- Трансформация энергии на биомембранах

Содержание
- 2. Трансформация энергии на биомембранах: Перенос электронов и запасание энергии Фотобиологические процессы Процессы рецепции Сократительные системы
- 3. РЕЦЕПЦИЯ
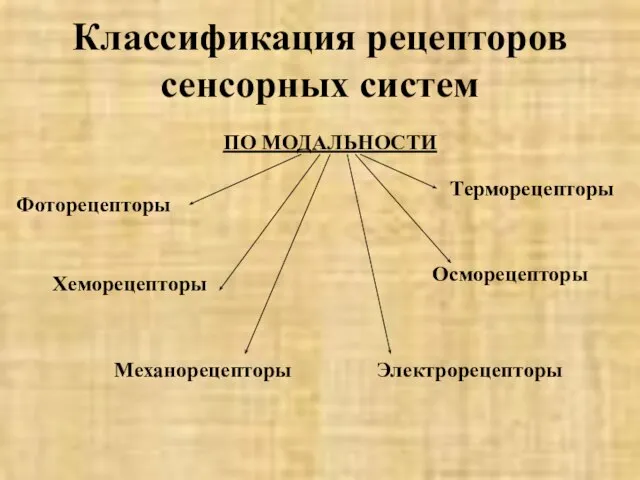
- 4. Классификация рецепторов сенсорных систем ПО МОДАЛЬНОСТИ Фоторецепторы Хеморецепторы Терморецепторы Механорецепторы Осморецепторы Электрорецепторы
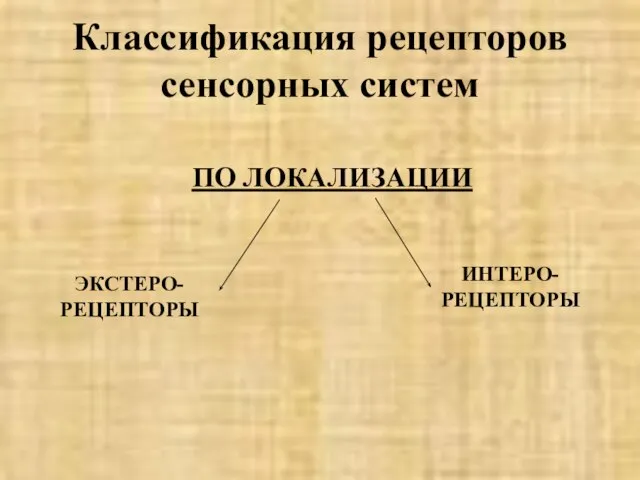
- 5. Классификация рецепторов сенсорных систем ПО ЛОКАЛИЗАЦИИ ЭКСТЕРО-РЕЦЕПТОРЫ ИНТЕРО-РЕЦЕПТОРЫ
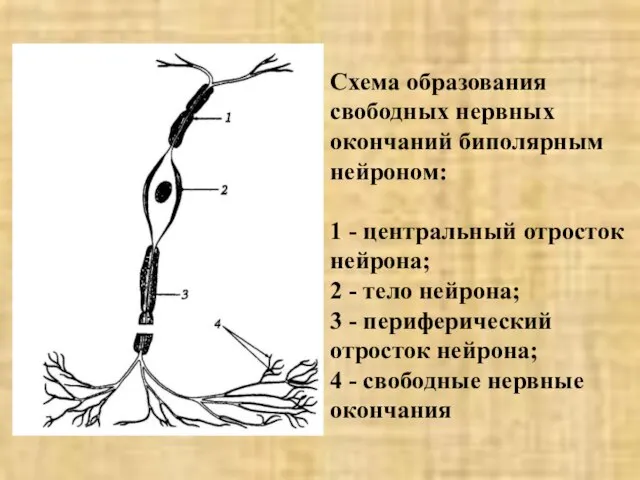
- 6. Схема образования свободных нервных окончаний биполярным нейроном: 1 - центральный отросток нейрона; 2 - тело нейрона;
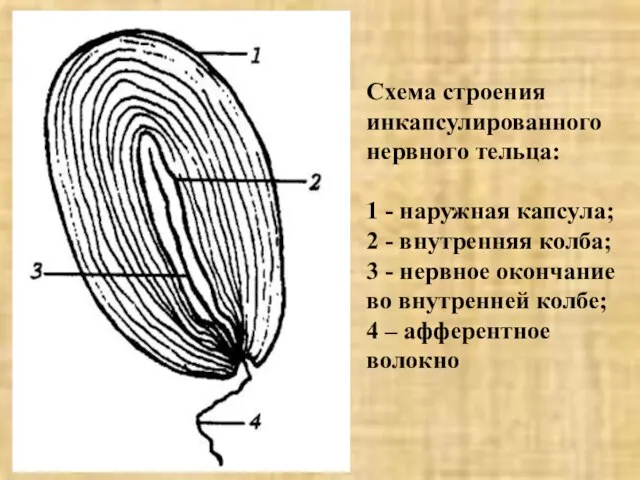
- 7. Схема строения инкапсулированного нервного тельца: 1 - наружная капсула; 2 - внутренняя колба; 3 - нервное
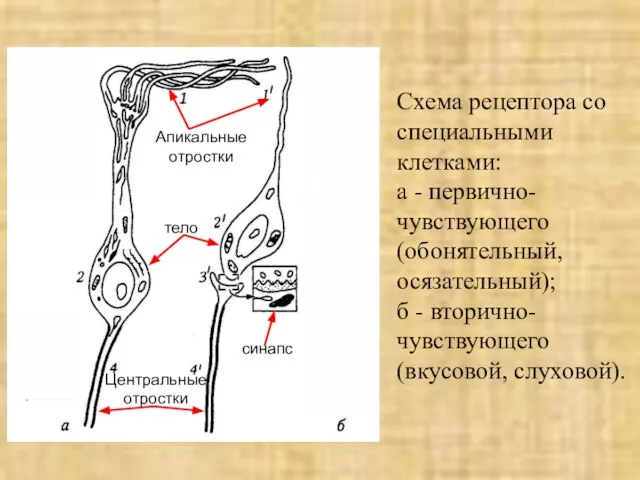
- 8. Схема рецептора со специальными клетками: а - первично-чувствующего (обонятельный, осязательный); б - вторично-чувствующего (вкусовой, слуховой). Апикальные
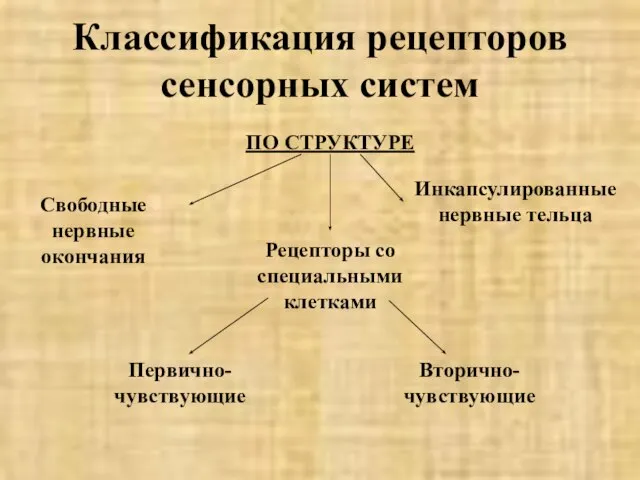
- 9. Классификация рецепторов сенсорных систем ПО СТРУКТУРЕ Свободные нервные окончания Первично-чувствующие Инкапсулированные нервные тельца Рецепторы со специальными
