- Возникновение адаптивного иммунитета

Содержание
- 2. Эволюция иммунной системы на фоне филогенетического дерева животных
- 3. Систематика и эволюция оболочников
- 4. Систематика и эволюция рыб
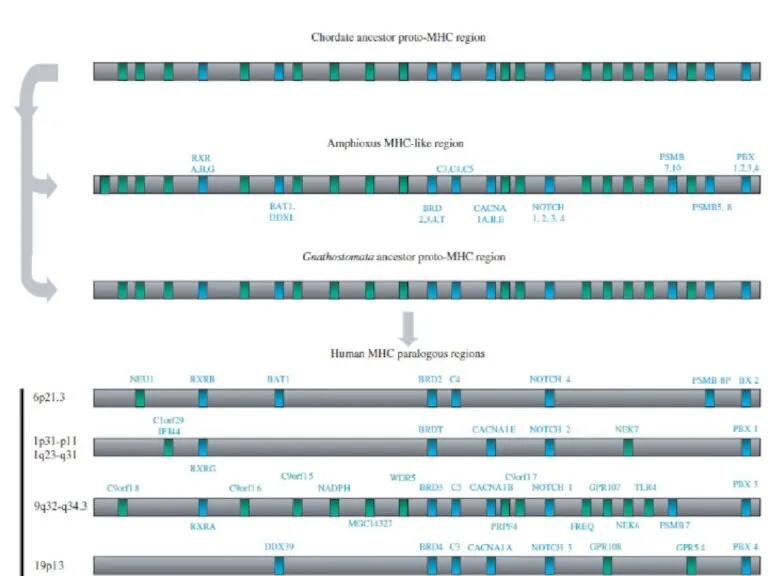
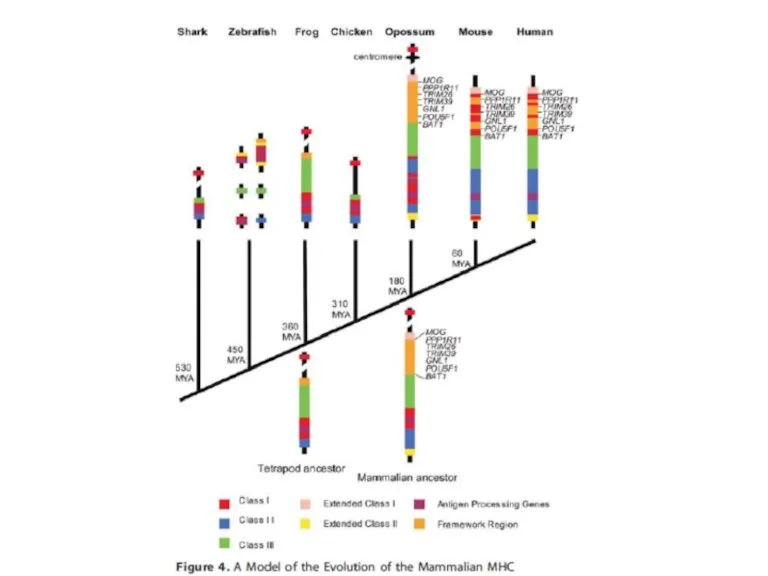
- 5. Две полногеномные дупликации как источник разнообразия паралогов
- 6. Синергичность компонентов адаптивного иммунитета создаёт проблемы с пониманием, как он мог возникнуть Что возникло раньше, антитела
- 7. Гипотеза Т. Boehm
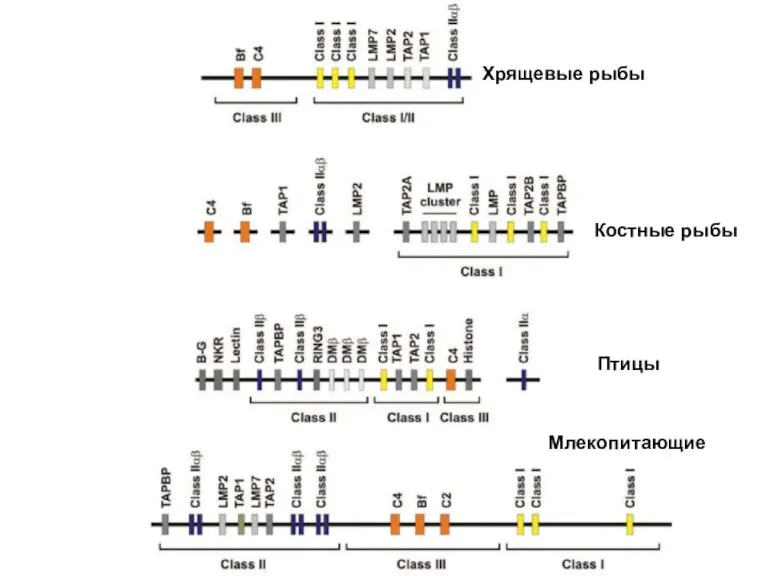
- 9. Хрящевые рыбы Костные рыбы Птицы Млекопитающие
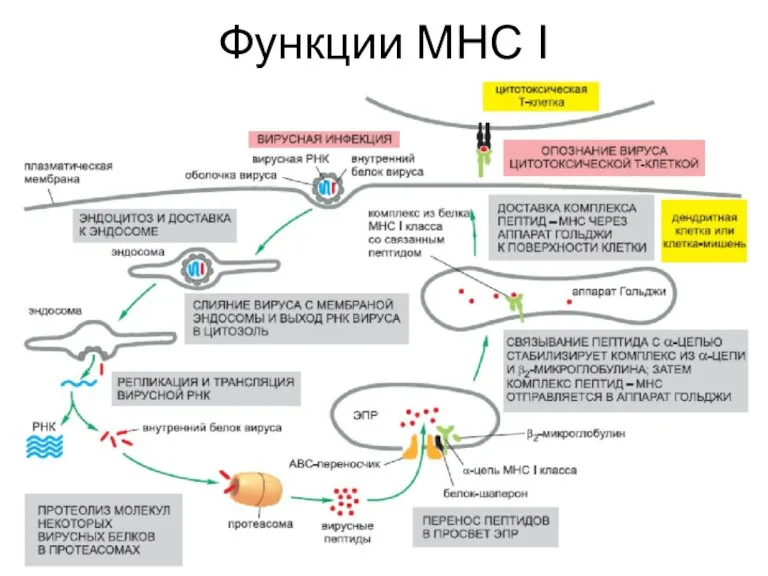
- 11. Функции МНС I
- 12. Функции MHC II
- 13. Возникновение МНС I
- 14. Треска не имеет функционального набора генов молекул MHCII, и рецептора CD4
- 15. Это является результатом цепочки хромосомных перестроек, затрагивающих локус МНС
- 16. Рыбы, экономящие на Т-хелперной ветви
- 17. Химера Callorhinchus milii утратила CD4…
- 18. … и несколько цитокинов и транскрипционных факторов (Venkatesh et al, Nature, 2014) CD8 T cell CD4
- 19. Роль рекомбиназ RAG David Schatz, клонировал предска-занные Сакано и Тонегавой рекомбиназы
- 20. Возникновение RAG генов После того, как группа Джона Маркелониса в 1994 году клонировала ген Rag1 акулы,
- 21. Однако гомологи RAG-генов нашлись после полного секвенирования генома морского ежа
- 22. Выяснилось, что гомологи RAG генов есть даже в геноме морского ежа Strongylocentrotus, однако отсутствуют во вчетверо
- 23. RAG1 сходен с транспозазой Transib из Kapitonov, Jurka 2005
- 24. Использование транспозона «в мирных целях»
- 28. Механизм переключения классов антител
- 29. Рецепторы врожденного иммунитета могли возникнуть из рецепторов NK-клеток
- 31. Rag-зависимая рекомбинация или конверсия генов?
