- Таламус. Ядра таламуса

Содержание
- 2. Таламус
- 3. Таламус делится прослойками белого вещества на три области: переднюю, латеральную и медиальную
- 5. Таламус Сложно организованная структура, содержащая очень большое количество нейронных скоплений и участвующая в реализации множества различных
- 6. Ядра таламуса Аксоны нервных клеток некоторых ядер таламуса могут заканчиваться не только в одной, но и
- 7. У млекопитающих происходит усиленное развитие таламуса и формируется таламонеокортикальная система интеграции
- 8. Таламус Коллектор всех афферентных (сенсорных) путей (за исключением обонятельных), идущих к большим полушариям. Это как бы
- 9. Классификация ядер таламуса Учитывая морфологические различия, характер проекции в кору, организацию афферентных путей и функциональные особенности,
- 10. Специфические ядра таламуса Сенсорные Моторные Лимбические
- 11. Специфические ядра таламуса 1. Имеют локальную проекцию в строго определенные участки коры. 2. Моносинаптически связаны аксосоматическими
- 12. Специфические ядра таламуса 4. Основную массу клеток составляют «релейные» нейроны с небольшим диаметром тела, длинным аксоном
- 13. Специфические ядра таламуса 5. Одиночное раздражение соответствующей рецепторной области вызывает в специфическом сенсорном ядре первичный электрический
- 14. Сенсорные ядра латеральные коленчатые тела медиальные коленчатые тела вентробазальный комплекс разрушение - полное необратимое выпадение определенной
- 16. Соматотопическая организация VPM VPL Коже лица соответствуют нейронные группировки в медиальной части ядра, коже туловища —
- 20. Медиальные коленчатые тела предкорковый уровень анализа слуховых сигналов Проекции в 41 и 42 поля коры Тонотопия
- 22. Латеральные коленчатые тела Проекция центральной ямки сетчатки представлена наиболее широко и локализована в верхних слоях, проекция
- 23. Рецептивные поля сетчатки в таламусе у хищников и приматов, имеют концентрическую форму и состоят из центральной
- 26. Моторные ядра В этих ядрах переключается афферентация, идущая от ядер мозжечка и бледного шара к моторной
- 28. Лимбические ядра Переключение импульсов из мамиллярных тел гипоталамуса в лимбическую кору; Ведущая афферентная система лимбических ядер
- 30. Ассоциативные ядра таламуса Ассоциативные ядра таламуса наиболее поздно дифференцируются и интенсивно развиваются в ходе эволюции
- 31. Ассоциативные ядра таламуса 1. Эфферентные волокна направляются к ассоциативным областям коры, а также частично и к
- 32. Ассоциативные ядра таламуса 4. Основную массу клеток составляют нейроны 3 типов: а) мультиполярные клетки с 4—6
- 33. Ассоциативные ядра таламуса 5.Электрическое раздражение ассоциативных ядер вызывает в ассоциативных областях коры вторичные ассоциативные ответы, которые
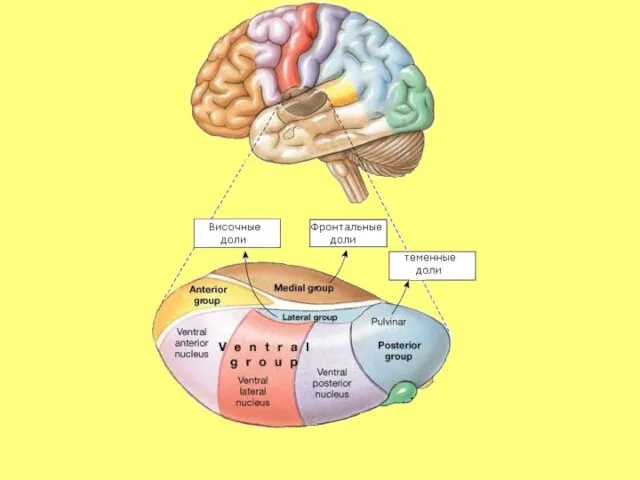
- 34. Ассоциативные ядра таламуса К ним относятся: MD, LP и подушка
- 36. Ассоциативные ядра таламуса В MD преобладают реакции нейронов на соматическую афферентацию в подушке — на зрительную.
- 37. Ассоциативные ядра таламуса Важной особенностью нейронов ассоциативных ядер является способность многих из них реагировать на раздражения