- 10. БИОФИЗИЧЕСКИЕ МЕХАНИЗМЫ ПРЕОБРАЗОВАНИЯ ИНФОРМАЦИИ В РЕЦЕПТОРАХ Независимо от структуры и особенностей функционирования, все рецепторы в принципе
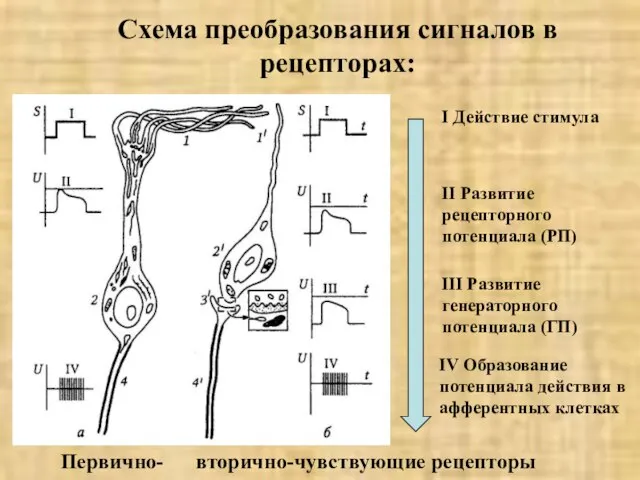
- 11. Схема преобразования сигналов в рецепторах: I Действие стимула II Развитие рецепторного потенциала (РП) III Развитие генераторного
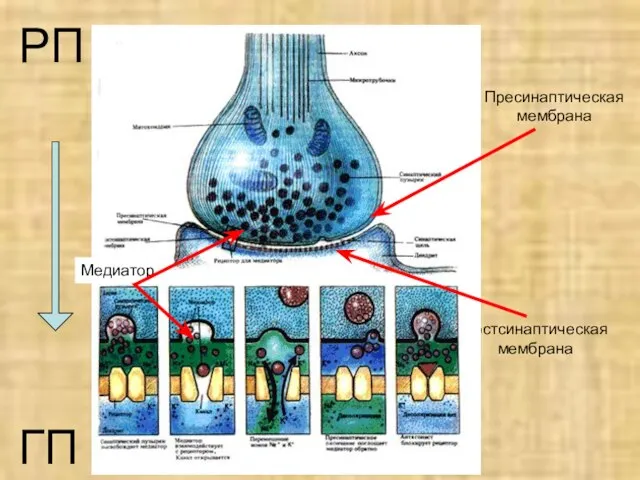
- 12. Пресинаптическая мембрана Постсинаптическая мембрана РП ГП Медиатор
- 15. Передача сигнала в обонятельных сенсорных нейронах.
- 17. Кодирование информации в рецепторныx аппаратах В технике отображение одного сигнала другим называется кодированием. Таким образом, рецепторы
- 18. Сигнальные признаки кода 1) средняя частота импульсации; 2) число сигналов в пачке; 3) продолжительность пачек; 4)
- 19. Кодирование силы раздражителя Роль рефрактерности в преобразовании аналогового сигнала в дискретный и кодировании силы раздражителя частотой
- 20. Э. Вебер сформулировал его так: ощущение растет в арифметической прогрессии при нарастании интенсивности вызвавшего его раздражителя
- 21. Соотношение между интенсивностью раздражителя (I) и частотой импульсов в афферентном волокне (ν ) называют силовой функцией.