- 32. Общая схема конверсии
- 33. Организация генов в кластеры или транслоконы или
- 34. Организации генов в кластеры более примитивна
- 35. Для переключения классов снова важна цитидин-дезаминаза AID
- 36. Эволюция семейств цитидиндезаминаз
- 38. Скачать презентацию
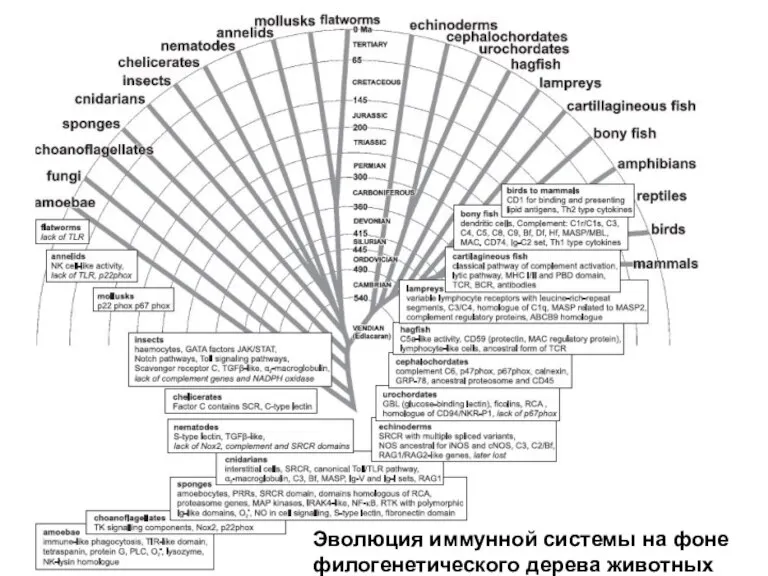
Слайд 3Систематика и эволюция оболочников
Систематика и эволюция оболочников
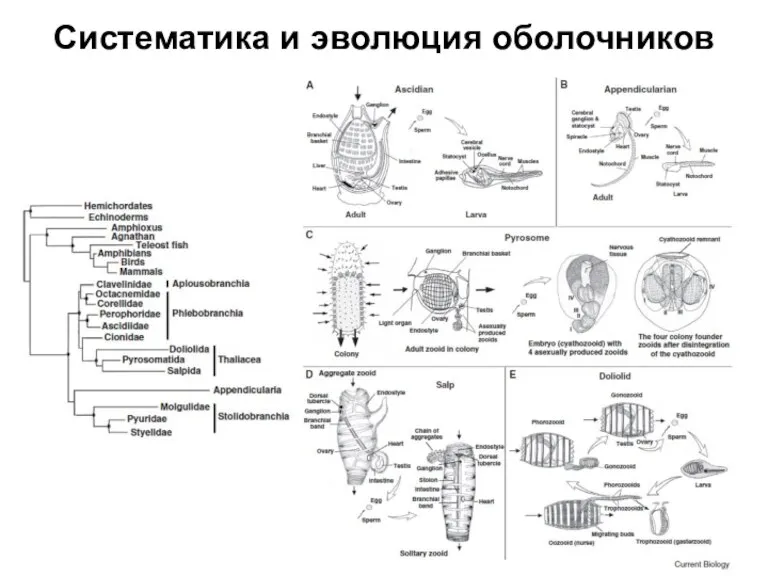
Слайд 4Систематика и эволюция рыб
Систематика и эволюция рыб
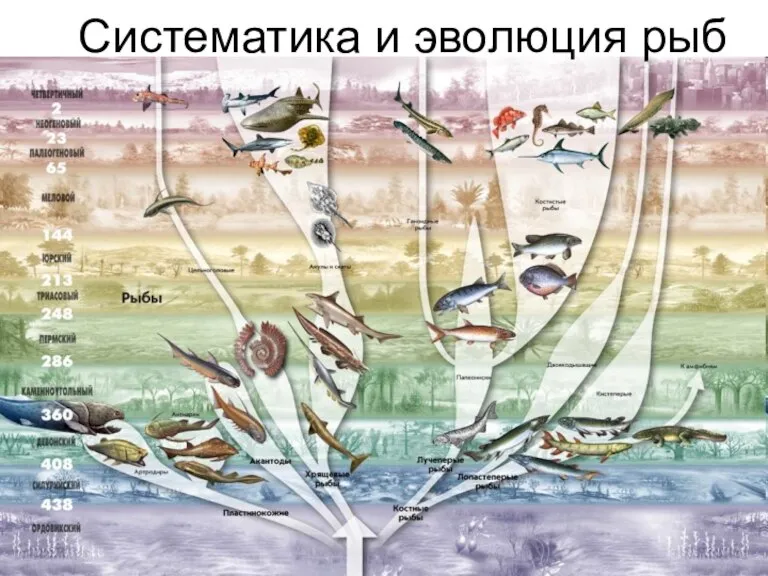
Слайд 5Две полногеномные дупликации как источник разнообразия паралогов
Две полногеномные дупликации как источник разнообразия паралогов
Слайд 6Синергичность компонентов адаптивного иммунитета создаёт проблемы с пониманием, как он мог возникнуть