- 38. Ассоциативные ядра таламуса Среди нейронов ассоциативных ядер таламуса есть такие, которые реагируют только на комплексные раздражения
- 39. Таламокортикальная ассоциативная система Таламофронтальная Таламопариетальная Таламотемпоральная
- 40. Неспецифические ядра таламуса морфологически и функционально связаны со многими системами и участвуют вместе с ретикулярной формацией
- 41. Неспецифические ядра таламуса 1. Проецируются на кору больших полушарий более диффузно, чем специфические ядра. 2. Связь
- 42. Неспецифические ядра таламуса 4. Нейронные сети имеют преимущественно «ретикулярное» строение, т.е. состоят, в основном, из густой
- 43. Неспецифические ядра таламуса Неспецифические ядра таламуса оказывают на кору головного мозга модулирующее влияние, регулируют ее функциональное
- 44. Неспецифические ядра таламуса Патологические процессы в этих структурах у человека или их повреждения во время операций
- 45. Неспецифические ядра таламуса На основании электрофизиологических экспериментов Г. Джаспер пришел к выводу, что неспецифическая система таламуса
- 46. Взаимодействие таламокортикальных систем Системы находятся в постоянном взаимодействии Неспецифическая система усиливает специфическую, Специфическая, наоборот, подавляет неспецифическую.
- 47. Взаимодействие таламокортикальных систем Под влиянием неспецифических импульсов реакция корковых нейронов на специфическую стимуляцию заметно усиливается, т.е.
- 48. В нейронных сетях и специфических, и неспецифических ядер таламуса происходят сложные интегративные процессы, связанные с переработкой
- 50. Кроме передачи проекционных влияний на кору, нейронные ядра таламуса могут сами осуществлять замыкание рефлекторных путей без
- 51. Удаление коры и подкорковых ядер У таких «таламических» животных сохраняются не только простые движения, но и
- 52. Таламус является высшим центром болевой чувствительности Импульсы, идущие к нейронам таламуса от поврежденных участков тела и
- 53. Таламус является высшим центром болевой чувствительности Ощущение боли связано с возбуждением нейронов неспецифических ядер таламуса; для
- 54. Таламические интегративные механизмы имеют большое значение для УР деятельности, формирования эмоциональных реакций и мотивационного поведения. Оказывая
- 55. эпифиз
- 58. Эпиталамус (надталамическая область) Шишковидное тело - эпифиз, которое при помощи поводков соединяется с медиальными поверхностями правого
- 60. Скачать презентацию
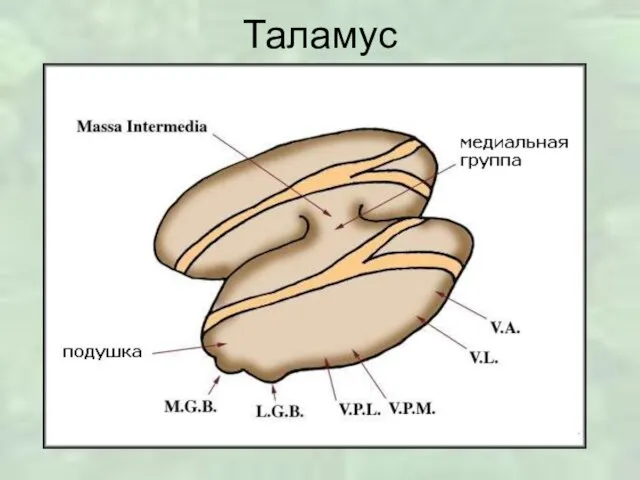
Слайд 3Таламус
делится прослойками белого вещества на три области: переднюю, латеральную и медиальную
Таламус
делится прослойками белого вещества на три области: переднюю, латеральную и медиальную
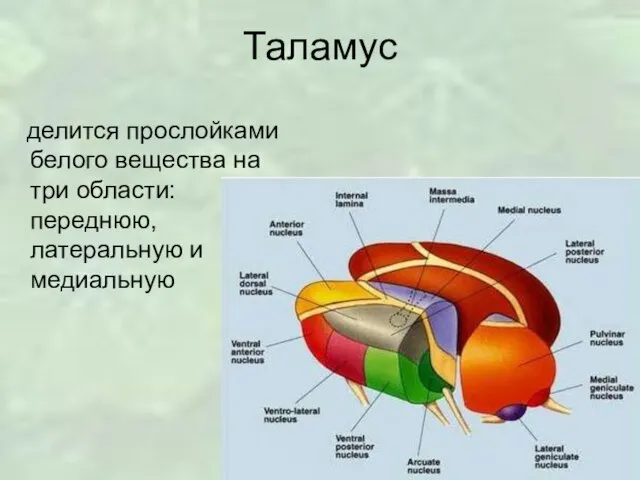
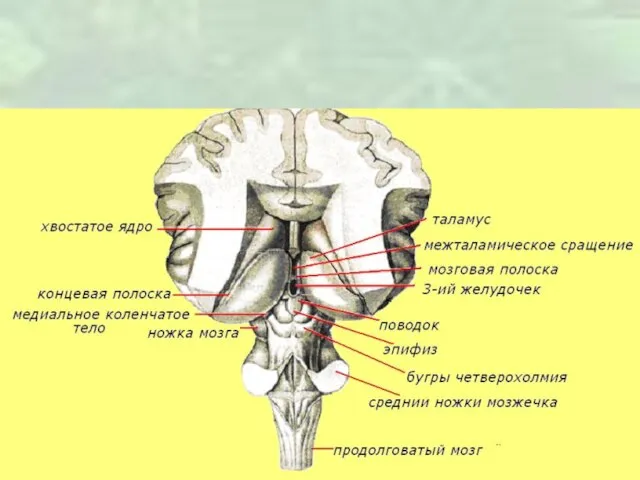
Слайд 5Таламус
Сложно организованная структура, содержащая очень большое количество нейронных скоплений и участвующая
Таламус
Сложно организованная структура, содержащая очень большое количество нейронных скоплений и участвующая