- 22. Графическое изображение законов Вебера-Фехнера (сверху) и Стивенса
- 23. Кодирование качества раздражителя. теория «меченой линии» Основана на принципе анатомической локализации. Содержание гипотезы: каждый рецептор и
- 24. Биофизика слуха
- 25. Схема органа слуха: А- ушная раковина; В - наружный слуховой проход; С - барабанная перепонка; D
- 26. Костная улитка: а - стержень улитки (сканирующая электронная микроскопия): б— улитка с частично вскрытой костной стенкой;
- 27. А - общий вид: 1 - лестница преддверия, 2 - барабанная лестница, 3 - вестибулярная мембрана,
- 28. Волосковая клетка — реконструкция по данным электронной микроскопии (по М. Энгштрему и И. Верселлу): 1 -
- 29. Зависимость местоположения максимального смешения базилярной мембраны человека от частоты звукового тона: а - вырезанная из улитки
- 31. Сократительные системы
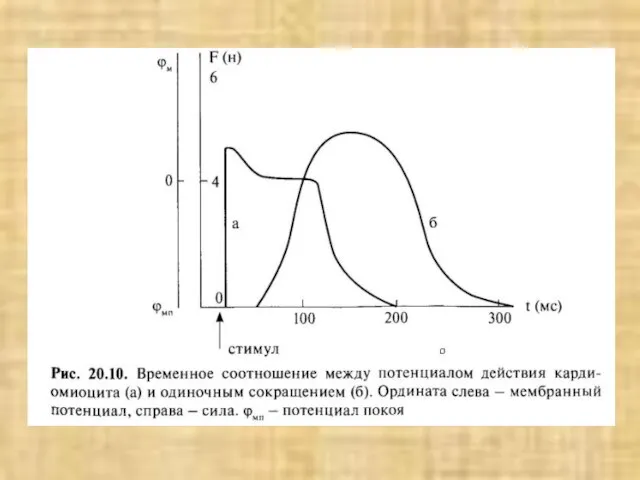
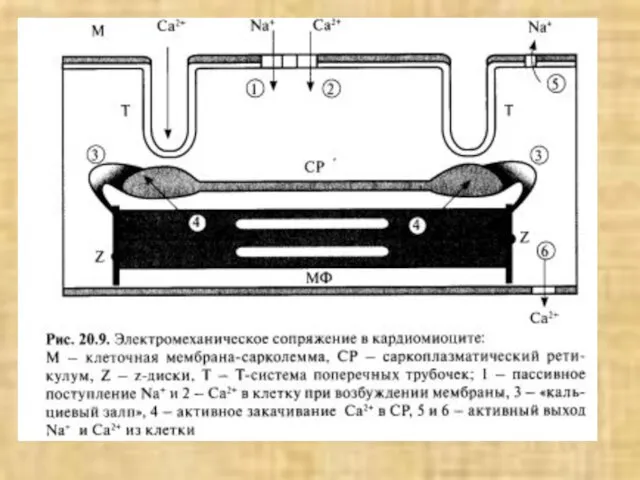
- 33. Схема структурной организации мышечной клетки (по Woledge R. et al., 1992)
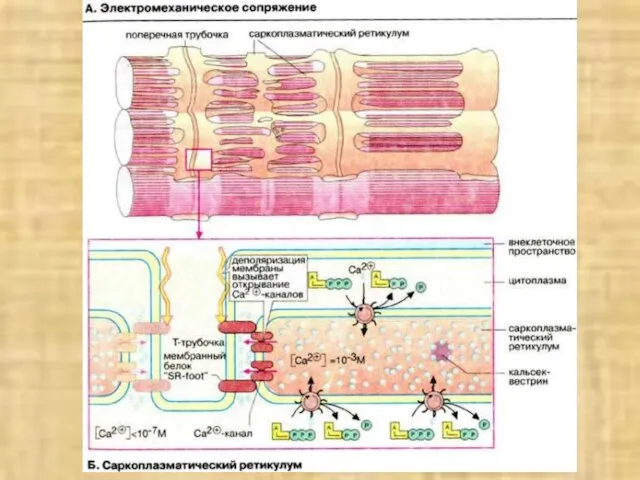
- 38. Активация мостика и генерация силы в саркомере
- 40. Основные положения модели скользящих нитей 1. Длины нитей актина и миозина в ходе сокращения не меняются.
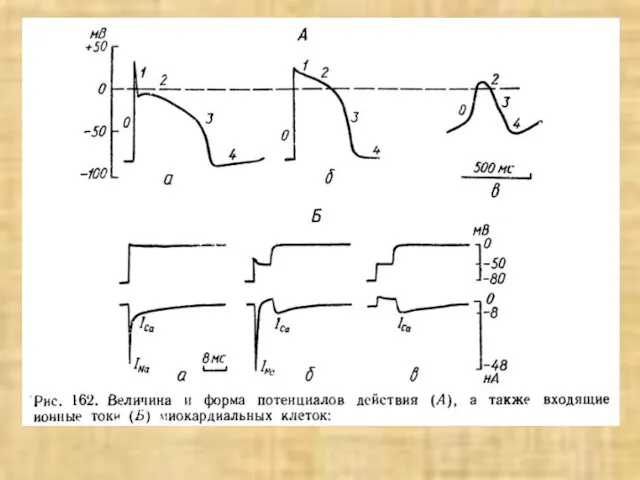
- 42. Типы миокардиальных клеток: а - мышечные клетки рабочего миокарда, б - проводящие волокна (волокна Пуркинье); в
- 48. Скачать презентацию
Слайд 2Трансформация энергии на биомембранах:
Перенос электронов и запасание энергии
Фотобиологические процессы
Процессы рецепции
Сократительные системы
Трансформация энергии на биомембранах:
Перенос электронов и запасание энергии
Фотобиологические процессы
Процессы рецепции
Сократительные системы

Слайд 3РЕЦЕПЦИЯ
РЕЦЕПЦИЯ

Слайд 4Классификация рецепторов сенсорных систем
ПО МОДАЛЬНОСТИ
Фоторецепторы
Хеморецепторы
Терморецепторы
Механорецепторы
Осморецепторы
Электрорецепторы
Классификация рецепторов сенсорных систем
ПО МОДАЛЬНОСТИ
Фоторецепторы
Хеморецепторы
Терморецепторы
Механорецепторы
Осморецепторы
Электрорецепторы
Слайд 5Классификация рецепторов сенсорных систем
ПО ЛОКАЛИЗАЦИИ
ЭКСТЕРО-РЕЦЕПТОРЫ
ИНТЕРО-РЕЦЕПТОРЫ
Классификация рецепторов сенсорных систем
ПО ЛОКАЛИЗАЦИИ
ЭКСТЕРО-РЕЦЕПТОРЫ
