- Трансляция

Содержание
- 2. ТРАНСЛЯЦИЯ Перевод генетической информации мРНК, записанной с помощью четырех нуклеотидов, в первичную структуру белка (полипептид), записанную
- 4. Рибосомы Рибосомы - внутриклеточный компартмент, где происходит трансляция. Полирибосомы (полисомы) - несколько рибосом, транслирующих одну и
- 5. Строение рибосом Химически рибосомы представляют собой нуклеопротеины, состоящие из РНК и белков в соотношении 1:1 у
- 6. Строение рибосом Рибосома состоит из 2 субъединиц: малая (30S – прокариоты или 40S - эукариоты) и
- 8. РИБОСОМНЫЙ ЦИКЛ ДЖ.УОТСОНА В начале синтеза полипептидной цепи субъединицы рибосомы объединяются на 5’-конце мРНК в функционирующую
- 9. ПЕРЕДАЧА ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ ПРИ СИНТЕЗЕ БЕЛКА ДНК: информация о последовательности аминокислот в полипептидной цепи записана в
- 10. Генетический код
- 11. ГЕНЕТИЧЕСКИЙ КОД (мРНК) Инициирующие кодоны – АУГ и ГУГ (кодируют включение формилметионина у прокариот или метионина
- 12. ЭТАПЫ СИНТЕЗА БЕЛКА Активация аминокислот с образованием амино-ацил-тРНК (аа-тРНК); Инициация полипептидной цепи; Элонгация полипептидной цепи; Терминация
- 13. Белоксинтезирующая система клетки мРНК – матрица, на которой записана последовательность аминокислот белка в виде последовательности триплетов.
- 14. АКТИВАЦИЯ АМИНОКИСЛОТ В цитозоле 20 аминокислот соединяются с соответствующими тРНК с образованием аа-тРНК. Ферменты – аа-тРНК-синтетазы
- 15. Реакция активации аминокислот Аминокислота + АТФ + тРНК ? аа–тРНК + АМФ + ФФ. 2 этапа:
- 16. Образование аминоацил-тРНК
- 17. Стадия инициации Необходимо: субъединицы рибосом, инициирующие факторы IF-1, IF-2, IF-3, (формил)метионин-тРНК, мРНК, ГТФ. Антикодон (формил)метионин-тРНК УАЦ
- 19. Стадия инициации (продолжение) В рибосоме – два участка связывания аа-тРНК: А (аминоацильный) имеет основное сродство к
- 20. Стадия элонгации Необходимы: набор аа-тРНК, факторы элонгации EF-T и EF-g, ГТФ. Этапы: 1) присоединение аа-тРНК к
- 23. Стадия терминации Факторы терминации FR-1 (УАА и УАГ) и FR-2 (УАА и УГА). Терминирующие кодоны (в
- 25. Фолдинг и процессинг От синтезированного пептида в цитозоле отщепляется инициирующая аминокислота (формил)метионин. Сигнальная последовательность на N-конце
- 28. Регуляция синтеза белка Возможна на всех стадиях. Индукция синтеза белка – вещество понижает сродство репрессора к
- 29. Препараты-регуляторы синтеза белка I класс. ИНДУКТОРЫ (анаболики) 1) Гормональные а) специфические – стероидные гормоны. Глюкокортикоиды (индукция
- 30. Ингибиторы транскрипции Рифампицин связывается с β-субъединицей РНК-полимеразы, ингибируя образование первой фосфодиэфирной связи в транскрипте. На уже
- 31. Ингибиторы трансляции Стрептомицин – препятствует связыванию с рибосомой формилметионин-тРНК, нарушая инициацию трансляции. Связывается с белком малой
- 32. Действие токсинов Аманитин (токсин бледной поганки), циклический пептид, связывается с эукариотической РНК-полимеразой II, блокируя синтез мРНК.
- 33. Ингибиторы репликации ДНК Антибиотики способны: 1) встраиваться (интеркаляция) между основаниями ДНК, ингибируя ее матричную активность (дауномицин,
- 37. Скачать презентацию
Слайд 2ТРАНСЛЯЦИЯ
Перевод генетической информации мРНК, записанной с помощью четырех нуклеотидов, в первичную структуру
ТРАНСЛЯЦИЯ
Перевод генетической информации мРНК, записанной с помощью четырех нуклеотидов, в первичную структуру