Слайд 6Ядра таламуса
Аксоны нервных клеток некоторых ядер таламуса могут заканчиваться не только
Ядра таламуса
Аксоны нервных клеток некоторых ядер таламуса могут заканчиваться не только

Слайд 7У млекопитающих происходит усиленное развитие таламуса
и формируется таламонеокортикальная система интеграции
У млекопитающих происходит усиленное развитие таламуса
и формируется таламонеокортикальная система интеграции

Слайд 8Таламус
Коллектор всех афферентных (сенсорных) путей (за исключением обонятельных), идущих к большим
Таламус
Коллектор всех афферентных (сенсорных) путей (за исключением обонятельных), идущих к большим

Слайд 9Классификация ядер таламуса
Учитывая морфологические различия, характер проекции в кору, организацию афферентных
Классификация ядер таламуса
Учитывая морфологические различия, характер проекции в кору, организацию афферентных

Слайд 10Специфические ядра таламуса
Сенсорные
Моторные
Лимбические
Специфические ядра таламуса
Сенсорные
Моторные
Лимбические

Слайд 11Специфические ядра таламуса
1. Имеют локальную проекцию в строго определенные участки коры.
2. Моносинаптически
Специфические ядра таламуса
1. Имеют локальную проекцию в строго определенные участки коры.
2. Моносинаптически

Слайд 12Специфические ядра таламуса
4. Основную массу клеток составляют «релейные» нейроны с небольшим диаметром
Специфические ядра таламуса
4. Основную массу клеток составляют «релейные» нейроны с небольшим диаметром

Слайд 13Специфические ядра таламуса
5. Одиночное раздражение соответствующей рецепторной области вызывает в специфическом сенсорном
Специфические ядра таламуса
5. Одиночное раздражение соответствующей рецепторной области вызывает в специфическом сенсорном