ИНТЕРО-РЕЦЕПТОРЫ
Слайд 6Схема образования свободных нервных окончаний биполярным нейроном:
1 - центральный отросток нейрона;
2
Схема образования свободных нервных окончаний биполярным нейроном:
1 - центральный отросток нейрона;
2
Слайд 7Схема строения инкапсулированного нервного тельца:
1 - наружная капсула;
2 - внутренняя колба;
Схема строения инкапсулированного нервного тельца:
1 - наружная капсула;
2 - внутренняя колба;
Слайд 8Схема рецептора со специальными клетками:
а - первично-чувствующего (обонятельный, осязательный);
б - вторично-чувствующего
Схема рецептора со специальными клетками:
а - первично-чувствующего (обонятельный, осязательный);
б - вторично-чувствующего
Слайд 9Классификация рецепторов сенсорных систем
ПО СТРУКТУРЕ
Свободные нервные окончания
Первично-чувствующие
Инкапсулированные нервные тельца
Рецепторы со специальными клетками
Вторично-чувствующие
Классификация рецепторов сенсорных систем
ПО СТРУКТУРЕ
Свободные нервные окончания
Первично-чувствующие
Инкапсулированные нервные тельца
Рецепторы со специальными клетками
Вторично-чувствующие
Слайд 10БИОФИЗИЧЕСКИЕ МЕХАНИЗМЫ ПРЕОБРАЗОВАНИЯ ИНФОРМАЦИИ В РЕЦЕПТОРАХ
Независимо от структуры и особенностей функционирования, все
БИОФИЗИЧЕСКИЕ МЕХАНИЗМЫ ПРЕОБРАЗОВАНИЯ ИНФОРМАЦИИ В РЕЦЕПТОРАХ
Независимо от структуры и особенностей функционирования, все

Слайд 11Схема преобразования сигналов в рецепторах:
I Действие стимула
II Развитие рецепторного потенциала (РП)
III Развитие
Схема преобразования сигналов в рецепторах:
I Действие стимула
II Развитие рецепторного потенциала (РП)
III Развитие
Слайд 12Пресинаптическая мембрана
Постсинаптическая мембрана
РП
ГП
Медиатор
Пресинаптическая мембрана
Постсинаптическая мембрана
РП
ГП
Медиатор


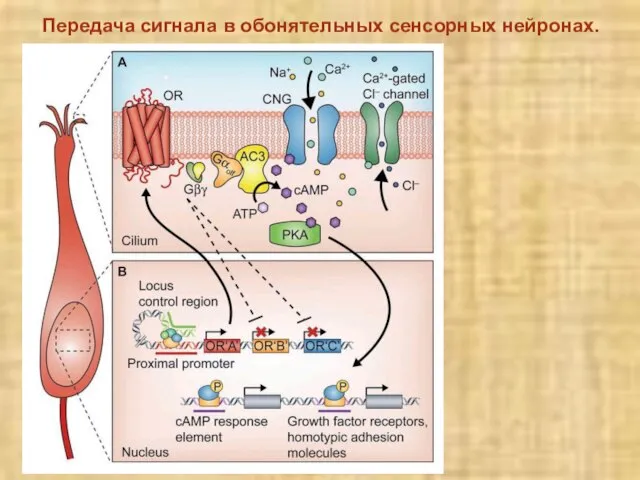
Слайд 15Передача сигнала в обонятельных сенсорных нейронах.
Передача сигнала в обонятельных сенсорных нейронах.
Слайд 17Кодирование информации в рецепторныx аппаратах
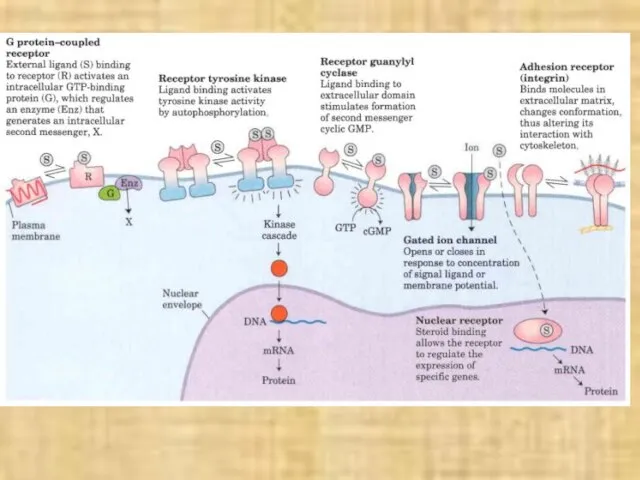
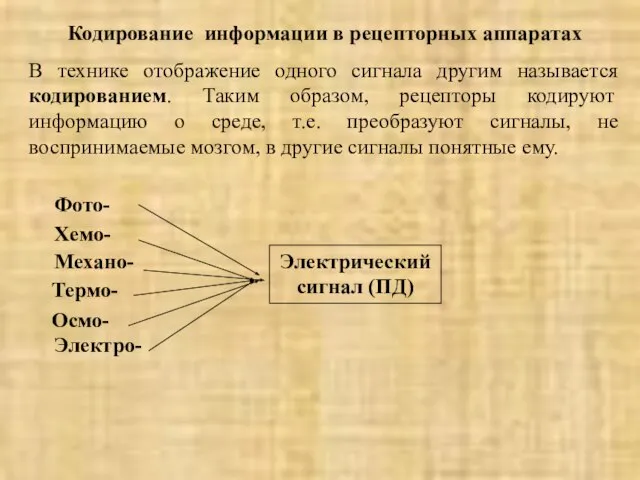
В технике отображение одного сигнала другим называется кодированием.
Кодирование информации в рецепторныx аппаратах
В технике отображение одного сигнала другим называется кодированием.