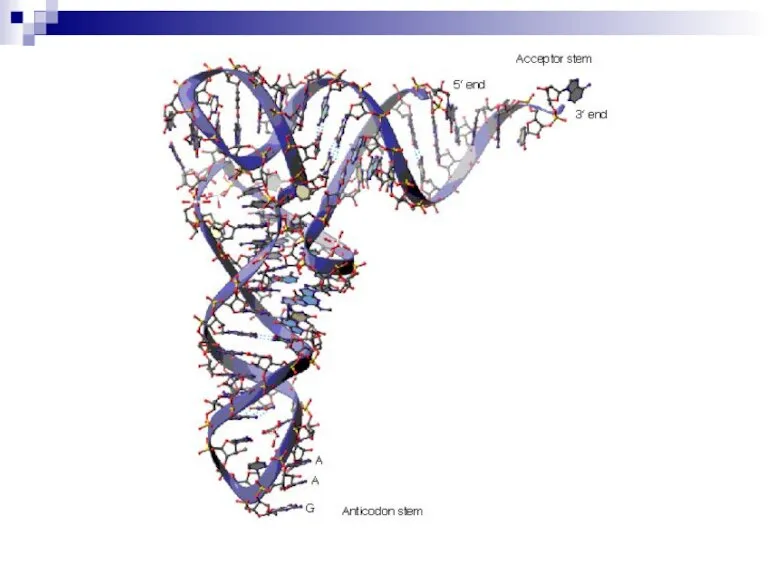
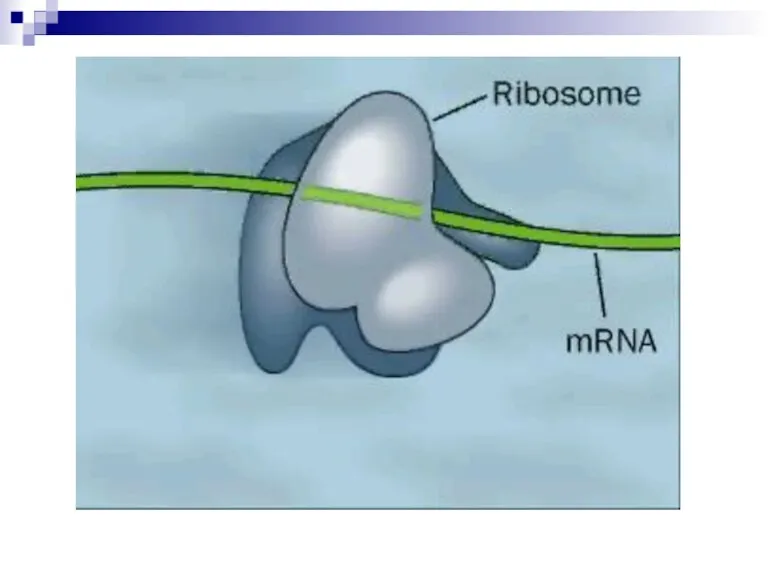
Слайд 4Рибосомы
Рибосомы - внутриклеточный компартмент, где происходит трансляция.
Полирибосомы (полисомы) - несколько рибосом, транслирующих
Рибосомы
Рибосомы - внутриклеточный компартмент, где происходит трансляция.
Полирибосомы (полисомы) - несколько рибосом, транслирующих

Слайд 5Строение рибосом
Химически рибосомы представляют собой нуклеопротеины, состоящие из РНК и белков в
Строение рибосом
Химически рибосомы представляют собой нуклеопротеины, состоящие из РНК и белков в

