- Динамическая структура популяции

Содержание
- 2. Методы оценки размеров популяции При проведении любых количественных экологических исследований важно с большой степенью точности дать
- 3. Объективные методы Квадрат. Если установлено число организмов в пределах некоторого числа квадратов, занимающих известную часть общей
- 4. 2) Частота вида. Это вероятность обнаружения определенного вида в пределах любого случайным образом брошенного квадрата в
- 5. 3) Проективное покрытие вида показывает, какая часть почвы занята особями данного вида, и дает оценку площади,
- 6. Прямое наблюдение. Прямой подсчет особей применим не только к сидячим или медленно передвигающимся животным, но и
- 7. Косвенные методы Метод неселективного изъятия В основе использования метода неселективного изъятия лежит явление постепенного снижения вероятности
- 8. Метод неселективного изъятия :
- 9. Методы мечения, выпуска и повторного отлова Он заключается в следующем. Отлавливается выборка животных численностью M, каждое
- 10. Метод многократного отлова и мечения Оценить численность популяции можно и проводя многократные отловы, при этом производя
- 11. В первый раз отловили 58 животных, всех их пометили и отпустили. Во второй раз также отловили
- 12. Нас интересует значение X, при котором Y = 100, т.е. при какой численности все особи в
- 13. Если теперь построить уравнение зависимости доли меченых организмов от совокупного числа меченых то полученная зависимость может
- 14. Половая структура популяции это соотношение между самцами и самками в популяции. Оно видоспецефично и напрямую зависит
- 15. В популяциях копытных соотношение между самками и самцами обычно составляет от 1:2 до 1:5, у приматов
- 16. Рождаемость Рождаемость (скорость рождения) — - это число особей (ΔNn), рождающихся в популяции за единицу времени
- 17. Плодовитость Способность к размножению (измеряется числом гамет, семян и т.д.). Плодовитость сильно зависит от степени развития
- 18. Смертность Смертность — способность популяции снижать численность особей в популяции за счет их гибели, причем причина
- 19. Смертность В реальных условиях среды наблюдается реализованная, или экологическая смертность. Абсолютная смертность выражается числом особей, умерших
- 20. Скорость изменения численности популяции ΔN/Δt Удельная скорость изменения численности r = b – d Если b
- 21. Выживаемость Это доля особей, доживших до определенного момента времени Зависимость числа выживших особей от времени (или
- 22. Кривые выживания Кривая типа I имеет почти линейную форму и характерна для организмов с равномерной смертностью
- 23. Кривая типа II Она характерна для организмов, которые в основном гибнут на начальных этапах своего развитияТакой
- 24. Экологические стратегии выживания В 1938 г эколог А. Роменский выделил три основные стратегии выживания: 1) виоленты
- 25. Динамика роста численности популяций 2 основных типа кривых роста численности Экспоненциальный рост численности Nt = N0ert,
- 26. Экспоненциальный рост (J-образная кривая) Это рост численности особей в неизменяющихся условиях
- 27. Воздействие экологических факторов на скорость роста популяции может довести численность популяции до стабильной (r=0) или ее
- 28. Логистический рост При N = K r=0
- 29. K- и r-стратегии популяций Стабильная динамика. К-стратегия обеспечивает выживание за счет качественного совершенствования. Равновесные популяции Нестабильная
- 31. Популяционные волны Периодические и непериодические колебания численности популяций под влиянием абиотических и биотических факторов среды называются
- 33. Скачать презентацию
Слайд 2Методы оценки размеров популяции
При проведении любых количественных экологических исследований важно с большой
Методы оценки размеров популяции
При проведении любых количественных экологических исследований важно с большой

Слайд 3Объективные методы
Квадрат. Если установлено число организмов в пределах некоторого числа квадратов, занимающих
Объективные методы
Квадрат. Если установлено число организмов в пределах некоторого числа квадратов, занимающих

Слайд 42) Частота вида. Это вероятность обнаружения определенного вида в пределах любого случайным образом брошенного
2) Частота вида. Это вероятность обнаружения определенного вида в пределах любого случайным образом брошенного

Слайд 53) Проективное покрытие вида показывает, какая часть почвы занята особями данного вида, и
3) Проективное покрытие вида показывает, какая часть почвы занята особями данного вида, и