Слайд 14Сенсорные ядра
латеральные коленчатые тела
медиальные коленчатые тела
вентробазальный комплекс
разрушение - полное необратимое выпадение
Сенсорные ядра
латеральные коленчатые тела
медиальные коленчатые тела
вентробазальный комплекс
разрушение - полное необратимое выпадение

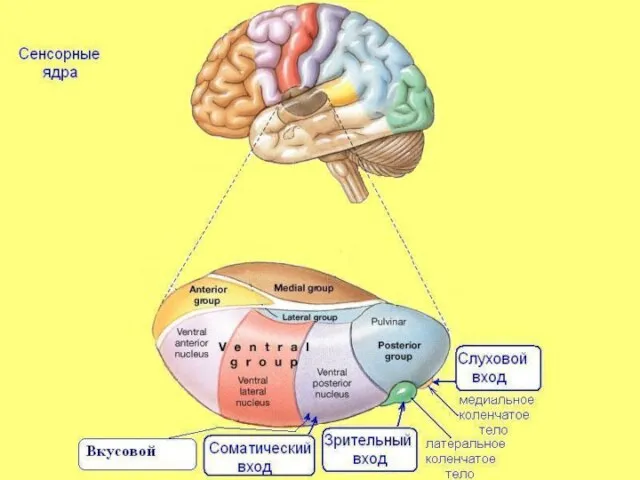
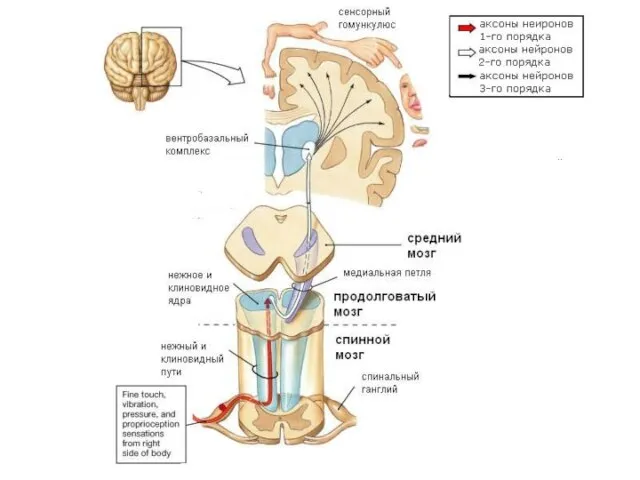
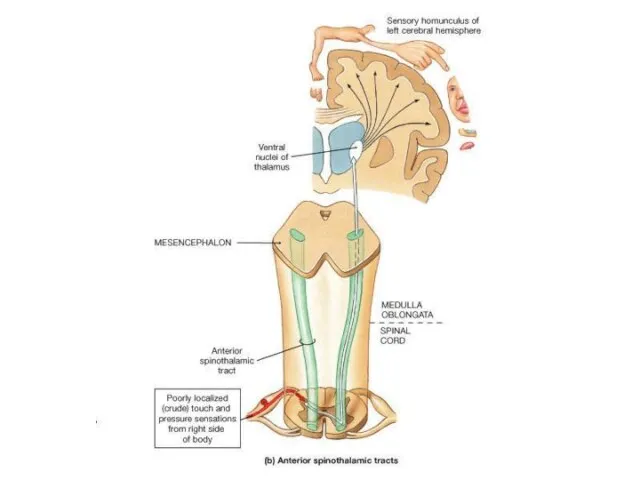
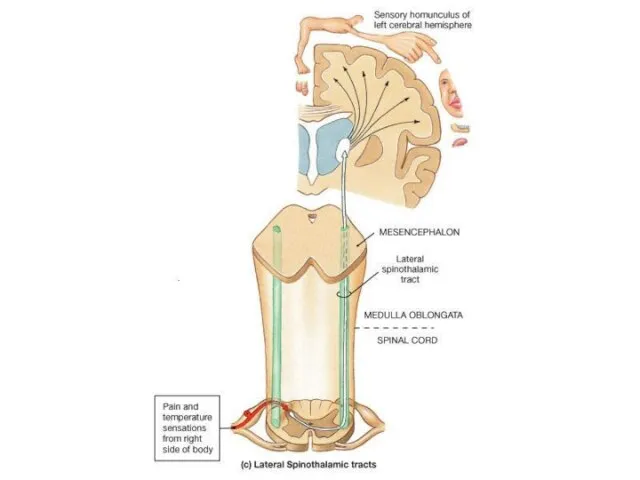
Слайд 16Соматотопическая организация VPM VPL
Коже лица соответствуют нейронные группировки в медиальной части
Соматотопическая организация VPM VPL
Коже лица соответствуют нейронные группировки в медиальной части

