- Структура, свойства и функции ферментов. Лекция №1

Содержание
- 2. ФЕРМЕНТЫ (ЭНЗИМЫ) – ВЕЩЕСТВА БЕЛКОВОЙ ПРИРОДЫ, СПОСОБНЫЕ КАТАЛИТИЧЕСКИ УСКОРЯТЬ ПРОТЕКАНИЕ ХИМИЧЕСКИХ РЕАКЦИЙ Модель фермента нуклеозидфосфорилазы
- 3. Значение для изучения: Ферменты - биологические регуляторы химических процессов в клетке (основа жизнедеятельности) Нарушения в их
- 4. Сходство ферментов с небиологическими катализаторами заключается в том, что: Небиологические катализаторы и энзимы ускоряют энергетически возможные
- 5. Изменение свободной энергии в ходе химической реакции, некатализируемой и катализируемой ферментами Известно, для того чтобы прошла
- 6. Сходство ферментов с небиологическими катализаторами заключается в том, что: В ходе катализа направление реакции не изменяется;
- 7. Отличие ферментов от небиологических катализаторов заключается в том, что: Скорость ферментативных реакций выше, чем реакций, катализируемых
- 8. Пять основных свойств ферментов как биологических катализаторов Субстратная специфичность Каталитическая эффективность Конформационная лабильность Способность ферментов к
- 9. Структура ферментов Фермент (энзим) Простой белок (РНК-аза, пищевые ферменты) Сложный белок (холофермент) Белковая часть (апофермент –
- 10. Небелковая часть (кофактор) Простетическая группа ФАД ФМН ПФ (Вит. В6) Кофермент НАД+ НАДФ+ HS-KoA Н4-фолат
- 11. Небелковая часть Производные витаминов Гемы, входящие в состав цитохромов, каталазы, пероксидазы, гуанилатциклазы, NO-синтазы и являющиеся простетической
- 12. Функции небелковой части фермента Перенос атомов водорода, электронов (окислительно-восстановительные реакции – НАД, ФАД и др.). Перенос
- 13. Активный центр ферментов Активный центр ферментов - это определенный участок белковой молекулы, представленный уникальной комбинацией аминокислотных
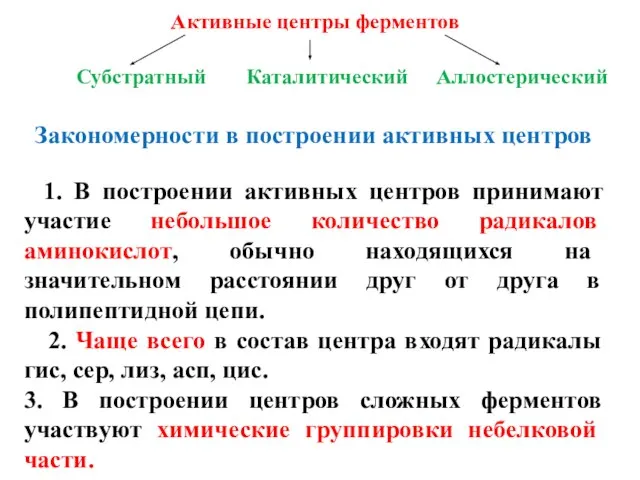
- 14. Активные центры ферментов Субстратный Каталитический Аллостерический Закономерности в построении активных центров 1. В построении активных центров