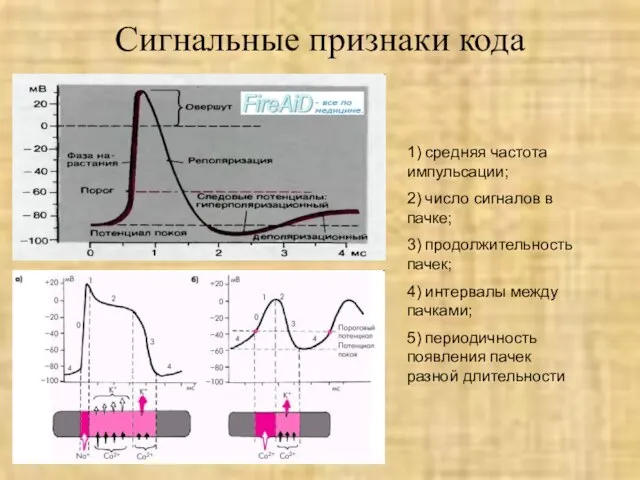
Слайд 18Сигнальные признаки кода
1) средняя частота импульсации;
2) число сигналов в пачке;
3) продолжительность пачек;
4)
Сигнальные признаки кода
1) средняя частота импульсации;
2) число сигналов в пачке;
3) продолжительность пачек;
4)
Слайд 19Кодирование силы раздражителя
Роль рефрактерности в преобразовании аналогового сигнала в дискретный и кодировании
Кодирование силы раздражителя
Роль рефрактерности в преобразовании аналогового сигнала в дискретный и кодировании
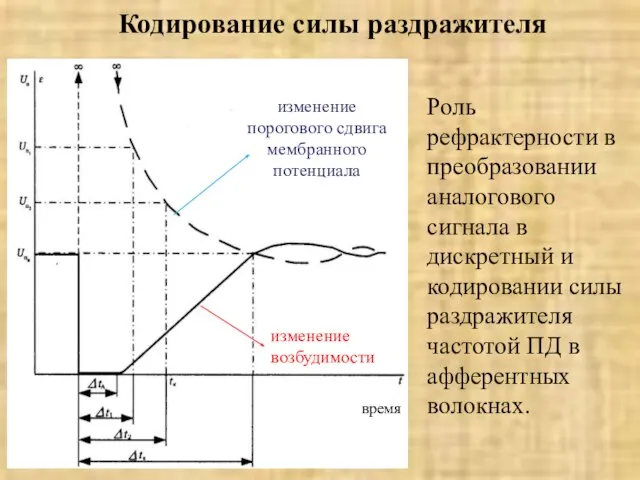
Слайд 20Э. Вебер сформулировал его так: ощущение растет в арифметической прогрессии при нарастании
Э. Вебер сформулировал его так: ощущение растет в арифметической прогрессии при нарастании

Слайд 21Соотношение между интенсивностью раздражителя (I) и частотой импульсов в афферентном волокне (ν
Соотношение между интенсивностью раздражителя (I) и частотой импульсов в афферентном волокне (ν

Слайд 22Графическое изображение законов Вебера-Фехнера (сверху) и Стивенса
Графическое изображение законов Вебера-Фехнера (сверху) и Стивенса
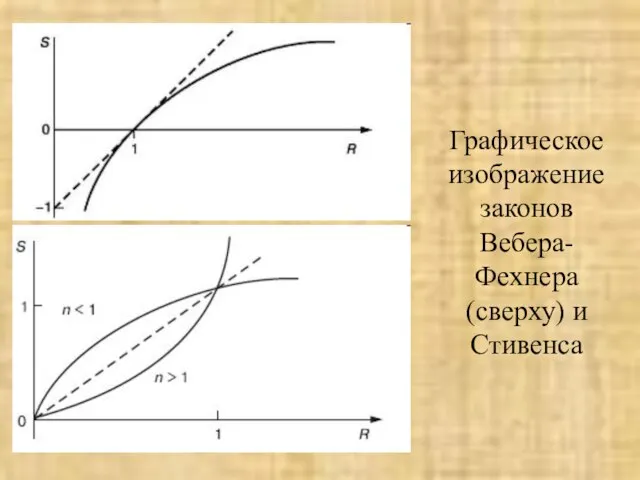
Слайд 23Кодирование качества раздражителя.
теория «меченой линии»
Основана на принципе анатомической локализации.
Содержание гипотезы: каждый рецептор
Кодирование качества раздражителя.
теория «меченой линии»
Основана на принципе анатомической локализации.