Слайд 6Прямое наблюдение. Прямой подсчет особей применим не только к сидячим или медленно
Прямое наблюдение. Прямой подсчет особей применим не только к сидячим или медленно

Слайд 7
Косвенные методы
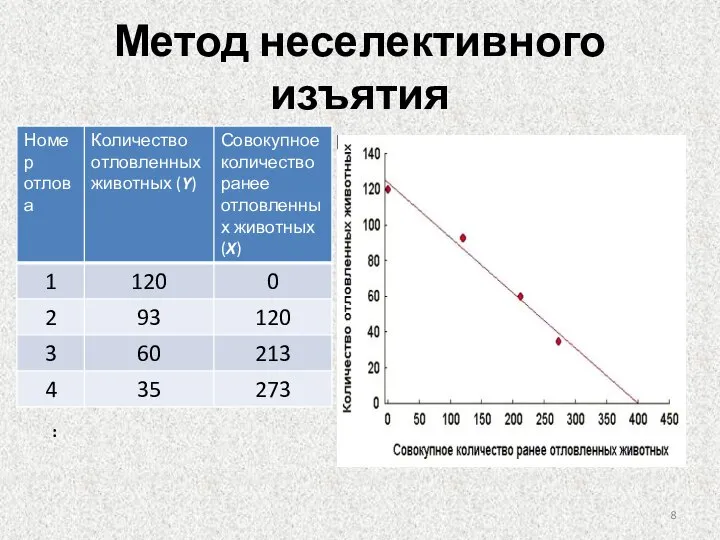
Метод неселективного изъятия
В основе использования метода неселективного изъятия лежит явление постепенного снижения вероятности
Косвенные методы
Метод неселективного изъятия
В основе использования метода неселективного изъятия лежит явление постепенного снижения вероятности

Слайд 8Метод неселективного изъятия
:
Метод неселективного изъятия
:
Слайд 9
Методы мечения, выпуска и повторного отлова
Он заключается в следующем. Отлавливается выборка животных
Методы мечения, выпуска и повторного отлова
Он заключается в следующем. Отлавливается выборка животных

Слайд 10
Метод многократного отлова и мечения
Оценить численность популяции можно и проводя многократные отловы,
Метод многократного отлова и мечения
Оценить численность популяции можно и проводя многократные отловы,