- 15. Активные центры ферментов Субстратный Каталитический Аллостерический Закономерности в построении активных центров 4. Если фермент является олиго-
- 16. Активный центр фермента Участок связывания Каталитический участок Обеспечивает субстратную специфичность (выбор субстрата) -Абсолютная субстратная специфичность -Групповая
- 17. АБСОЛЮТНАЯ СУБСТРАТНАЯ СПЕЦИФИЧНОСТЬ - активный центр фермента комплементарен только одному субстрату Мочевина
- 18. ГРУППОВАЯ СУБСТРАТНАЯ СПЕЦИФИЧНОСТЬ - фермент катализирует однотипную реакцию с небольшим количеством (группой) структурно похожих субстратов. Гидролиз
- 19. СТЕРЕОСПЕЦИФИЧНОСТЬ - фермент проявляет абсолютную специфичность только к одному из существующих стереоизомеров субстрата Стереоспецифичность к D-сахарам
- 20. Каталитические пути превращения глюкозо-6-фосфата
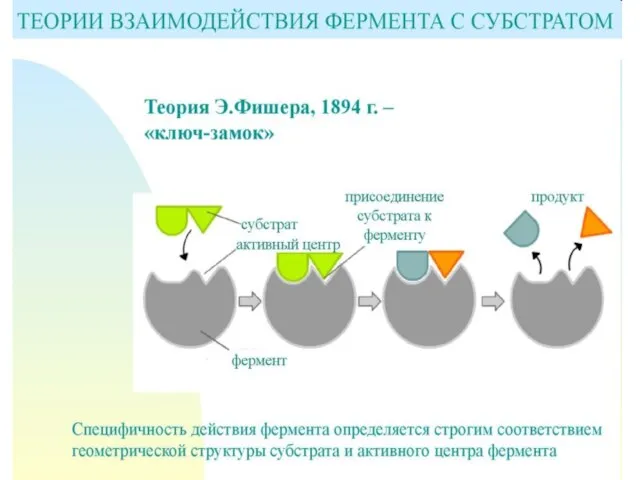
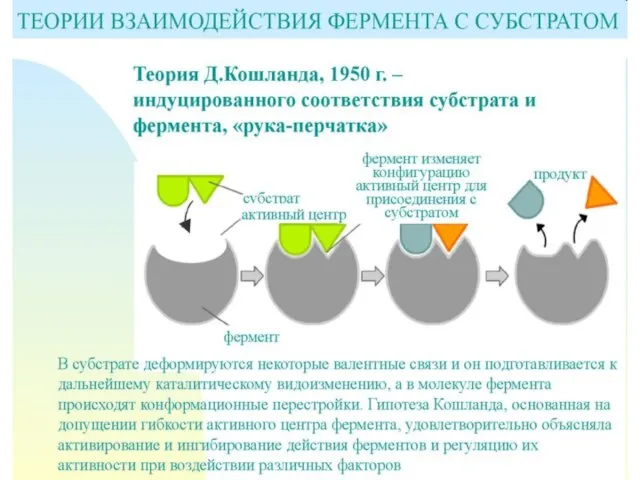
- 21. Механизмы специфичности Теория Фишера (модель "жесткой матрицы", "ключ-замок") – активный центр фермента строго соответствует конфигурации субстрата
- 23. Этапы ферментативного катализа I - этап сближения и ориентации субстрата в активном центре фермента; II -
- 24. Факторы, влияющие на активность ферментов Концентрация фермента (энзима) [E] Концентрация субстрата [S] Температура Концентрация ионов Н
- 25. Влияние [E] [S] - const 1. Рассмотрим зависимость скорости реакции от количества фермента. При условии избытка
- 26. График Михаэлиса - Ментен При высоких концентрациях субстрата наступает насыщение фермента субстратом, то есть наступает такой
- 27. Кm (константа Михаэлиса) – это такая концентрация субстрата, при которой скорость ферментативной реакции равна половине максимальной.
- 28. Уравнение Михаэлиса - Ментен где Vmax — максимальная скорость реакции; Km — константа Михаэлиса, равная концентрации
- 29. Значение Km 1. Показывает сродство [E] и [S]
- 30. Например, этиленгликоль – составная часть антифриза и алкогольдегидрогеназа (АДГ) будет превращать его в щавелевую кислоту, которая
- 31. Значение Km 2. Km – важная количественная характеристика фермента, определяет с каким [S] будет наиболее эффективно