Синергичность компонентов адаптивного иммунитета создаёт проблемы с пониманием, как он мог возникнуть

Слайд 7Гипотеза Т. Boehm
Гипотеза Т. Boehm

Слайд 9Хрящевые рыбы
Костные рыбы
Птицы
Млекопитающие
Хрящевые рыбы
Костные рыбы
Птицы
Млекопитающие
Слайд 11Функции МНС I
Функции МНС I
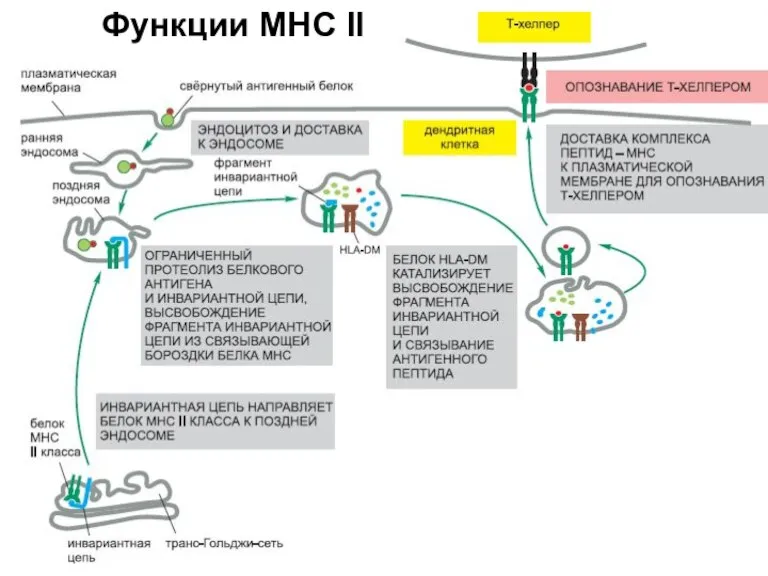
Слайд 12Функции MHC II
Функции MHC II
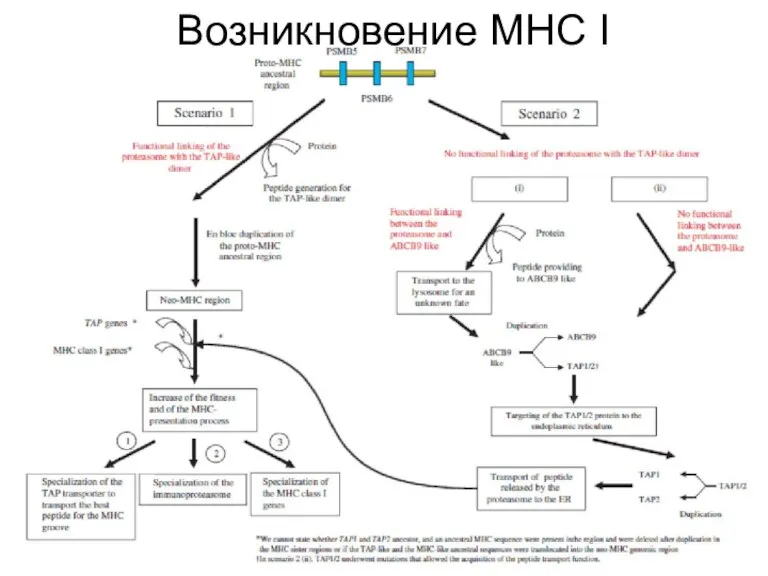
Слайд 13Возникновение МНС I
Возникновение МНС I
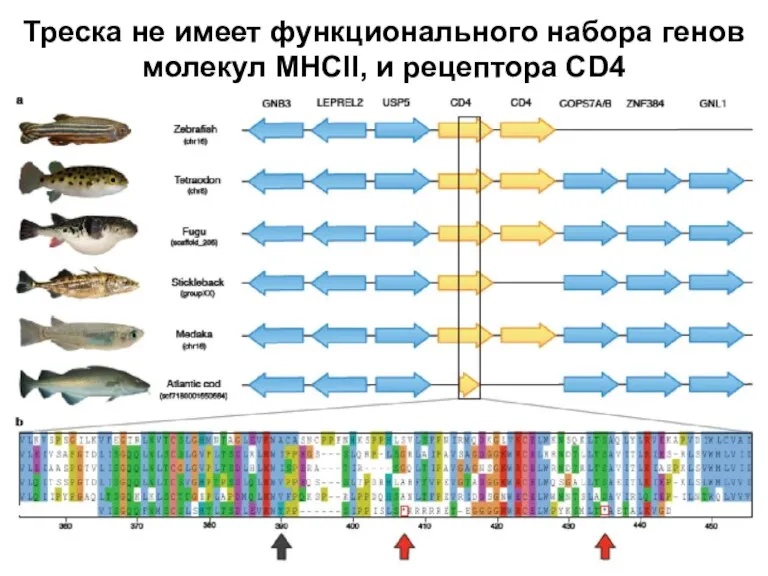
Слайд 14Треска не имеет функционального набора генов молекул MHCII, и рецептора CD4
Треска не имеет функционального набора генов молекул MHCII, и рецептора CD4
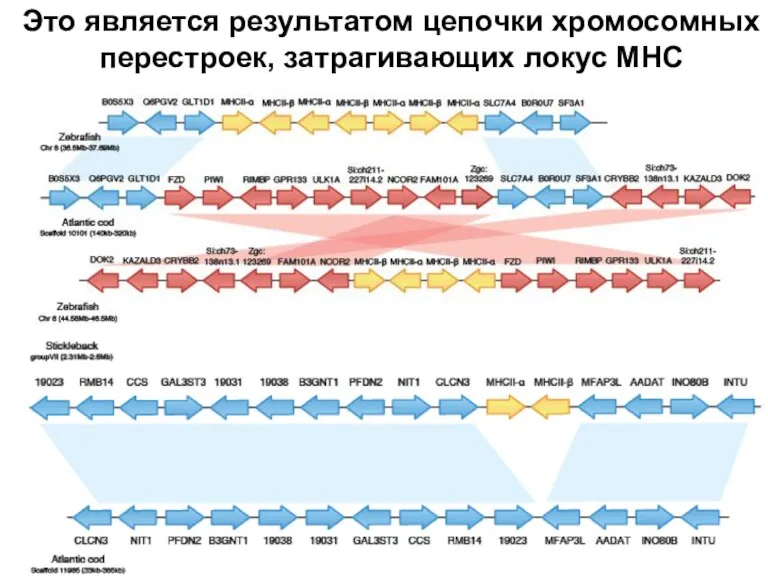