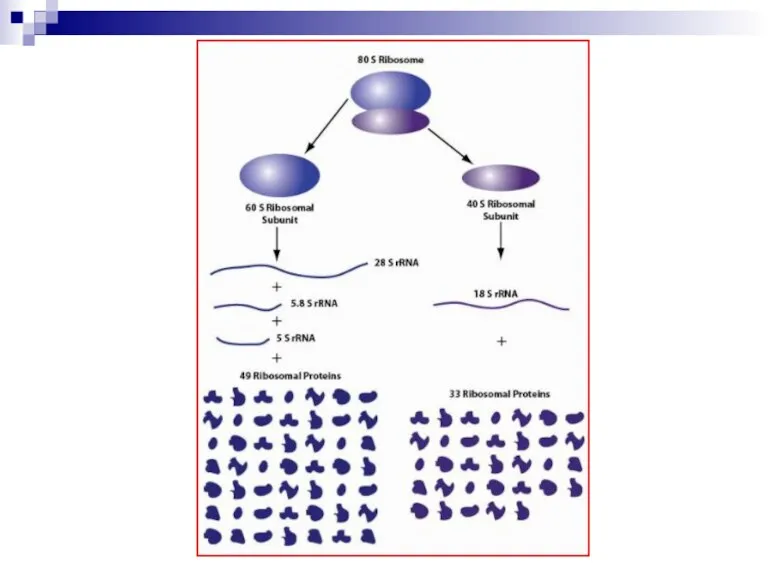
Слайд 6Строение рибосом
Рибосома состоит из 2 субъединиц: малая (30S – прокариоты или 40S
Строение рибосом
Рибосома состоит из 2 субъединиц: малая (30S – прокариоты или 40S

Слайд 8РИБОСОМНЫЙ ЦИКЛ ДЖ.УОТСОНА
В начале синтеза полипептидной цепи субъединицы рибосомы объединяются на 5’-конце
РИБОСОМНЫЙ ЦИКЛ ДЖ.УОТСОНА
В начале синтеза полипептидной цепи субъединицы рибосомы объединяются на 5’-конце

Слайд 9ПЕРЕДАЧА ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ ПРИ СИНТЕЗЕ БЕЛКА
ДНК: информация о последовательности аминокислот в полипептидной
ПЕРЕДАЧА ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ ПРИ СИНТЕЗЕ БЕЛКА
ДНК: информация о последовательности аминокислот в полипептидной

Слайд 10Генетический код
Генетический код
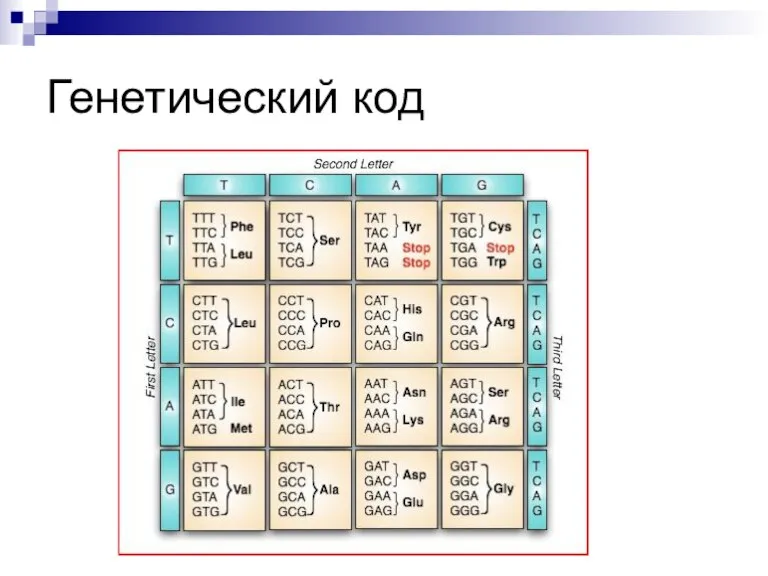
Слайд 11ГЕНЕТИЧЕСКИЙ КОД (мРНК)
Инициирующие кодоны – АУГ и ГУГ (кодируют включение формилметионина у
ГЕНЕТИЧЕСКИЙ КОД (мРНК)
Инициирующие кодоны – АУГ и ГУГ (кодируют включение формилметионина у