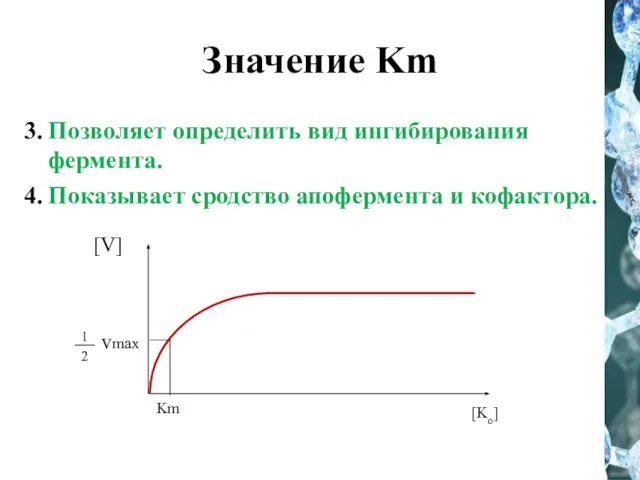
- 32. Значение Km 3. Позволяет определить вид ингибирования фермента. 4. Показывает сродство апофермента и кофактора.
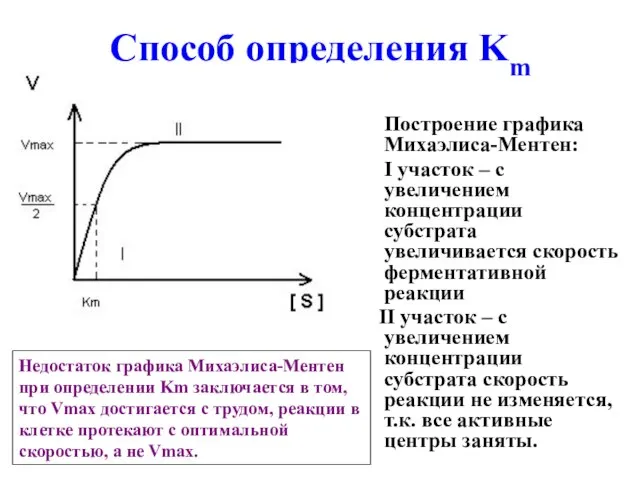
- 33. Способ определения Km Построение графика Михаэлиса-Ментен: I участок – с увеличением концентрации субстрата увеличивается скорость ферментативной
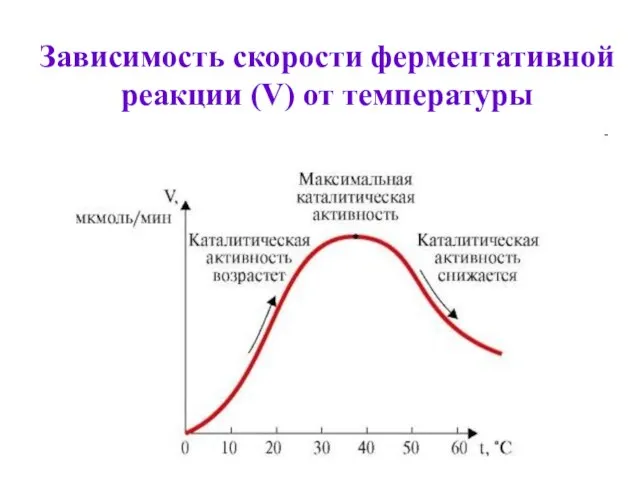
- 34. Зависимость скорости ферментативной реакции от температуры При t=36-380С ферменты обладают наибольшей активностью. Эта температура называется температурный
- 35. Зависимость скорости ферментативной реакции (V) от температуры
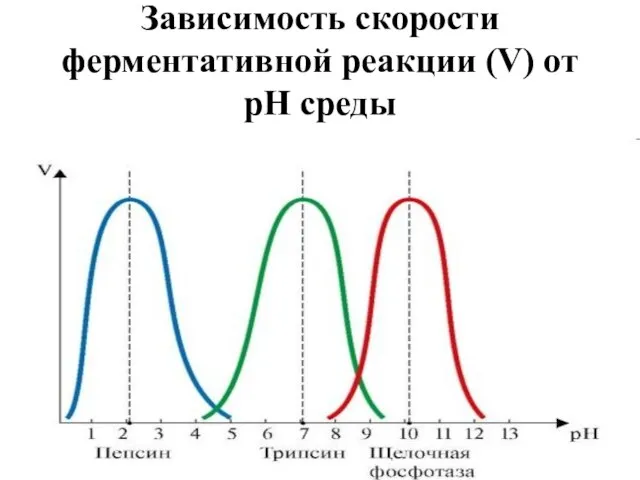
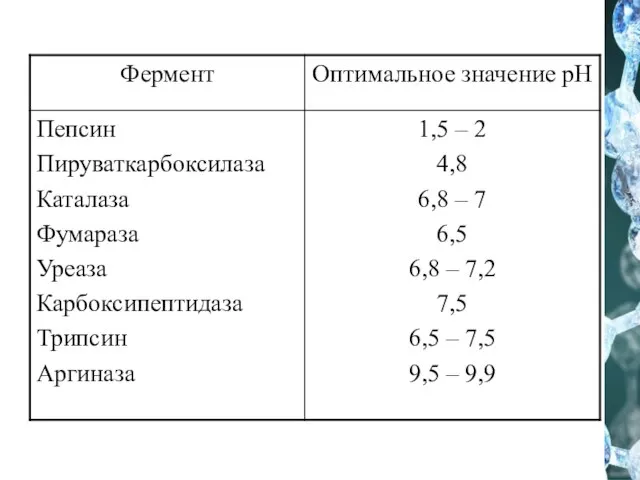
- 36. Зависимость скорости ферментативной реакции (V) от рН среды
- 38. Классификация и номенклатура ферментов
- 41. 1. Оксидоредуктазы катализируют различные окислительно-восстановительные реакции. Класс делится на подклассы: а) дегидрогеназы катализируют реакции дегидрирования (отщепления
- 42. 1. Оксидоредуктазы б) оксидазы - катализируют реакции окисления с участием молекулярного кислорода
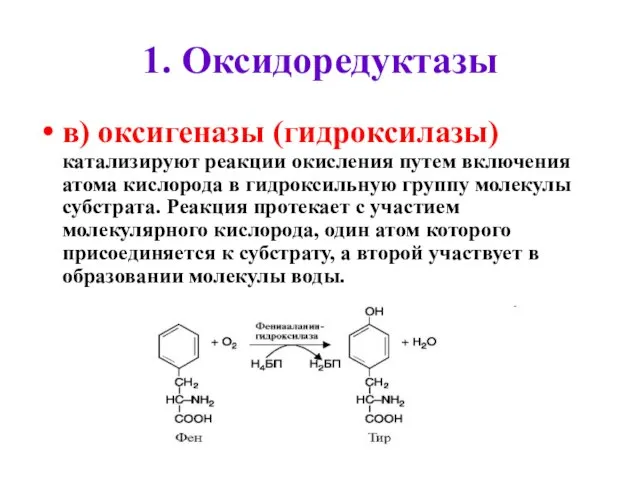
- 43. 1. Оксидоредуктазы в) оксигеназы (гидроксилазы) катализируют реакции окисления путем включения атома кислорода в гидроксильную группу молекулы
- 44. 2. Трансферазы - катализируют реакции переноса функциональных групп. В зависимости от переносимой группы подразделяются на подклассы:
- 45. 3. Гидролазы катализируют реакции гидролиза (расщепления ковалентной связи с присоединением молекулы воды по месту разрыва). Разделяются
- 46. 4. Лиазы К лиазам относятся ферменты, отщепляющие от субстратов негидролитическим путем определенные группы, такие, как СО2,
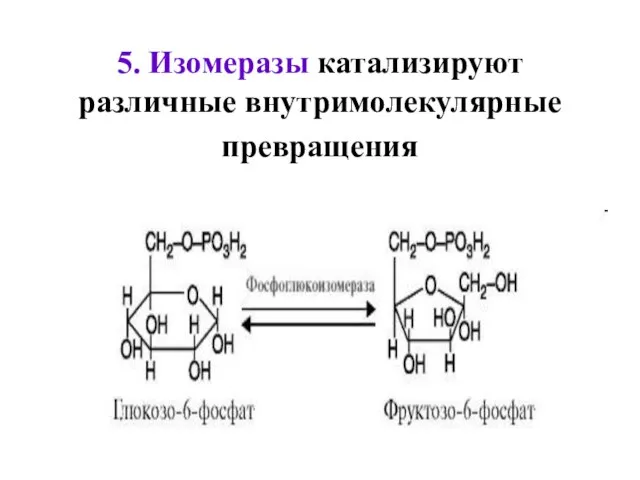
- 47. 5. Изомеразы катализируют различные внутримолекулярные превращения
- 49. Скачать презентацию
Слайд 2ФЕРМЕНТЫ (ЭНЗИМЫ) – ВЕЩЕСТВА БЕЛКОВОЙ ПРИРОДЫ, СПОСОБНЫЕ КАТАЛИТИЧЕСКИ УСКОРЯТЬ ПРОТЕКАНИЕ ХИМИЧЕСКИХ РЕАКЦИЙ
Модель
ФЕРМЕНТЫ (ЭНЗИМЫ) – ВЕЩЕСТВА БЕЛКОВОЙ ПРИРОДЫ, СПОСОБНЫЕ КАТАЛИТИЧЕСКИ УСКОРЯТЬ ПРОТЕКАНИЕ ХИМИЧЕСКИХ РЕАКЦИЙ
Модель