Слайд 15Это является результатом цепочки хромосомных перестроек, затрагивающих локус МНС
Это является результатом цепочки хромосомных перестроек, затрагивающих локус МНС
Слайд 16Рыбы, экономящие на Т-хелперной ветви
Рыбы, экономящие на Т-хелперной ветви

Слайд 17Химера Callorhinchus milii утратила CD4…
Химера Callorhinchus milii утратила CD4…

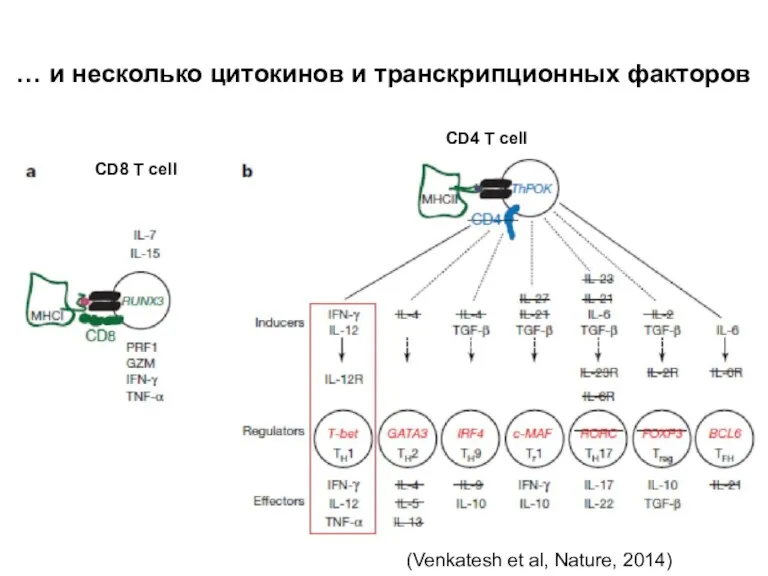
Слайд 18… и несколько цитокинов и транскрипционных факторов
(Venkatesh et al, Nature, 2014)
CD8 T
… и несколько цитокинов и транскрипционных факторов
(Venkatesh et al, Nature, 2014)
CD8 T
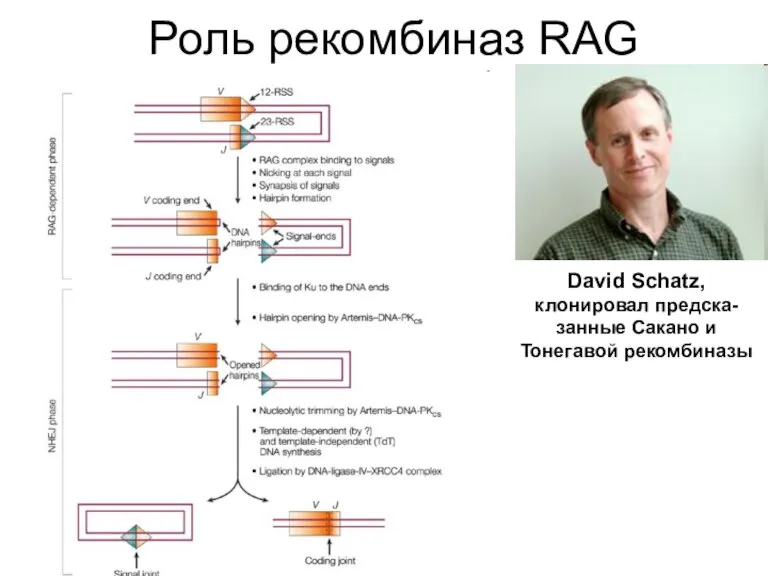
Слайд 19Роль рекомбиназ RAG
David Schatz, клонировал предска-занные Сакано и Тонегавой рекомбиназы
Роль рекомбиназ RAG