Содержание гипотезы: каждый рецептор

Слайд 24Биофизика слуха
Биофизика слуха

Слайд 25Схема органа слуха: А- ушная раковина; В - наружный слуховой проход; С
Схема органа слуха: А- ушная раковина; В - наружный слуховой проход; С
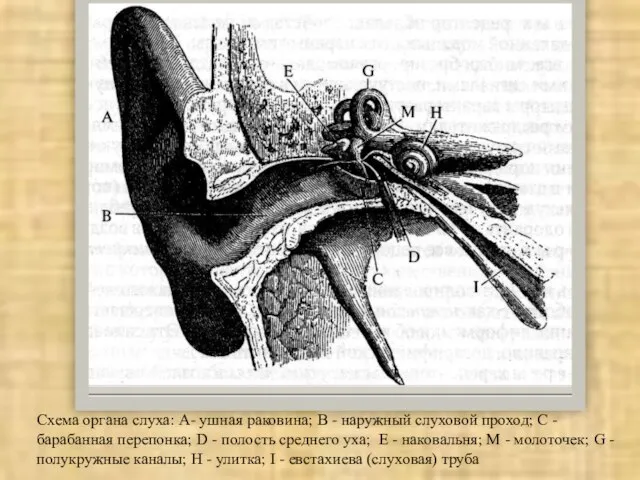
Слайд 26Костная улитка: а - стержень улитки (сканирующая электронная микроскопия): б— улитка с
Костная улитка: а - стержень улитки (сканирующая электронная микроскопия): б— улитка с
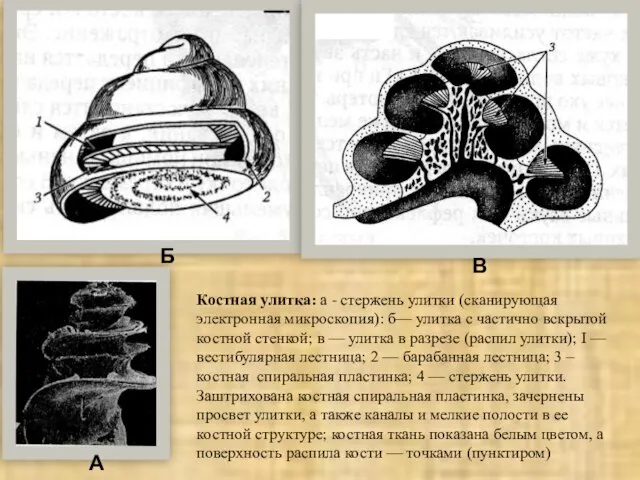
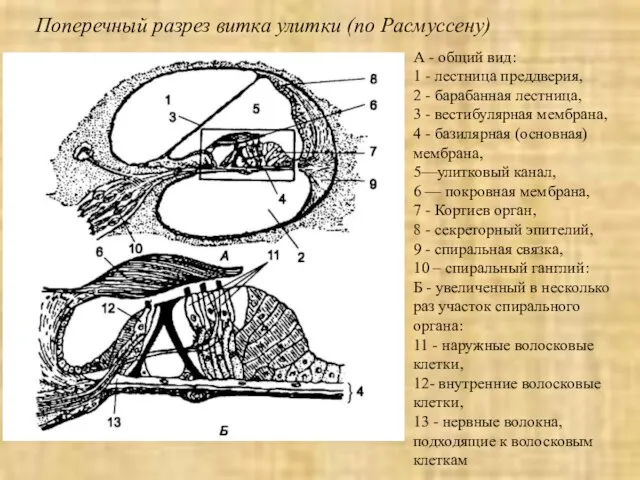
Слайд 27А - общий вид:
1 - лестница преддверия,
2 - барабанная лестница,
3
А - общий вид:
1 - лестница преддверия,
2 - барабанная лестница,
3
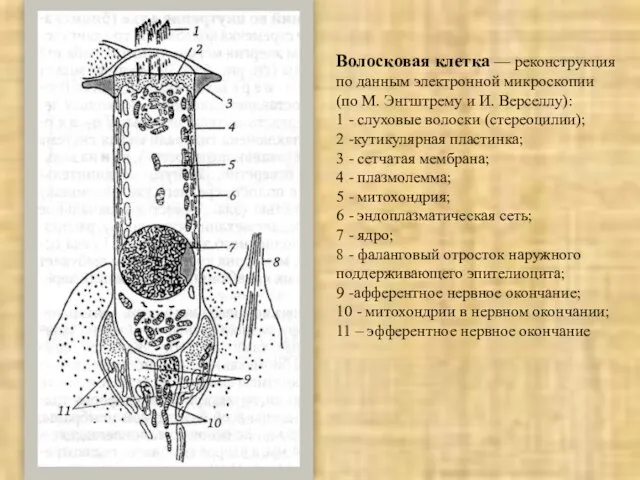
Слайд 28Волосковая клетка — реконструкция
по данным электронной микроскопии
(по М. Энгштрему и И. Верселлу):
1
Волосковая клетка — реконструкция
по данным электронной микроскопии
(по М. Энгштрему и И. Верселлу):
1
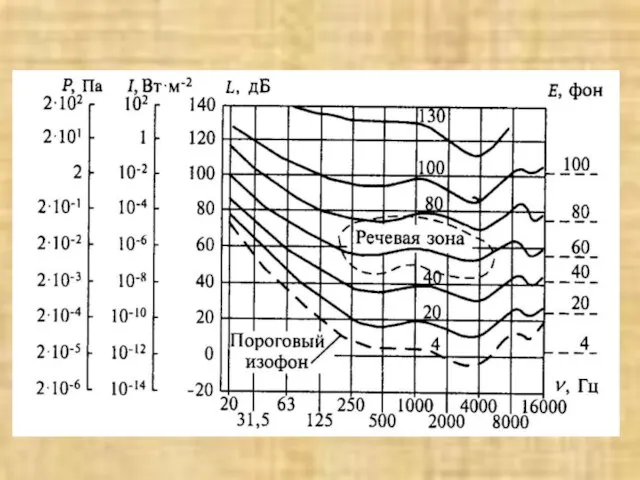
Слайд 29Зависимость местоположения максимального смешения базилярной мембраны человека от частоты звукового тона:
а -
Зависимость местоположения максимального смешения базилярной мембраны человека от частоты звукового тона:
а -

Слайд 31Сократительные системы
Сократительные системы

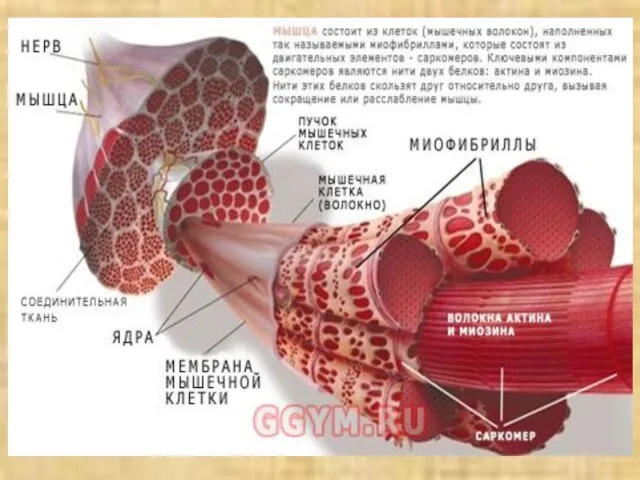
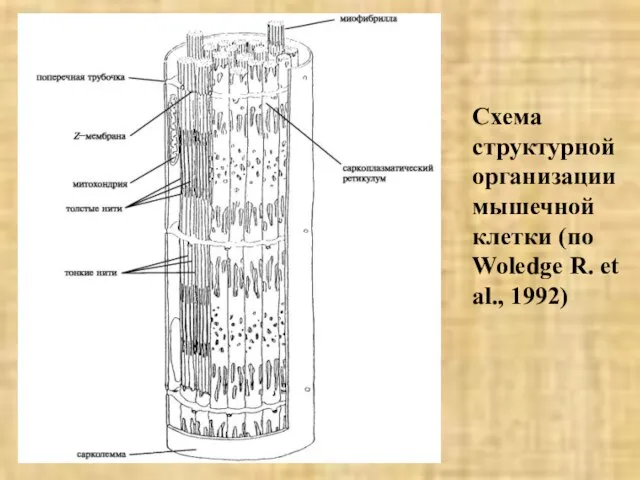
