая система
Организм человека как биологическая система Интерактивная игра Букварь природы
Интерактивная игра Букварь природы