David Schatz, клонировал предска-занные Сакано и Тонегавой рекомбиназы
Слайд 20Возникновение RAG генов
После того, как группа Джона Маркелониса в 1994 году клонировала
Возникновение RAG генов
После того, как группа Джона Маркелониса в 1994 году клонировала

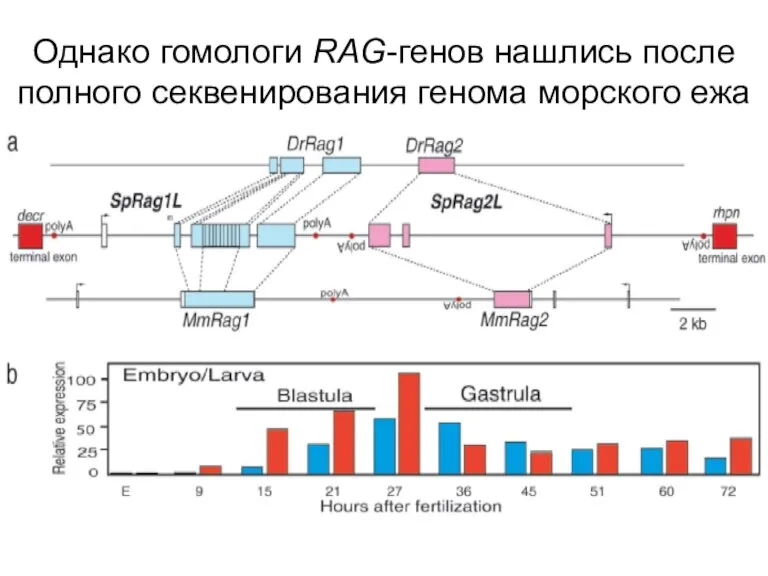
Слайд 21Однако гомологи RAG-генов нашлись после полного секвенирования генома морского ежа
Однако гомологи RAG-генов нашлись после полного секвенирования генома морского ежа
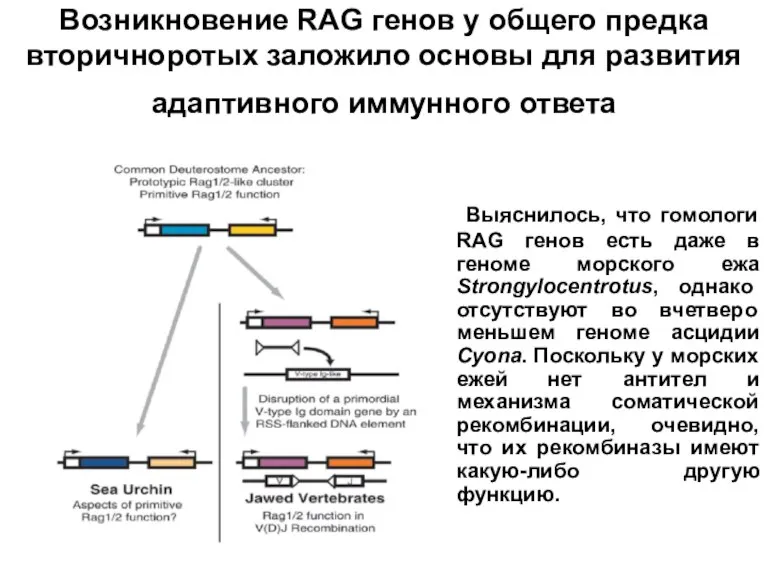
Слайд 22 Выяснилось, что гомологи RAG генов есть даже в геноме морского ежа Strongylocentrotus,
Выяснилось, что гомологи RAG генов есть даже в геноме морского ежа Strongylocentrotus,
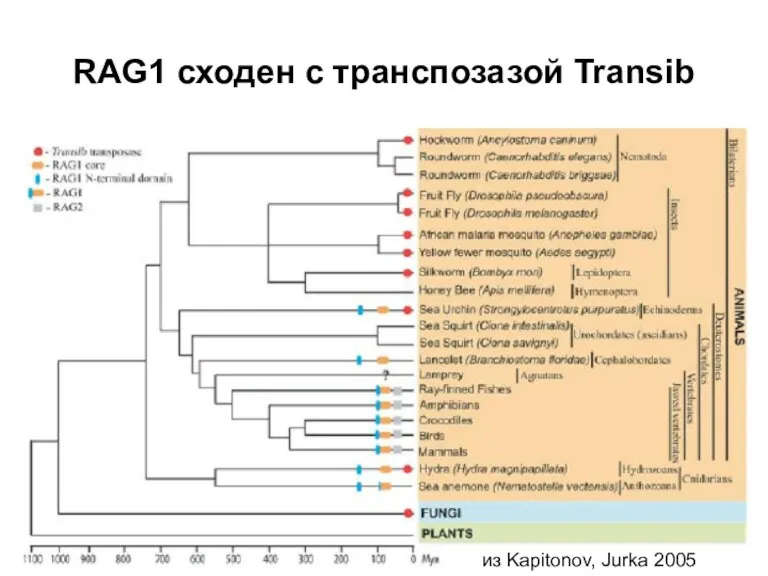
Слайд 23RAG1 сходен с транспозазой Transib