Слайд 33Схема структурной организации мышечной клетки (по Woledge R. et al., 1992)
Схема структурной организации мышечной клетки (по Woledge R. et al., 1992)

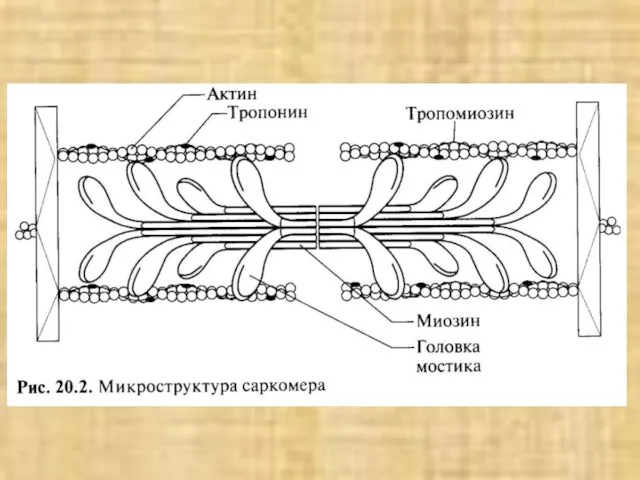
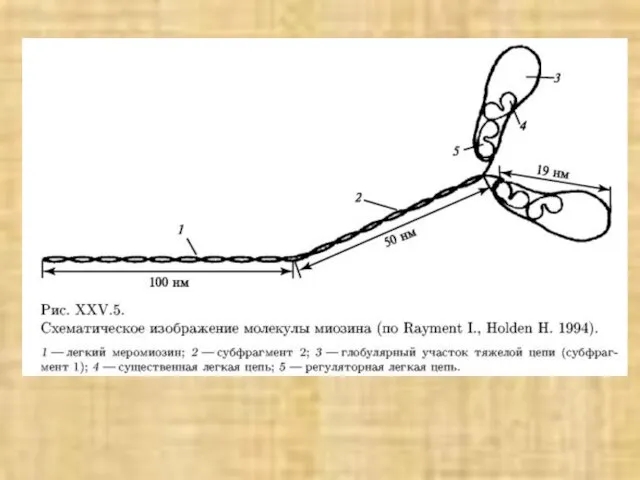
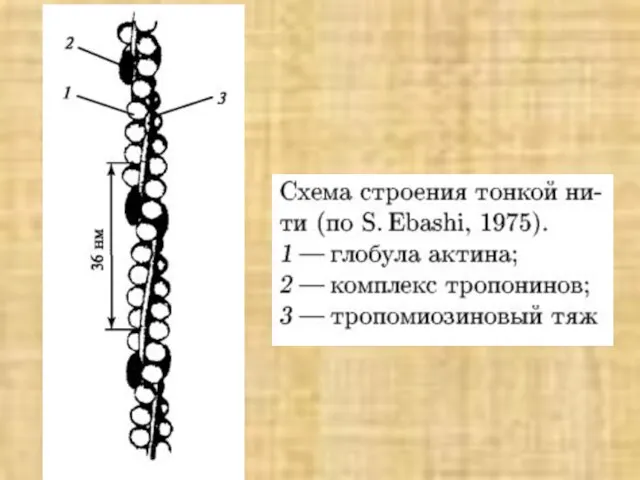
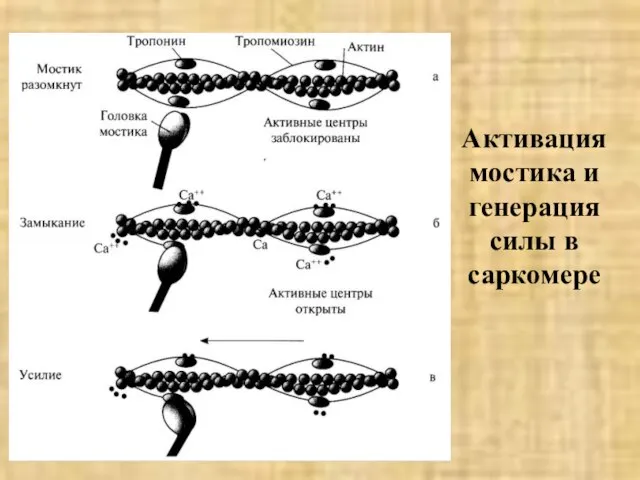
Слайд 38Активация мостика и генерация силы в саркомере
Активация мостика и генерация силы в саркомере
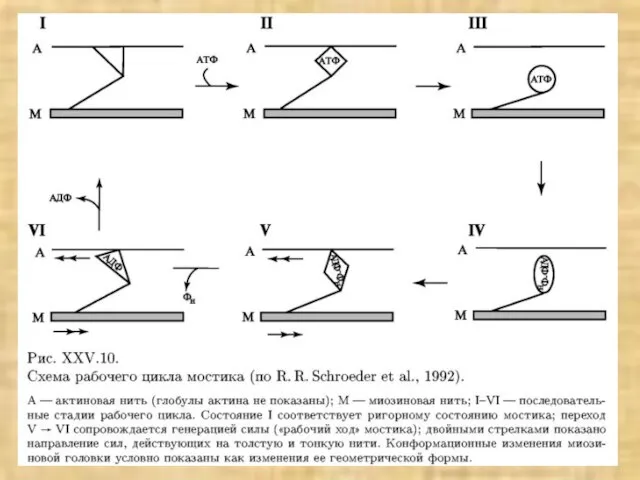
Слайд 40Основные положения модели скользящих нитей
1. Длины нитей актина и миозина в ходе
Основные положения модели скользящих нитей
1. Длины нитей актина и миозина в ходе

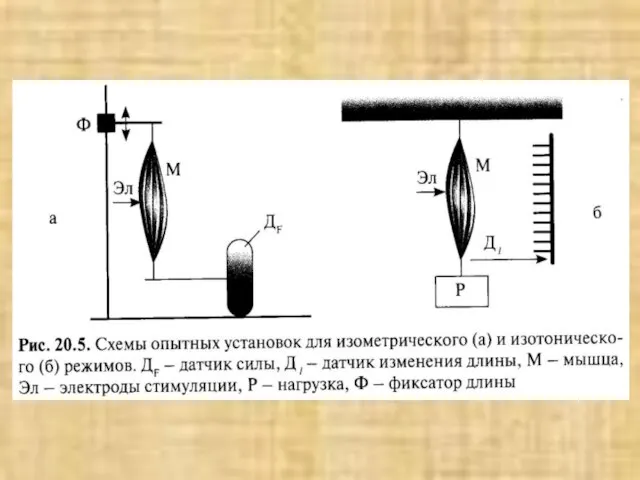
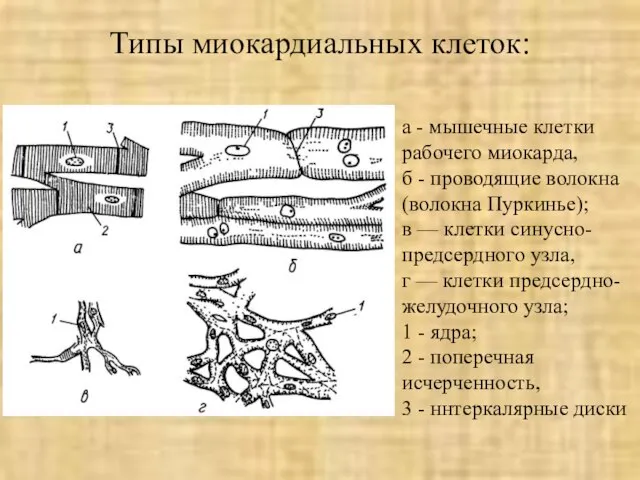
Слайд 42Типы миокардиальных клеток:
а - мышечные клетки рабочего миокарда,
б - проводящие волокна
Типы миокардиальных клеток:
а - мышечные клетки рабочего миокарда,
б - проводящие волокна
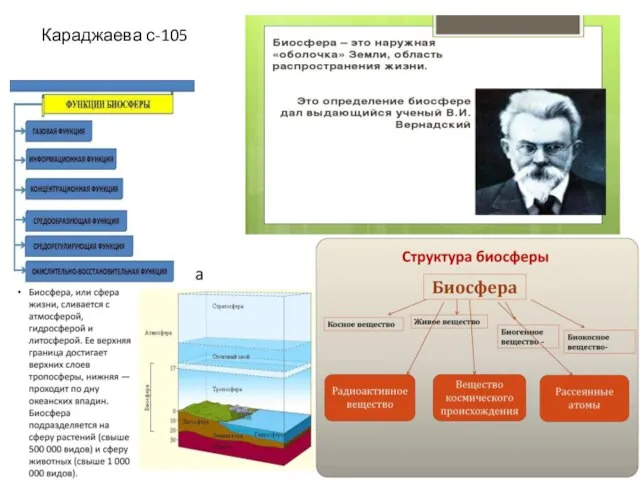 Структура биосферы
Структура биосферы Мир насекомых. Фоторабота Артамоновой Елизаветы
Мир насекомых. Фоторабота Артамоновой Елизаветы Анатомические и функциональные цепи
Анатомические и функциональные цепи Экстерьер передних конечностей
Экстерьер передних конечностей Берегите зрение
Берегите зрение Белки
Белки Движение крови в организме. Органы кровообращения
Движение крови в организме. Органы кровообращения Устойчивость лесов