Слайд 3Значение для изучения:
Ферменты - биологические регуляторы химических процессов в клетке (основа жизнедеятельности)
Нарушения
Значение для изучения:
Ферменты - биологические регуляторы химических процессов в клетке (основа жизнедеятельности)
Нарушения

Слайд 4Сходство ферментов с небиологическими катализаторами заключается в том, что:
Небиологические катализаторы и энзимы
Сходство ферментов с небиологическими катализаторами заключается в том, что:
Небиологические катализаторы и энзимы

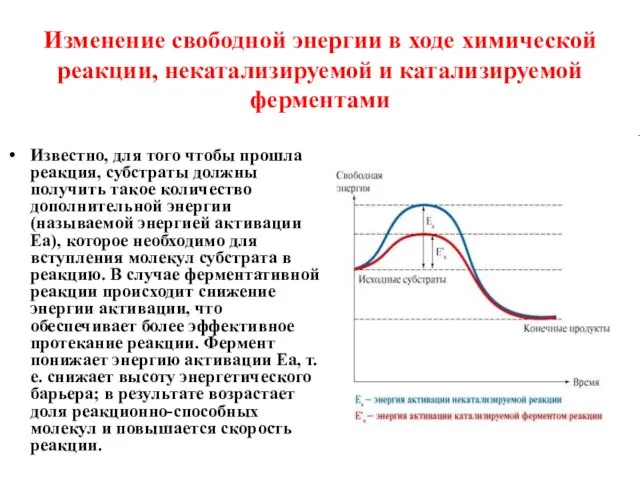
Слайд 5Изменение свободной энергии в ходе химической реакции, некатализируемой и катализируемой ферментами
Известно, для
Изменение свободной энергии в ходе химической реакции, некатализируемой и катализируемой ферментами
Известно, для
Слайд 6Сходство ферментов с небиологическими катализаторами заключается в том, что:
В ходе катализа направление
Сходство ферментов с небиологическими катализаторами заключается в том, что:
В ходе катализа направление

Слайд 7Отличие ферментов от небиологических катализаторов заключается в том, что:
Скорость ферментативных реакций выше,
Отличие ферментов от небиологических катализаторов заключается в том, что:
Скорость ферментативных реакций выше,

Слайд 8Пять основных свойств ферментов
как биологических катализаторов
Субстратная специфичность
Каталитическая эффективность
Конформационная лабильность
Способность ферментов к
Пять основных свойств ферментов
как биологических катализаторов
Субстратная специфичность
Каталитическая эффективность
Конформационная лабильность
Способность ферментов к