из Kapitonov, Jurka 2005
RAG1 сходен с транспозазой Transib
из Kapitonov, Jurka 2005
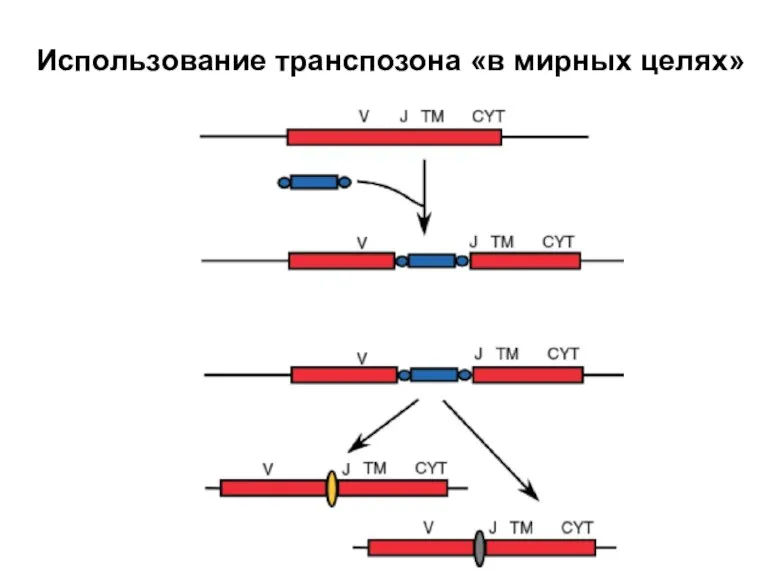
Слайд 24Использование транспозона «в мирных целях»
Использование транспозона «в мирных целях»
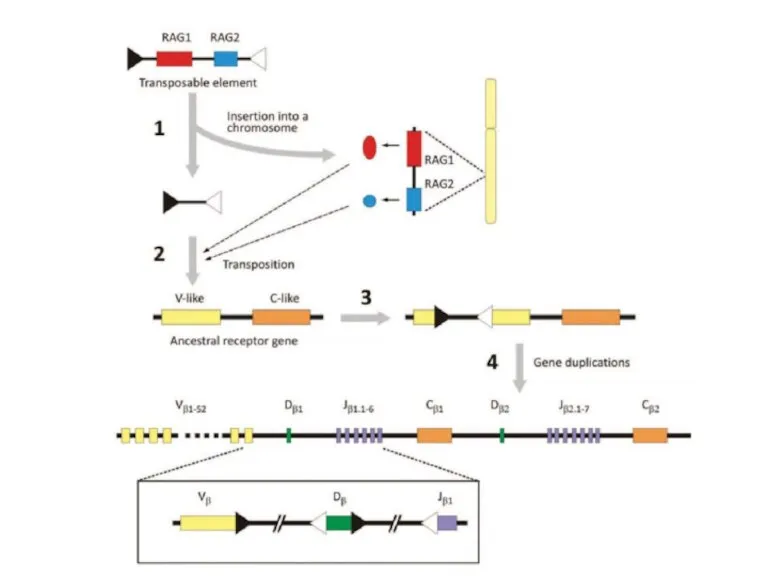
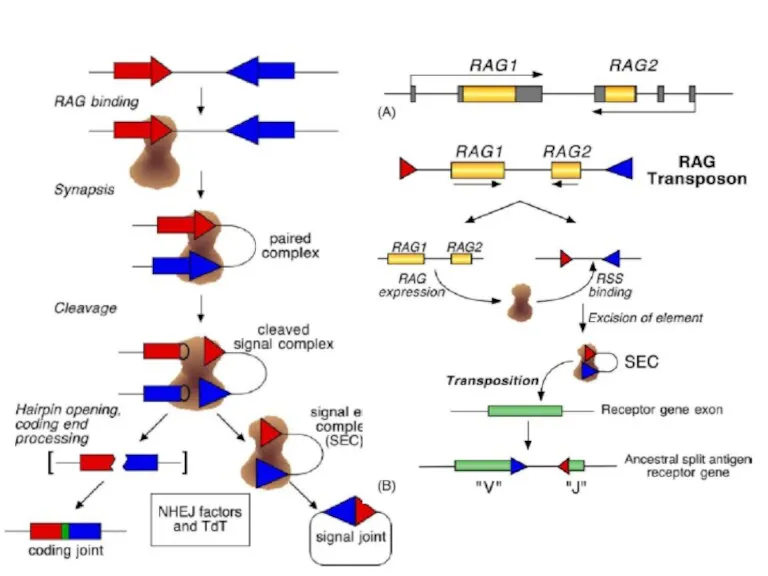
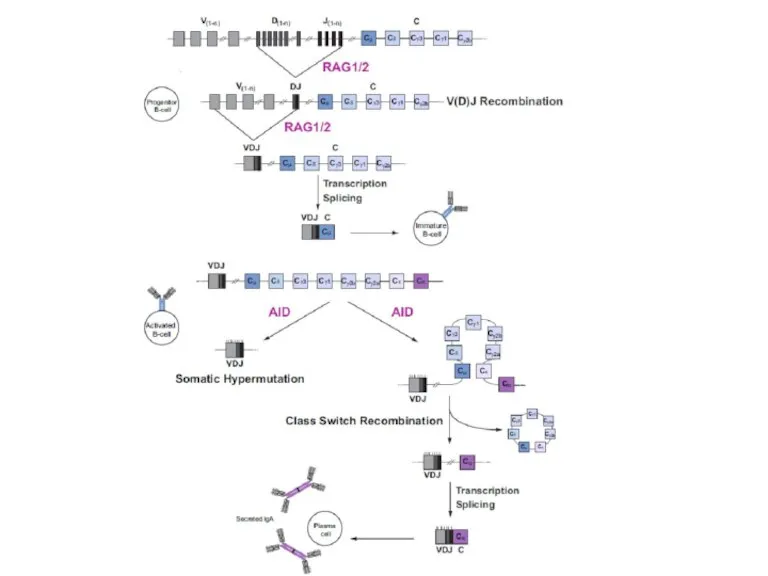
Слайд 28Механизм переключения классов антител
Механизм переключения классов антител
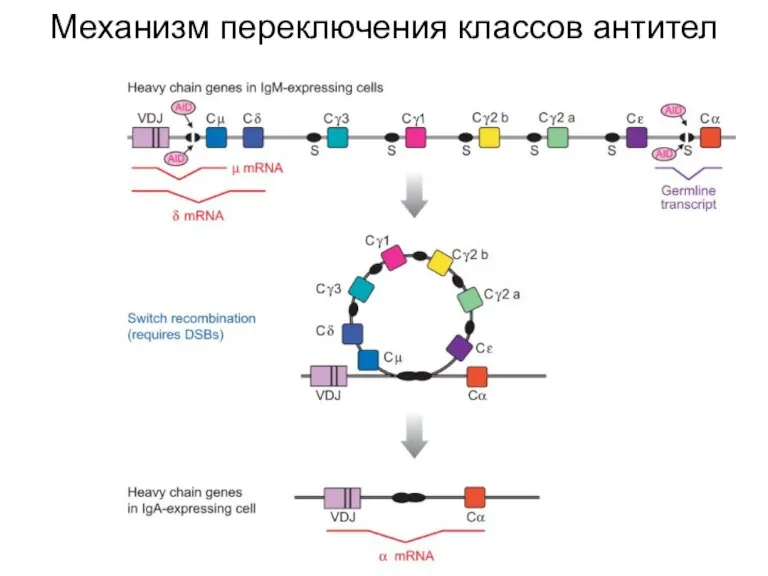
Слайд 29Рецепторы врожденного иммунитета могли возникнуть из рецепторов NK-клеток
Рецепторы врожденного иммунитета могли возникнуть из рецепторов NK-клеток
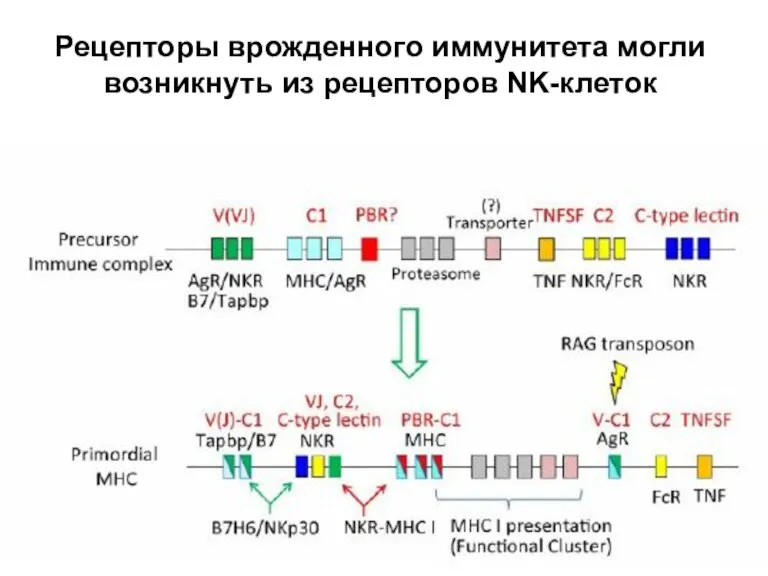

Слайд 31Rag-зависимая рекомбинация или конверсия генов?