Устойчивость лесов Основы физиологии равновесия
Основы физиологии равновесия Основы физиологии пищеварения. Тема №2
Основы физиологии пищеварения. Тема №2 Эпителиальная ткань
Эпителиальная ткань Строение зрительного анализатора
Строение зрительного анализатора Типы взаимодействия между организмами в сообществе
Типы взаимодействия между организмами в сообществе Экология паразитов
Экология паразитов Презентация на тему Биоценоз
Презентация на тему Биоценоз  Скаты. Общие характеристики
Скаты. Общие характеристики химия 14 пар
химия 14 пар Проблема филогенетических отношений билатерий
Проблема филогенетических отношений билатерий Презентация на тему ОДНОКЛЕТОЧНЫЕ ОРГАНИЗМЫ
Презентация на тему ОДНОКЛЕТОЧНЫЕ ОРГАНИЗМЫ  Лето. Луговые цветы
Лето. Луговые цветы Презентация на тему Глаз как оптический прибор и зрение
Презентация на тему Глаз как оптический прибор и зрение  Боль в спине. Истории пациентов (собак)
Боль в спине. Истории пациентов (собак) Для чего человеку зубы
Для чего человеку зубы Презентация на тему Веселая биология
Презентация на тему Веселая биология  Головной мозг. Нервная система. Рефлекс. Нервный импульс
Головной мозг. Нервная система. Рефлекс. Нервный импульс Комнатные растения
Комнатные растения Шток-роза
Шток-роза Рапс және қыша дақылдарының зиянкестері
Рапс және қыша дақылдарының зиянкестері