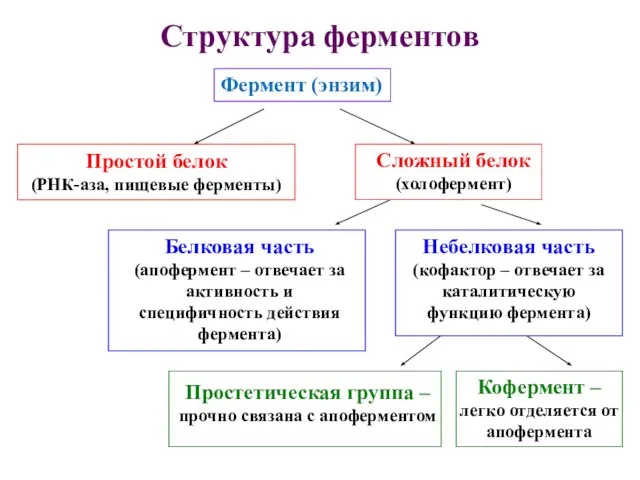
Слайд 9Структура ферментов
Фермент (энзим)
Простой белок
(РНК-аза, пищевые ферменты)
Сложный белок
(холофермент)
Белковая часть (апофермент – отвечает
Структура ферментов
Фермент (энзим)
Простой белок
(РНК-аза, пищевые ферменты)
Сложный белок
(холофермент)
Белковая часть (апофермент – отвечает
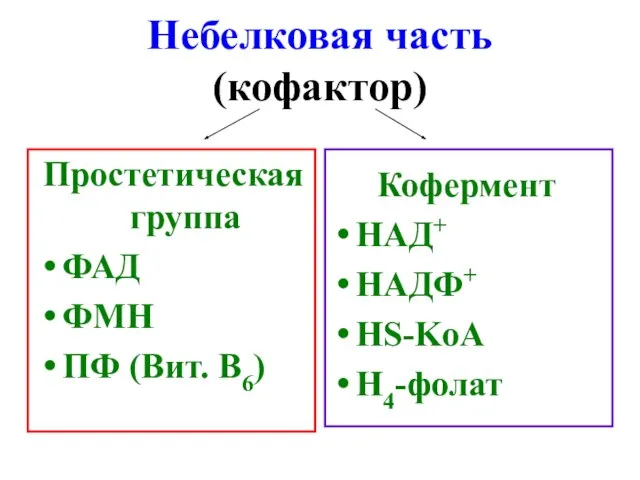
Слайд 10Небелковая часть
(кофактор)
Простетическая группа
ФАД
ФМН
ПФ (Вит. В6)
Кофермент
НАД+
НАДФ+
HS-KoA
Н4-фолат
Небелковая часть
(кофактор)
Простетическая группа
ФАД
ФМН
ПФ (Вит. В6)
Кофермент
НАД+
НАДФ+
HS-KoA
Н4-фолат
Слайд 11Небелковая часть
Производные витаминов
Гемы, входящие в состав цитохромов, каталазы, пероксидазы, гуанилатциклазы, NO-синтазы и
Небелковая часть
Производные витаминов
Гемы, входящие в состав цитохромов, каталазы, пероксидазы, гуанилатциклазы, NO-синтазы и

Слайд 12Функции небелковой части фермента
Перенос атомов водорода, электронов (окислительно-восстановительные реакции – НАД, ФАД
Функции небелковой части фермента
Перенос атомов водорода, электронов (окислительно-восстановительные реакции – НАД, ФАД

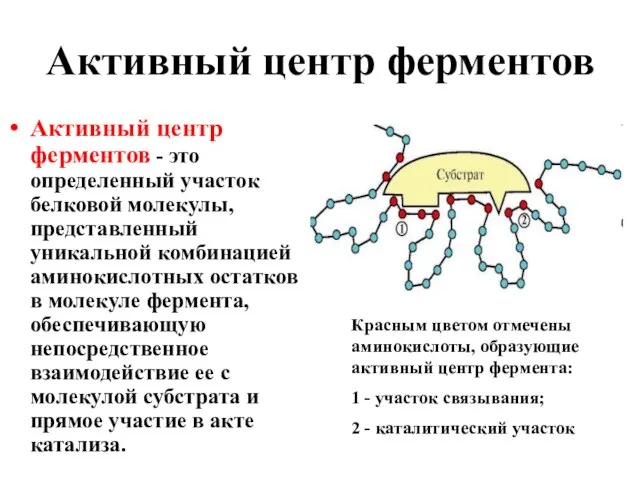
Слайд 13Активный центр ферментов
Активный центр ферментов - это определенный участок белковой молекулы, представленный
Активный центр ферментов
Активный центр ферментов - это определенный участок белковой молекулы, представленный
Слайд 14Активные центры ферментов
Субстратный
Каталитический
Аллостерический
Закономерности в построении активных центров
1. В построении активных
Активные центры ферментов
Субстратный
Каталитический
Аллостерический
Закономерности в построении активных центров
1. В построении активных
Слайд 15Активные центры ферментов
Субстратный
Каталитический
Аллостерический
Закономерности в построении активных центров
4. Если фермент является
Активные центры ферментов
Субстратный
Каталитический
Аллостерический
Закономерности в построении активных центров
4. Если фермент является