Слайд 20Медиальные коленчатые тела
предкорковый уровень анализа слуховых сигналов
Проекции в 41 и
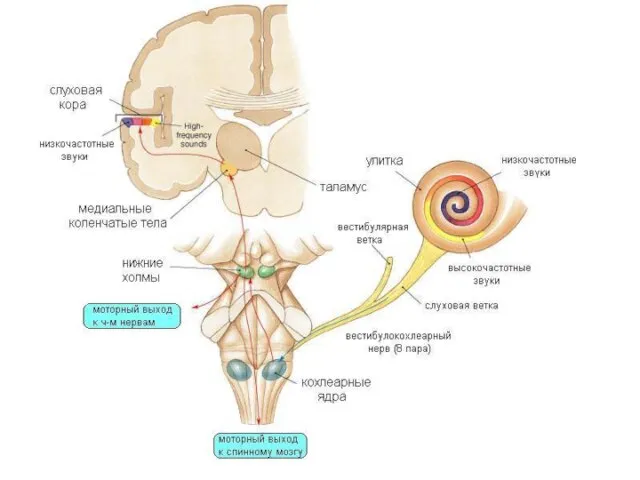
Медиальные коленчатые тела
предкорковый уровень анализа слуховых сигналов
Проекции в 41 и

Слайд 22Латеральные коленчатые тела
Проекция центральной ямки сетчатки представлена наиболее широко и локализована
Латеральные коленчатые тела
Проекция центральной ямки сетчатки представлена наиболее широко и локализована

Слайд 23Рецептивные поля сетчатки в таламусе у хищников и приматов, имеют концентрическую форму
Рецептивные поля сетчатки в таламусе у хищников и приматов, имеют концентрическую форму

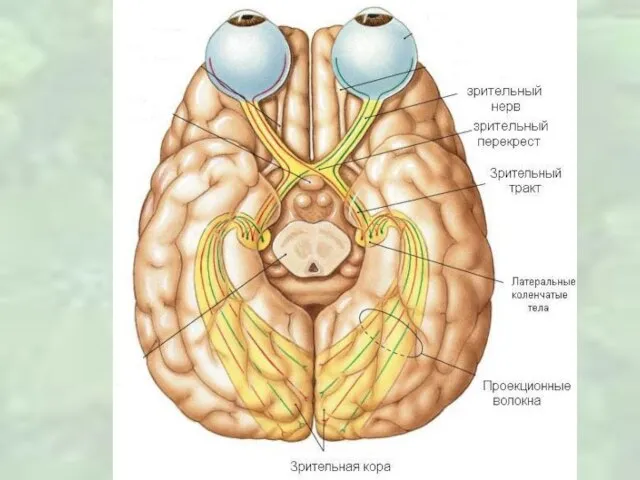
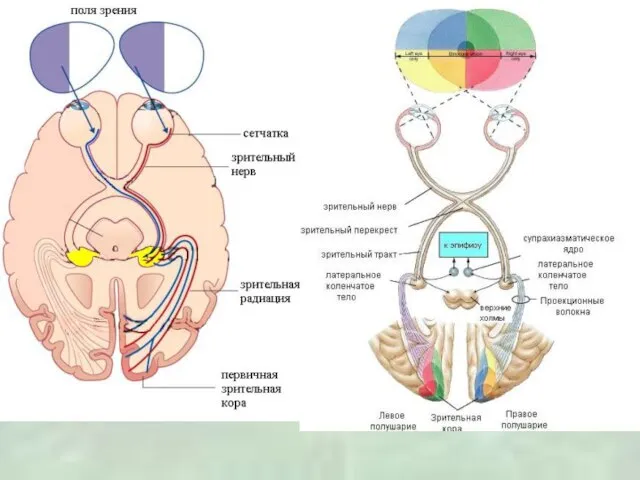
Слайд 26Моторные ядра
В этих ядрах переключается афферентация, идущая от ядер мозжечка и бледного
Моторные ядра
В этих ядрах переключается афферентация, идущая от ядер мозжечка и бледного