Rag-зависимая рекомбинация или конверсия генов?
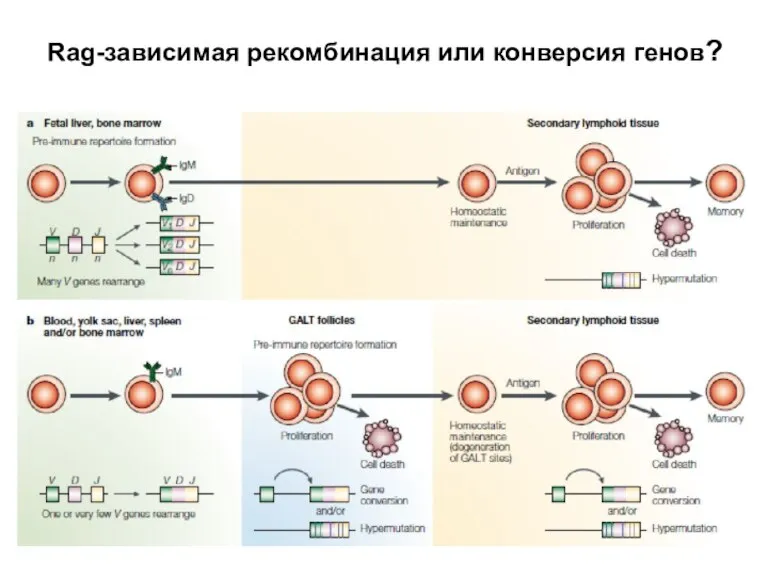
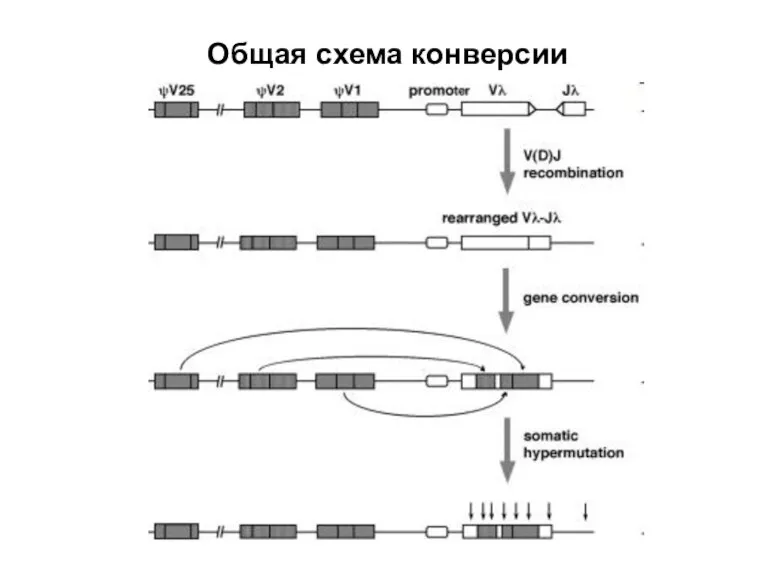
Слайд 32Общая схема конверсии
Общая схема конверсии
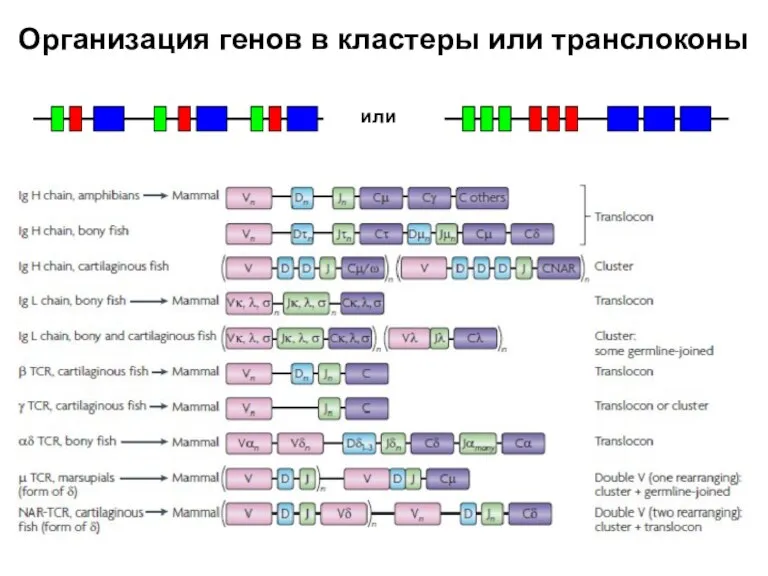
Слайд 33Организация генов в кластеры или транслоконы
или
Организация генов в кластеры или транслоконы
или
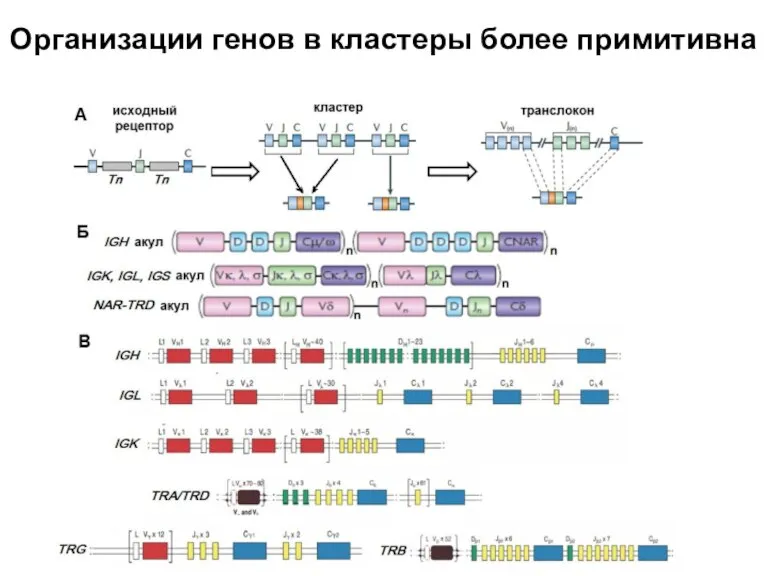
Слайд 34Организации генов в кластеры более примитивна
Организации генов в кластеры более примитивна
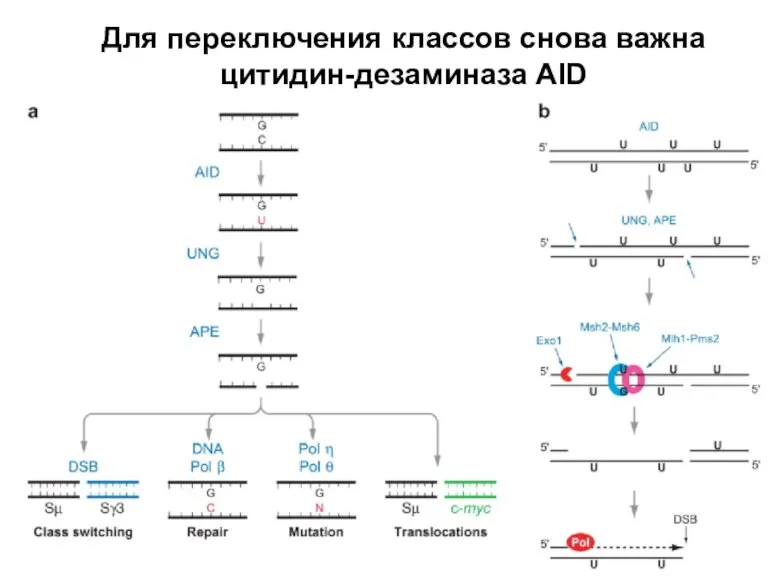
Слайд 35Для переключения классов снова важна цитидин-дезаминаза AID
Для переключения классов снова важна цитидин-дезаминаза AID
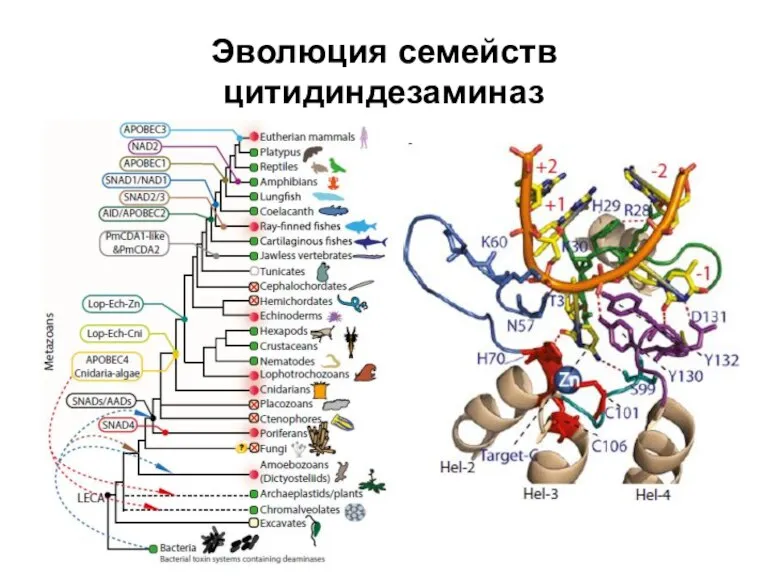
Слайд 36Эволюция семейств цитидиндезаминаз
Эволюция семейств цитидиндезаминаз
 Победа ЖШС-дағы қырдың қызыл және симментал тұқым бұқашықтарының өсіпөнуі мен жетілу ерекшеліктері
Победа ЖШС-дағы қырдың қызыл және симментал тұқым бұқашықтарының өсіпөнуі мен жетілу ерекшеліктері Технологии и материалы озеленения стен
Технологии и материалы озеленения стен Михаил Иванович Грибушин (1832 – 1889)
Михаил Иванович Грибушин (1832 – 1889) Животные и растения Унинского района
Животные и растения Унинского района Структура скелетной мышцы
Структура скелетной мышцы Аттестационная работа. Повышение квалификации учителей химии и биологии в организации проектно-исследовательской деятельности
Аттестационная работа. Повышение квалификации учителей химии и биологии в организации проектно-исследовательской деятельности Признаки двудольных и однодольных