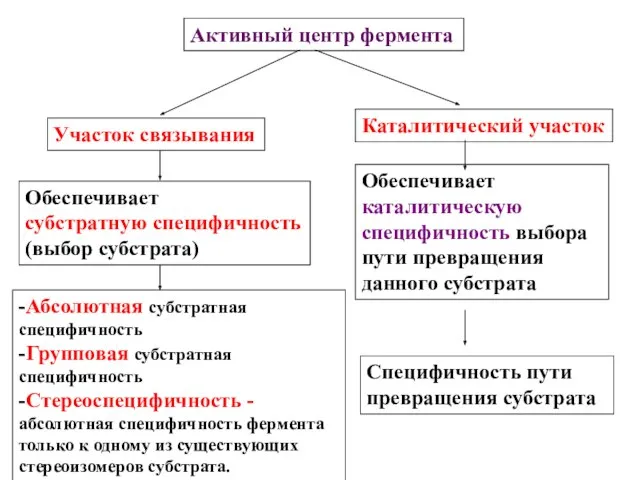
Слайд 16Активный центр фермента
Участок связывания
Каталитический участок
Обеспечивает
субстратную специфичность
(выбор субстрата)
-Абсолютная субстратная специфичность
-Групповая субстратная специфичность
-Стереоспецифичность
Активный центр фермента
Участок связывания
Каталитический участок
Обеспечивает
субстратную специфичность
(выбор субстрата)
-Абсолютная субстратная специфичность
-Групповая субстратная специфичность
-Стереоспецифичность
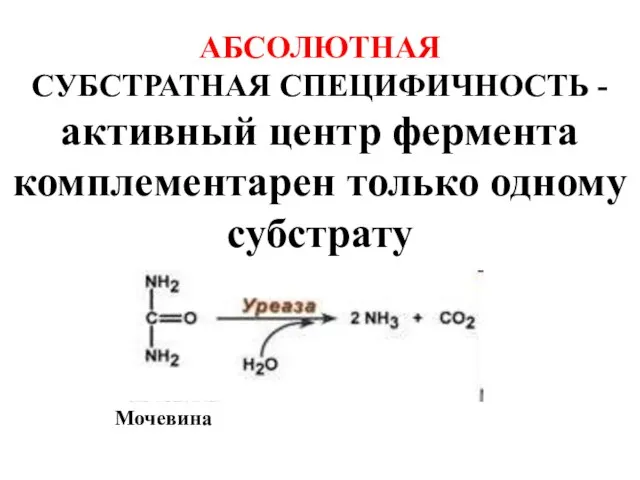
Слайд 17АБСОЛЮТНАЯ
СУБСТРАТНАЯ СПЕЦИФИЧНОСТЬ - активный центр фермента комплементарен только одному субстрату
Мочевина
АБСОЛЮТНАЯ
СУБСТРАТНАЯ СПЕЦИФИЧНОСТЬ - активный центр фермента комплементарен только одному субстрату
Мочевина
Слайд 18ГРУППОВАЯ
СУБСТРАТНАЯ СПЕЦИФИЧНОСТЬ - фермент катализирует однотипную реакцию с небольшим количеством (группой)
ГРУППОВАЯ СУБСТРАТНАЯ СПЕЦИФИЧНОСТЬ - фермент катализирует однотипную реакцию с небольшим количеством (группой)

Слайд 19СТЕРЕОСПЕЦИФИЧНОСТЬ - фермент проявляет абсолютную специфичность только к одному из существующих стереоизомеров
СТЕРЕОСПЕЦИФИЧНОСТЬ - фермент проявляет абсолютную специфичность только к одному из существующих стереоизомеров