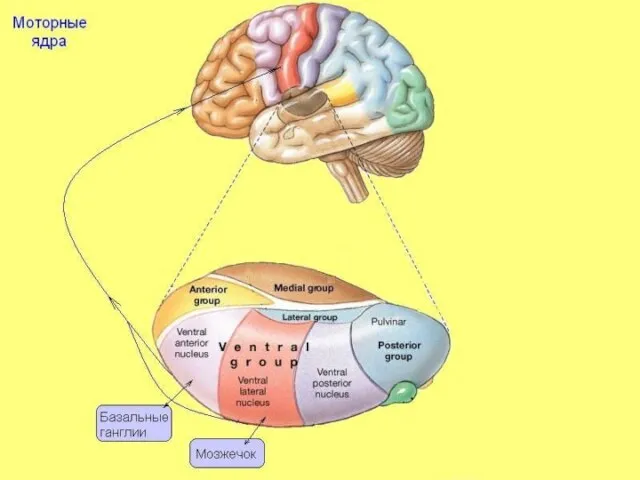
Слайд 28Лимбические ядра
Переключение импульсов из мамиллярных тел гипоталамуса в лимбическую кору;
Ведущая афферентная
Лимбические ядра
Переключение импульсов из мамиллярных тел гипоталамуса в лимбическую кору;
Ведущая афферентная

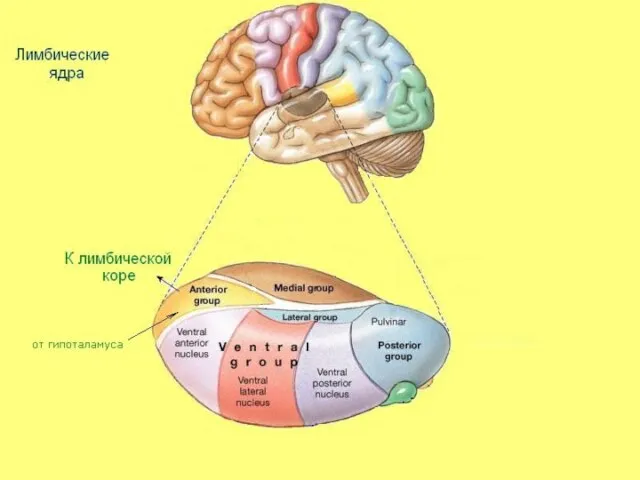
Слайд 30Ассоциативные ядра таламуса
Ассоциативные ядра таламуса наиболее поздно дифференцируются и интенсивно развиваются
Ассоциативные ядра таламуса
Ассоциативные ядра таламуса наиболее поздно дифференцируются и интенсивно развиваются

Слайд 31Ассоциативные ядра таламуса
1. Эфферентные волокна направляются к ассоциативным областям коры, а также
Ассоциативные ядра таламуса
1. Эфферентные волокна направляются к ассоциативным областям коры, а также

Слайд 32Ассоциативные ядра таламуса
4. Основную массу клеток составляют нейроны 3 типов:
а) мультиполярные
Ассоциативные ядра таламуса
4. Основную массу клеток составляют нейроны 3 типов:
а) мультиполярные

Слайд 33Ассоциативные ядра таламуса
5.Электрическое раздражение ассоциативных ядер вызывает в ассоциативных областях коры вторичные
Ассоциативные ядра таламуса
5.Электрическое раздражение ассоциативных ядер вызывает в ассоциативных областях коры вторичные