Признаки двудольных и однодольных Иммунная система человека
Иммунная система человека 2 типа разложения органики
2 типа разложения органики Классификация аминокислот в зависимости от природы радикалов. (Лекция 3)
Классификация аминокислот в зависимости от природы радикалов. (Лекция 3) Среда для культивирования бактерий
Среда для культивирования бактерий Ағызылымды суларды тазалау
Ағызылымды суларды тазалау Царства живой природы
Царства живой природы Презентация на тему Аристотель. Заслуги в биологии
Презентация на тему Аристотель. Заслуги в биологии  Анимация рыбка
Анимация рыбка Аппарат Гольджи
Аппарат Гольджи Семена и плоды растений
Семена и плоды растений Частная физиология ЦНС. Медиаторные системы. Ацетилхолин
Частная физиология ЦНС. Медиаторные системы. Ацетилхолин Сохраним пчелу – Сохраним планету!
Сохраним пчелу – Сохраним планету! Удивительный мир растений
Удивительный мир растений Пищеварение в желудке и в кишечнике
Пищеварение в желудке и в кишечнике Рабочая тетрадь. Биология 9 класс
Рабочая тетрадь. Биология 9 класс Деление клетки (митоз)
Деление клетки (митоз) Трудно птицам зимовать, надо птицам помогать!
Трудно птицам зимовать, надо птицам помогать! Пчелы и муравьи – общественные насекомые
Пчелы и муравьи – общественные насекомые Моя любимая порода собак
Моя любимая порода собак Презентация на тему Семейство Астровые или Сложноцветные
Презентация на тему Семейство Астровые или Сложноцветные  Особенности, многообразие и классификация животных. 7 класс
Особенности, многообразие и классификация животных. 7 класс