Слайд 12ЭТАПЫ СИНТЕЗА БЕЛКА
Активация аминокислот с образованием амино-ацил-тРНК (аа-тРНК);
Инициация полипептидной цепи;
Элонгация полипептидной
ЭТАПЫ СИНТЕЗА БЕЛКА
Активация аминокислот с образованием амино-ацил-тРНК (аа-тРНК);
Инициация полипептидной цепи;
Элонгация полипептидной

Слайд 13Белоксинтезирующая система клетки
мРНК – матрица, на которой записана последовательность аминокислот белка в
Белоксинтезирующая система клетки
мРНК – матрица, на которой записана последовательность аминокислот белка в

Слайд 14АКТИВАЦИЯ АМИНОКИСЛОТ
В цитозоле 20 аминокислот соединяются с соответствующими тРНК с образованием аа-тРНК.
Ферменты
АКТИВАЦИЯ АМИНОКИСЛОТ
В цитозоле 20 аминокислот соединяются с соответствующими тРНК с образованием аа-тРНК.
Ферменты

Слайд 15Реакция активации аминокислот
Аминокислота + АТФ + тРНК ?
аа–тРНК + АМФ +
Реакция активации аминокислот
Аминокислота + АТФ + тРНК ? аа–тРНК + АМФ +

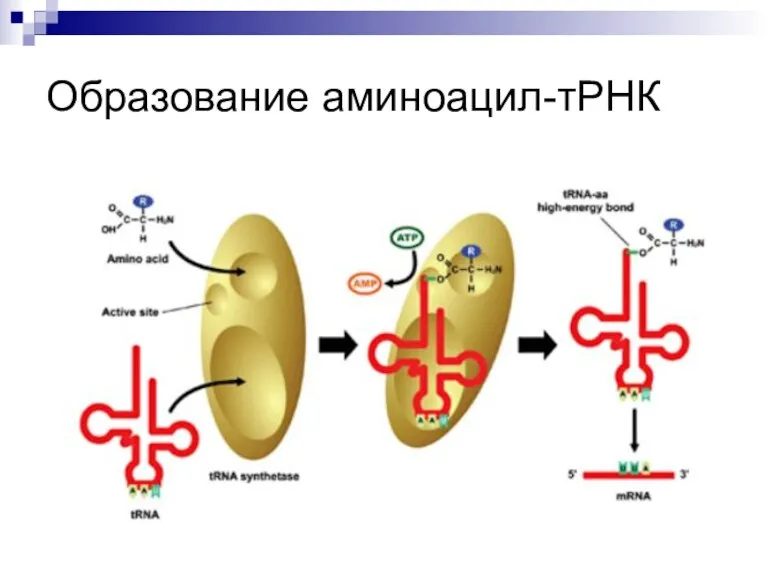
Слайд 16Образование аминоацил-тРНК
Образование аминоацил-тРНК
Слайд 17Стадия инициации
Необходимо: субъединицы рибосом, инициирующие факторы IF-1, IF-2, IF-3, (формил)метионин-тРНК, мРНК, ГТФ.
Антикодон
Стадия инициации
Необходимо: субъединицы рибосом, инициирующие факторы IF-1, IF-2, IF-3, (формил)метионин-тРНК, мРНК, ГТФ.
Антикодон