Слайд 34Ассоциативные ядра таламуса
К ним относятся: MD, LP и подушка
Ассоциативные ядра таламуса
К ним относятся: MD, LP и подушка

Слайд 36Ассоциативные ядра таламуса
В MD преобладают реакции нейронов на соматическую афферентацию
Ассоциативные ядра таламуса
В MD преобладают реакции нейронов на соматическую афферентацию

Слайд 37Ассоциативные ядра таламуса
Важной особенностью нейронов ассоциативных ядер является способность многих из них
Ассоциативные ядра таламуса
Важной особенностью нейронов ассоциативных ядер является способность многих из них

Слайд 38Ассоциативные ядра таламуса
Среди нейронов ассоциативных ядер таламуса есть такие, которые реагируют
Ассоциативные ядра таламуса
Среди нейронов ассоциативных ядер таламуса есть такие, которые реагируют

Слайд 39Таламокортикальная ассоциативная система
Таламофронтальная
Таламопариетальная
Таламотемпоральная
Таламокортикальная ассоциативная система
Таламофронтальная
Таламопариетальная
Таламотемпоральная

Слайд 40Неспецифические ядра таламуса
морфологически и функционально связаны со многими системами и
Неспецифические ядра таламуса
морфологически и функционально связаны со многими системами и

Слайд 41Неспецифические ядра таламуса
1. Проецируются на кору больших полушарий более диффузно, чем
Неспецифические ядра таламуса
1. Проецируются на кору больших полушарий более диффузно, чем

Слайд 42Неспецифические ядра таламуса
4. Нейронные сети имеют преимущественно «ретикулярное» строение, т.е. состоят,
Неспецифические ядра таламуса
4. Нейронные сети имеют преимущественно «ретикулярное» строение, т.е. состоят,

Слайд 43Неспецифические ядра таламуса
Неспецифические ядра таламуса оказывают на кору головного мозга модулирующее влияние,
Неспецифические ядра таламуса
Неспецифические ядра таламуса оказывают на кору головного мозга модулирующее влияние,

Слайд 44Неспецифические ядра таламуса
Патологические процессы в этих структурах у человека или их повреждения
Неспецифические ядра таламуса
Патологические процессы в этих структурах у человека или их повреждения

Слайд 45Неспецифические ядра таламуса
На основании электрофизиологических экспериментов Г. Джаспер пришел к выводу, что
Неспецифические ядра таламуса
На основании электрофизиологических экспериментов Г. Джаспер пришел к выводу, что

Слайд 46Взаимодействие таламокортикальных систем
Системы находятся в постоянном взаимодействии
Неспецифическая система усиливает специфическую,
Взаимодействие таламокортикальных систем
Системы находятся в постоянном взаимодействии
Неспецифическая система усиливает специфическую,

Слайд 47Взаимодействие таламокортикальных систем
Под влиянием неспецифических импульсов реакция корковых нейронов на специфическую стимуляцию
Взаимодействие таламокортикальных систем
Под влиянием неспецифических импульсов реакция корковых нейронов на специфическую стимуляцию

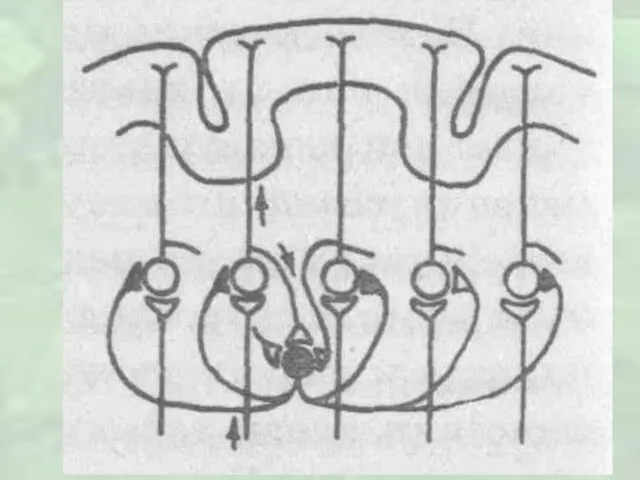
Слайд 48В нейронных сетях и специфических, и неспецифических ядер таламуса происходят сложные интегративные
В нейронных сетях и специфических, и неспецифических ядер таламуса происходят сложные интегративные