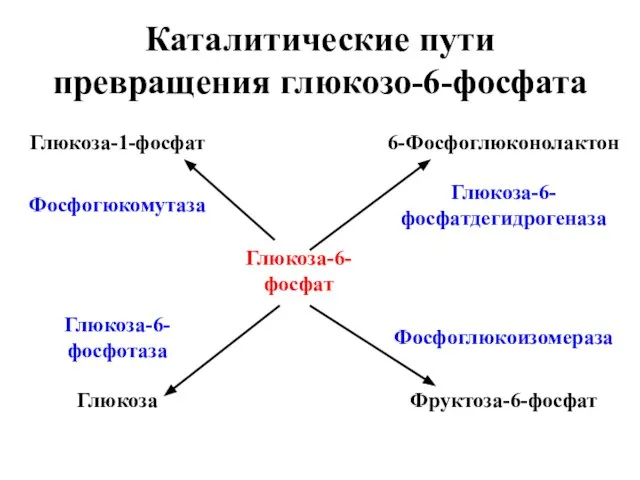
Слайд 20Каталитические пути превращения глюкозо-6-фосфата
Каталитические пути превращения глюкозо-6-фосфата
Слайд 21Механизмы специфичности
Теория Фишера (модель "жесткой матрицы", "ключ-замок") – активный центр фермента строго
Механизмы специфичности
Теория Фишера (модель "жесткой матрицы", "ключ-замок") – активный центр фермента строго
Слайд 23Этапы ферментативного катализа
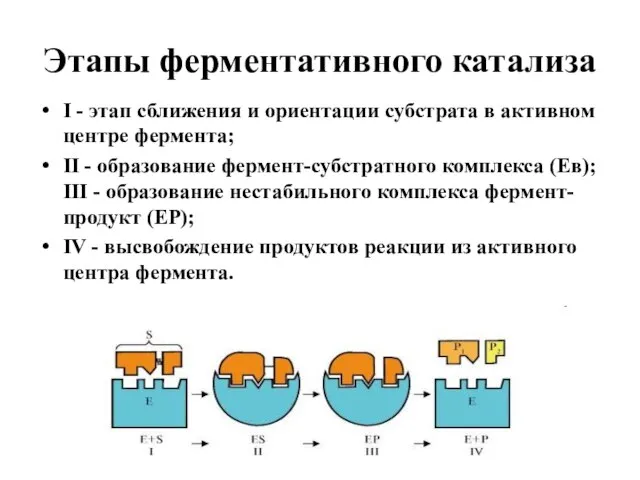
I - этап сближения и ориентации субстрата в активном центре
Этапы ферментативного катализа
I - этап сближения и ориентации субстрата в активном центре
Слайд 24Факторы, влияющие
на активность ферментов
Концентрация фермента (энзима) [E]
Концентрация субстрата [S]
Температура
Концентрация ионов Н (рН)
Низкомолекулярные
Факторы, влияющие
на активность ферментов
Концентрация фермента (энзима) [E]
Концентрация субстрата [S]
Температура
Концентрация ионов Н (рН)
Низкомолекулярные
![Факторы, влияющие на активность ферментов Концентрация фермента (энзима) [E] Концентрация субстрата [S]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/907273/slide-23.jpg)
Слайд 25Влияние [E]
[S] - const
1. Рассмотрим зависимость скорости реакции от
количества фермента. При
Влияние [E]
[S] - const
1. Рассмотрим зависимость скорости реакции от
количества фермента. При
![Влияние [E] [S] - const 1. Рассмотрим зависимость скорости реакции от количества](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/907273/slide-24.jpg)
Слайд 26График Михаэлиса - Ментен
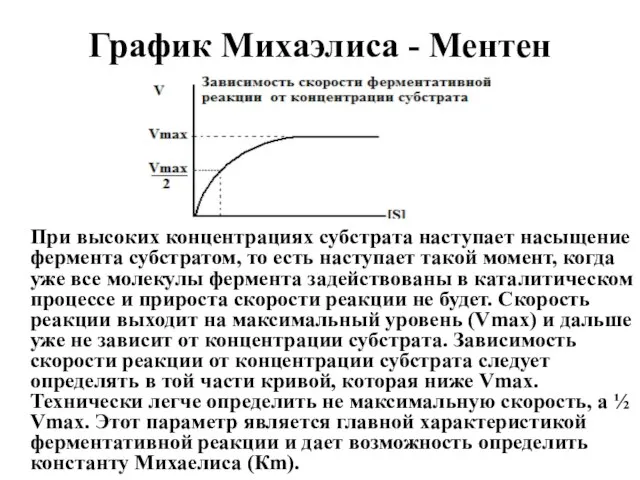
При высоких концентрациях субстрата наступает насыщение фермента субстратом, то
График Михаэлиса - Ментен
При высоких концентрациях субстрата наступает насыщение фермента субстратом, то
Слайд 27Кm (константа Михаэлиса) – это такая
концентрация субстрата, при которой скорость ферментативной реакции
Кm (константа Михаэлиса) – это такая
концентрация субстрата, при которой скорость ферментативной реакции

Слайд 28Уравнение Михаэлиса - Ментен
где
Vmax — максимальная скорость реакции;
Km — константа Михаэлиса, равная
Уравнение Михаэлиса - Ментен
где
Vmax — максимальная скорость реакции;
Km — константа Михаэлиса, равная

Слайд 29Значение Km
1. Показывает сродство [E] и [S]
Значение Km
1. Показывает сродство [E] и [S]
![Значение Km 1. Показывает сродство [E] и [S]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/907273/slide-28.jpg)
Слайд 30Например, этиленгликоль – составная часть антифриза и алкогольдегидрогеназа (АДГ) будет превращать его
Например, этиленгликоль – составная часть антифриза и алкогольдегидрогеназа (АДГ) будет превращать его

Слайд 31Значение Km
2. Km – важная количественная характеристика фермента, определяет с каким [S]
Значение Km
2. Km – важная количественная характеристика фермента, определяет с каким [S]