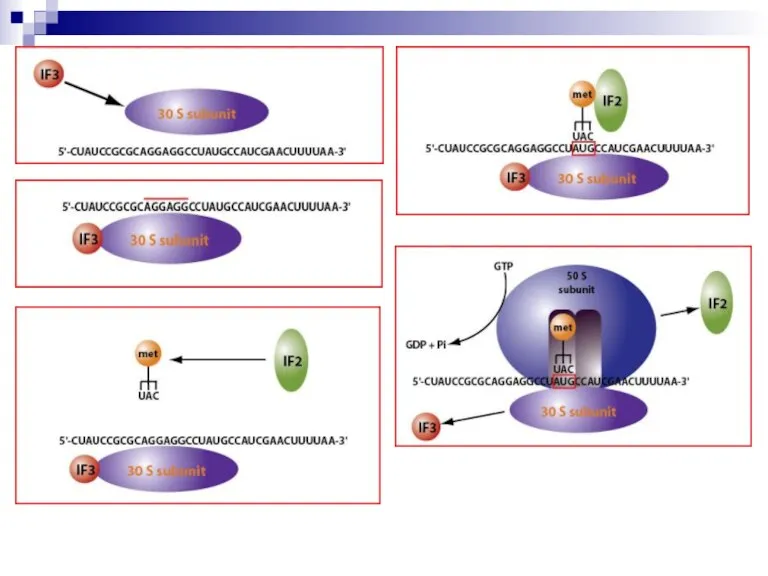
Слайд 19Стадия инициации (продолжение)
В рибосоме – два участка связывания аа-тРНК: А (аминоацильный) имеет
Стадия инициации (продолжение)
В рибосоме – два участка связывания аа-тРНК: А (аминоацильный) имеет

Слайд 20Стадия элонгации
Необходимы: набор аа-тРНК, факторы элонгации EF-T и EF-g, ГТФ.
Этапы: 1) присоединение
Стадия элонгации
Необходимы: набор аа-тРНК, факторы элонгации EF-T и EF-g, ГТФ.
Этапы: 1) присоединение

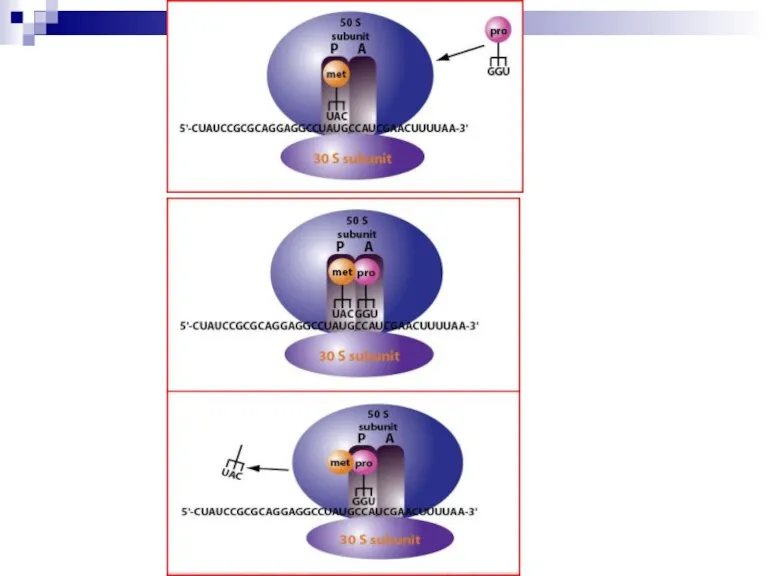
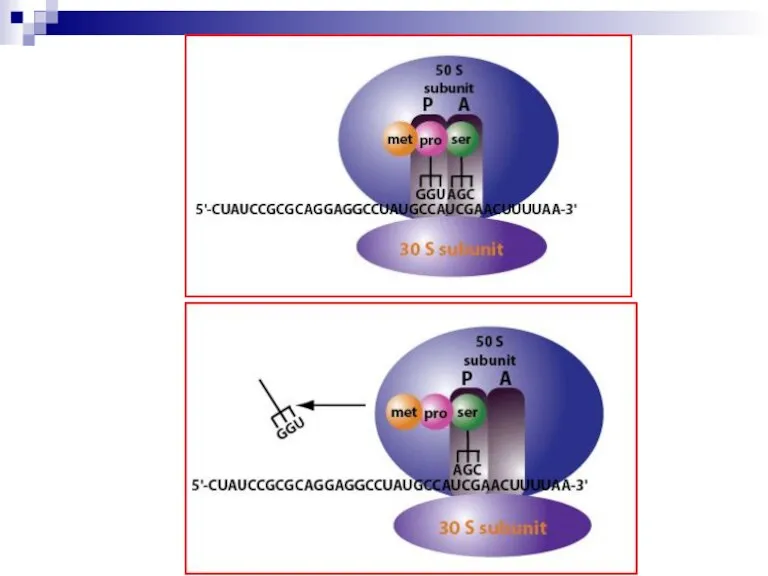
Слайд 23Стадия терминации
Факторы терминации FR-1 (УАА и УАГ) и FR-2 (УАА и УГА).
Стадия терминации
Факторы терминации FR-1 (УАА и УАГ) и FR-2 (УАА и УГА).