Слайд 50Кроме передачи проекционных влияний на кору, нейронные ядра таламуса могут сами осуществлять
Кроме передачи проекционных влияний на кору, нейронные ядра таламуса могут сами осуществлять

Слайд 51Удаление коры и подкорковых ядер
У таких «таламических» животных сохраняются не только простые
Удаление коры и подкорковых ядер
У таких «таламических» животных сохраняются не только простые

Слайд 52Таламус является высшим центром болевой чувствительности
Импульсы, идущие к нейронам таламуса от
Таламус является высшим центром болевой чувствительности
Импульсы, идущие к нейронам таламуса от

Слайд 53Таламус является высшим центром болевой чувствительности
Ощущение боли связано с возбуждением нейронов неспецифических
Таламус является высшим центром болевой чувствительности
Ощущение боли связано с возбуждением нейронов неспецифических

Слайд 54Таламические интегративные механизмы имеют большое значение для УР деятельности, формирования эмоциональных реакций
Таламические интегративные механизмы имеют большое значение для УР деятельности, формирования эмоциональных реакций

Слайд 55эпифиз
эпифиз


Слайд 58Эпиталамус
(надталамическая область)
Шишковидное тело - эпифиз, которое при помощи поводков соединяется с
Эпиталамус
(надталамическая область)
Шишковидное тело - эпифиз, которое при помощи поводков соединяется с

 Позвоночный столб
Позвоночный столб Брюхоногие моллюски Выполнил : Севостьянов Дмитрий
Брюхоногие моллюски Выполнил : Севостьянов Дмитрий Разбор заданий ЕГЭ по биологии
Разбор заданий ЕГЭ по биологии Астра однолетняя
Астра однолетняя Удивительные растения
Удивительные растения Белок кинезин
Белок кинезин Нарцисс
Нарцисс Высотная поясность
Высотная поясность Дефекты зрения
Дефекты зрения Экологические системы
Экологические системы Иннервация тазового дна, синдром Алкока, синдром запирательного нерва
Иннервация тазового дна, синдром Алкока, синдром запирательного нерва Анатомическое строение листа
Анатомическое строение листа Высшие, или плацентарные звери: насекомоядные и рукокрылые, грызуны и зайцеобразные, хищные
Высшие, или плацентарные звери: насекомоядные и рукокрылые, грызуны и зайцеобразные, хищные Кошечки. Породы кошек
Кошечки. Породы кошек Покрытосеменные растения. Урок обобщения, повторения и контроля
Покрытосеменные растения. Урок обобщения, повторения и контроля Популяции. Изменчивость популяции
Популяции. Изменчивость популяции Біологічні основи запліднення та розвитку організму
Біологічні основи запліднення та розвитку організму Опасные растения и насекомые Крыма
Опасные растения и насекомые Крыма Сцепленное наследование и типы взаимодействия генов
Сцепленное наследование и типы взаимодействия генов Класс земноводные. Amphibia
Класс земноводные. Amphibia Джозеф Редьярд Кіплінг (1865-1936)
Джозеф Редьярд Кіплінг (1865-1936) Презентация на тему Глаз как орган зрения и оптическая система
Презентация на тему Глаз как орган зрения и оптическая система  Фотосинтез. Общая схема фотосинтеза
Фотосинтез. Общая схема фотосинтеза История открытия пенициллина
История открытия пенициллина Многообразие животного мира
Многообразие животного мира Презентация на тему Змеи
Презентация на тему Змеи  Общие закономерности физиологии и ее основные понятия
Общие закономерности физиологии и ее основные понятия Органы чувств. Орган слуха и равновесия
Органы чувств. Орган слуха и равновесия