Слайд 32Значение Km
3. Позволяет определить вид ингибирования фермента.
4. Показывает сродство апофермента и кофактора.
Значение Km
3. Позволяет определить вид ингибирования фермента.
4. Показывает сродство апофермента и кофактора.
Слайд 33Способ определения Km
Построение графика Михаэлиса-Ментен:
I участок – с увеличением
Способ определения Km
Построение графика Михаэлиса-Ментен:
I участок – с увеличением
Слайд 34Зависимость скорости ферментативной реакции от температуры
При t=36-380С ферменты обладают наибольшей активностью. Эта
Зависимость скорости ферментативной реакции от температуры
При t=36-380С ферменты обладают наибольшей активностью. Эта

Слайд 35Зависимость скорости ферментативной реакции (V) от температуры
Зависимость скорости ферментативной реакции (V) от температуры
Слайд 36Зависимость скорости ферментативной реакции (V) от рН среды
Зависимость скорости ферментативной реакции (V) от рН среды
Слайд 38Классификация и номенклатура ферментов
Классификация и номенклатура ферментов


Слайд 411. Оксидоредуктазы катализируют различные окислительно-восстановительные реакции. Класс делится на подклассы:
а) дегидрогеназы катализируют
1. Оксидоредуктазы катализируют различные окислительно-восстановительные реакции. Класс делится на подклассы:
а) дегидрогеназы катализируют

Слайд 421. Оксидоредуктазы
б) оксидазы - катализируют реакции окисления с участием молекулярного кислорода
1. Оксидоредуктазы
б) оксидазы - катализируют реакции окисления с участием молекулярного кислорода

Слайд 431. Оксидоредуктазы
в) оксигеназы (гидроксилазы) катализируют реакции окисления путем включения атома кислорода в
1. Оксидоредуктазы
в) оксигеназы (гидроксилазы) катализируют реакции окисления путем включения атома кислорода в
Слайд 442. Трансферазы - катализируют реакции переноса функциональных групп.
В зависимости от переносимой группы
2. Трансферазы - катализируют реакции переноса функциональных групп.
В зависимости от переносимой группы

Слайд 453. Гидролазы катализируют реакции гидролиза (расщепления ковалентной связи с присоединением молекулы воды
3. Гидролазы катализируют реакции гидролиза (расщепления ковалентной связи с присоединением молекулы воды

Слайд 464. Лиазы
К лиазам относятся ферменты, отщепляющие от субстратов негидролитическим путем определенные группы,
4. Лиазы
К лиазам относятся ферменты, отщепляющие от субстратов негидролитическим путем определенные группы,

Слайд 475. Изомеразы катализируют различные внутримолекулярные превращения
5. Изомеразы катализируют различные внутримолекулярные превращения
 Зоологическая азбука
Зоологическая азбука Что такое биология?
Что такое биология? Видоизменения побегов. 6 класс
Видоизменения побегов. 6 класс Хищники. Программа Путешествие по планете Земля
Хищники. Программа Путешествие по планете Земля zritelny_analizator_prezintatsia
zritelny_analizator_prezintatsia Голосеменные Покрытосеменные растения
Голосеменные Покрытосеменные растения Sprache und Gehirn. Anatomische Grundlagen
Sprache und Gehirn. Anatomische Grundlagen Водный обмен. Значение воды в организме человека
Водный обмен. Значение воды в организме человека Приспособления организмов к жизни в природе (5 класс)
Приспособления организмов к жизни в природе (5 класс) Презентация на тему ОБЩИЙ ОБЗОР ОРГАНИЗМА ЧЕЛОВЕКА
Презентация на тему ОБЩИЙ ОБЗОР ОРГАНИЗМА ЧЕЛОВЕКА  Строение листа растения
Строение листа растения Введение, предмет и задачи медицинской микробиологии и иммунологии. Классификация микроорганизмов
Введение, предмет и задачи медицинской микробиологии и иммунологии. Классификация микроорганизмов Жизнь в океане и на суше
Жизнь в океане и на суше Создание портативных камер для культивирования животных клеток
Создание портативных камер для культивирования животных клеток Принципы транскрипции. Структура промотора прокариот
Принципы транскрипции. Структура промотора прокариот Место человека в системе органического мира
Место человека в системе органического мира Витамин Д
Витамин Д Юрский период
Юрский период Орган слуха и равновесия
Орган слуха и равновесия Sınıf CEPHALOPODA
Sınıf CEPHALOPODA Презентация на тему Класс Птицы
Презентация на тему Класс Птицы  Обучение и сон
Обучение и сон Домашние птицы и их детеныши
Домашние птицы и их детеныши Расстройства речи (невропатологический аспект)
Расстройства речи (невропатологический аспект) Жизнь без мутаций
Жизнь без мутаций Презентация на тему Осторожно: клещи!
Презентация на тему Осторожно: клещи!  Паразитизм. Классификация паразитов и форм паразитизма
Паразитизм. Классификация паразитов и форм паразитизма Мозг и нервная система человека
Мозг и нервная система человека