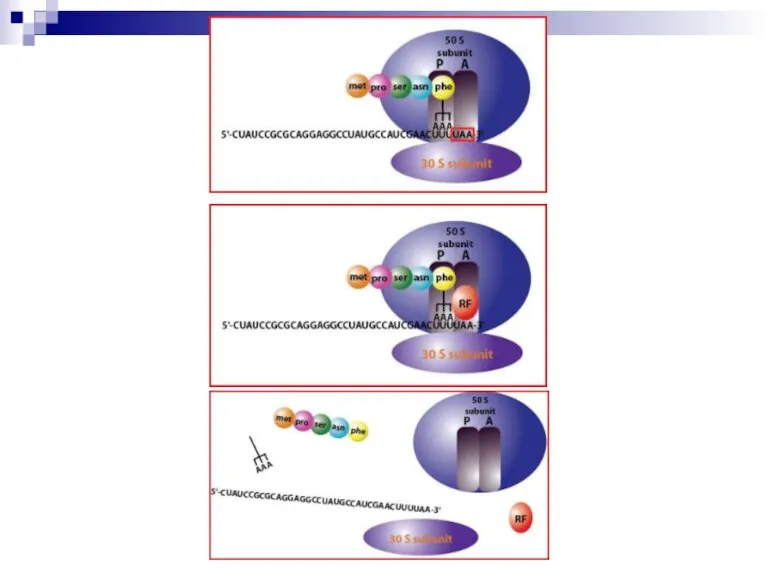
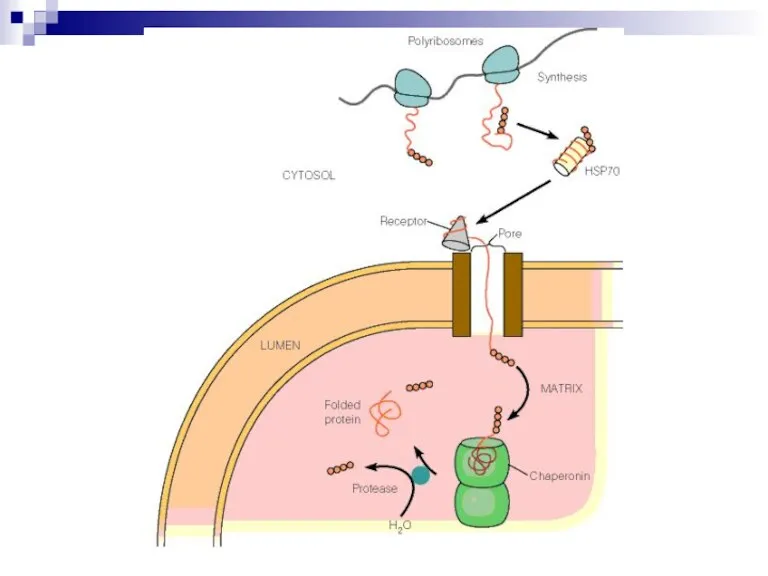
Слайд 25Фолдинг и процессинг
От синтезированного пептида в цитозоле отщепляется инициирующая аминокислота (формил)метионин.
Сигнальная последовательность
Фолдинг и процессинг
От синтезированного пептида в цитозоле отщепляется инициирующая аминокислота (формил)метионин.
Сигнальная последовательность

Слайд 28Регуляция синтеза белка
Возможна на всех стадиях.
Индукция синтеза белка – вещество понижает сродство
Регуляция синтеза белка
Возможна на всех стадиях.
Индукция синтеза белка – вещество понижает сродство

Слайд 29Препараты-регуляторы синтеза белка
I класс. ИНДУКТОРЫ (анаболики)
1) Гормональные
а) специфические – стероидные
Препараты-регуляторы синтеза белка
I класс. ИНДУКТОРЫ (анаболики)
1) Гормональные
а) специфические – стероидные

Слайд 30Ингибиторы транскрипции
Рифампицин связывается с
β-субъединицей РНК-полимеразы, ингибируя образование первой фосфодиэфирной связи в
Ингибиторы транскрипции
Рифампицин связывается с β-субъединицей РНК-полимеразы, ингибируя образование первой фосфодиэфирной связи в

Слайд 31Ингибиторы трансляции
Стрептомицин – препятствует связыванию с рибосомой формилметионин-тРНК, нарушая инициацию трансляции. Связывается
Ингибиторы трансляции
Стрептомицин – препятствует связыванию с рибосомой формилметионин-тРНК, нарушая инициацию трансляции. Связывается

Слайд 32Действие токсинов
Аманитин (токсин бледной поганки), циклический пептид, связывается с эукариотической РНК-полимеразой II,
Действие токсинов
Аманитин (токсин бледной поганки), циклический пептид, связывается с эукариотической РНК-полимеразой II,

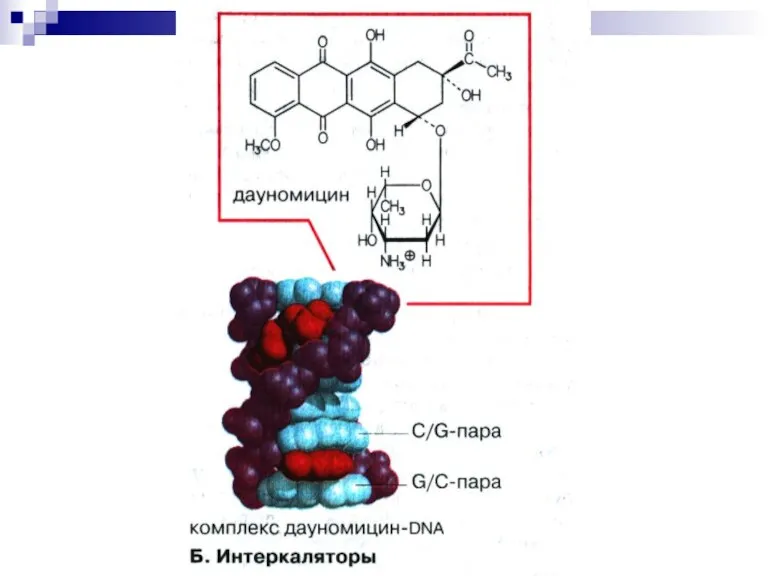
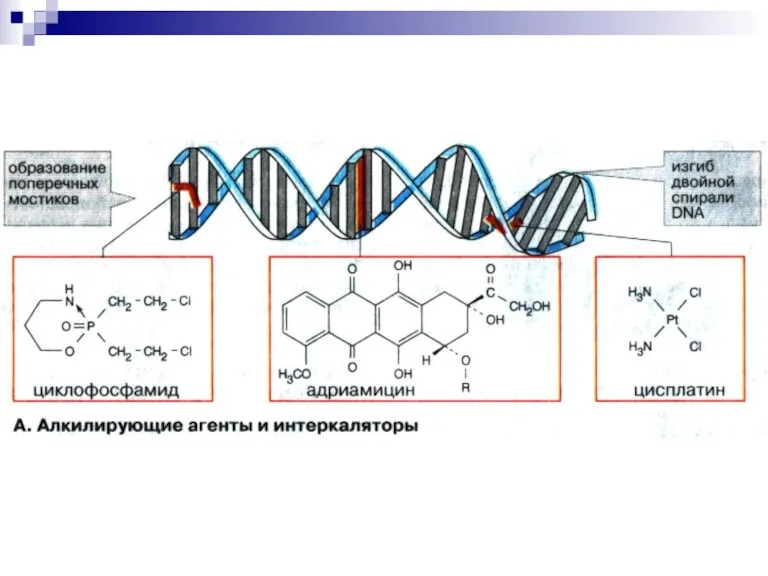
Слайд 33Ингибиторы репликации ДНК
Антибиотики способны:
1) встраиваться (интеркаляция) между основаниями ДНК,
Ингибиторы репликации ДНК
Антибиотики способны:
1) встраиваться (интеркаляция) между основаниями ДНК,

 Полигенное наследование
Полигенное наследование Эпифиз – төрт төмпешіктің алдыңғы төмпешіктернінің арасында орналасқан
Эпифиз – төрт төмпешіктің алдыңғы төмпешіктернінің арасында орналасқан Анатомия насекомых
Анатомия насекомых Растения у нашего дома
Растения у нашего дома Классификация условных и безусловных рефлексов по теории И.П. Павлова
Классификация условных и безусловных рефлексов по теории И.П. Павлова Структурно-функциональная дифференцировка прокариот
Структурно-функциональная дифференцировка прокариот Приспособления организмов к жизни в природе
Приспособления организмов к жизни в природе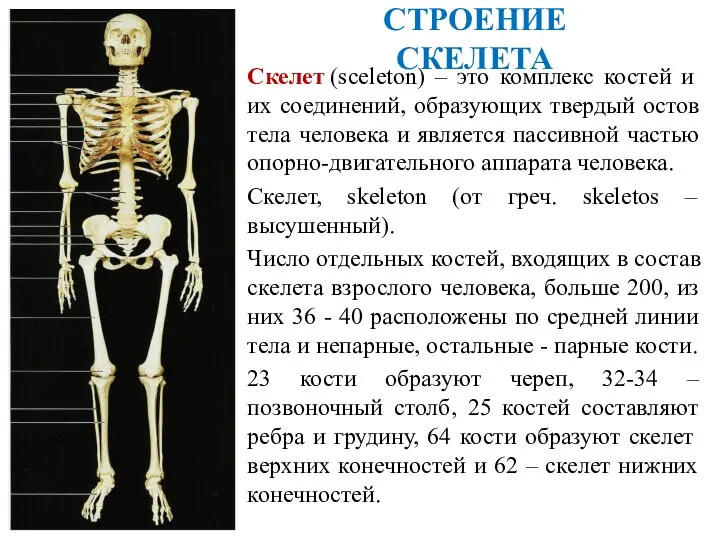 Презентация кости
Презентация кости Вуглеводи, ліпіди, нуклеїнові кислоти. Обмін речовин. Тест
Вуглеводи, ліпіди, нуклеїнові кислоти. Обмін речовин. Тест Эко - невидаль
Эко - невидаль Общие свойства живых организмов
Общие свойства живых организмов Возраст дерева
Возраст дерева Водоросли. Виды водорослей
Водоросли. Виды водорослей Жук олень
Жук олень Презентация на тему РАЗМНОЖЕНИЕ И ОПЛОДОТВОРЕНИЕ У РАСТЕНИЙ
Презентация на тему РАЗМНОЖЕНИЕ И ОПЛОДОТВОРЕНИЕ У РАСТЕНИЙ  Общая характеристика обмена веществ и преобразование энергии
Общая характеристика обмена веществ и преобразование энергии Внимание. Категории внимания
Внимание. Категории внимания Фотосинтез. Значение фотосинтеза
Фотосинтез. Значение фотосинтеза Молекулярная биология
Молекулярная биология Сохраним мир птиц
Сохраним мир птиц Значение насекомых в природе
Значение насекомых в природе Качкодзьоб - таємнича тварина
Качкодзьоб - таємнича тварина Паразитизм среди хордовых животных
Паразитизм среди хордовых животных Бактерии и вирусы
Бактерии и вирусы Строение корня
Строение корня Усики, лапки, крылышки
Усики, лапки, крылышки Презентация на тему Подтипы Бесчерепные и Черепные
Презентация на тему Подтипы Бесчерепные и Черепные  Работы И.В.Мичурина
Работы И.В.Мичурина