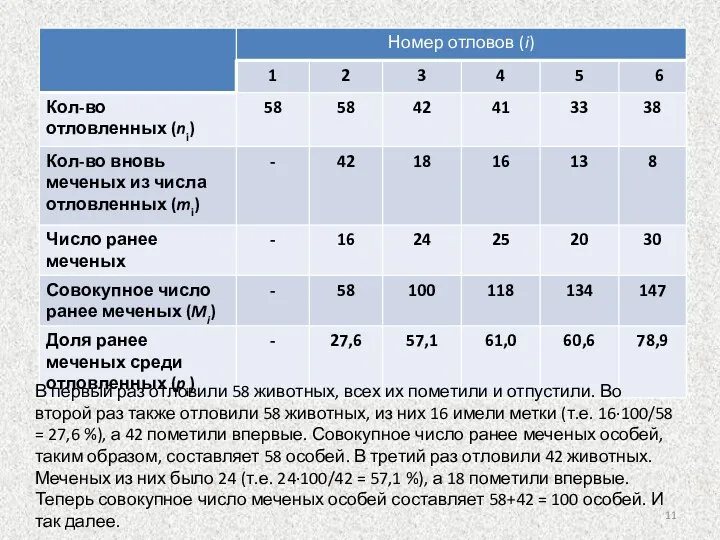
Слайд 11В первый раз отловили 58 животных, всех их пометили и отпустили. Во
В первый раз отловили 58 животных, всех их пометили и отпустили. Во
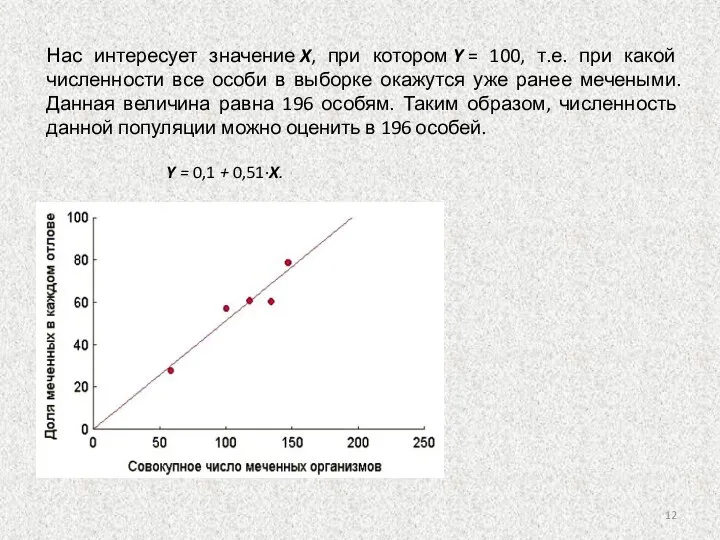
Слайд 12Нас интересует значение X, при котором Y = 100, т.е. при какой численности все особи
Нас интересует значение X, при котором Y = 100, т.е. при какой численности все особи
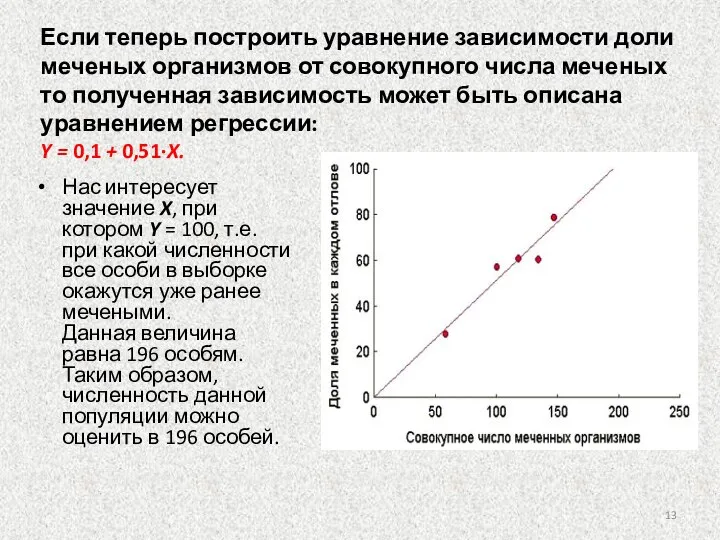
Слайд 13Если теперь построить уравнение зависимости доли меченых организмов от совокупного числа меченых
Если теперь построить уравнение зависимости доли меченых организмов от совокупного числа меченых
Слайд 14Половая структура популяции
это соотношение между самцами и самками в популяции. Оно видоспецефично
Половая структура популяции
это соотношение между самцами и самками в популяции. Оно видоспецефично

Слайд 15В популяциях копытных соотношение между самками и самцами обычно составляет от 1:2
В популяциях копытных соотношение между самками и самцами обычно составляет от 1:2

Слайд 16Рождаемость
Рождаемость (скорость рождения) — - это число особей (ΔNn), рождающихся в популяции
Рождаемость
Рождаемость (скорость рождения) — - это число особей (ΔNn), рождающихся в популяции

Слайд 17Плодовитость
Способность к размножению (измеряется числом гамет, семян и т.д.). Плодовитость сильно зависит от
Плодовитость
Способность к размножению (измеряется числом гамет, семян и т.д.). Плодовитость сильно зависит от

Слайд 18
Смертность
Смертность — способность популяции снижать численность особей в популяции за счет их гибели,
Смертность
Смертность — способность популяции снижать численность особей в популяции за счет их гибели,

Слайд 19Смертность
В реальных условиях среды наблюдается реализованная, или экологическая смертность.
Абсолютная смертность выражается числом особей,
Смертность
В реальных условиях среды наблюдается реализованная, или экологическая смертность.
Абсолютная смертность выражается числом особей,

Слайд 20Скорость изменения численности популяции ΔN/Δt
Удельная скорость изменения численности r = b –
Скорость изменения численности популяции ΔN/Δt
Удельная скорость изменения численности r = b –

Слайд 21Выживаемость
Это доля особей, доживших до определенного момента времени
Зависимость числа выживших особей
Выживаемость
Это доля особей, доживших до определенного момента времени
Зависимость числа выживших особей

Слайд 22Кривые выживания
Кривая типа I имеет почти линейную форму и характерна для организмов с равномерной
Кривые выживания
Кривая типа I имеет почти линейную форму и характерна для организмов с равномерной
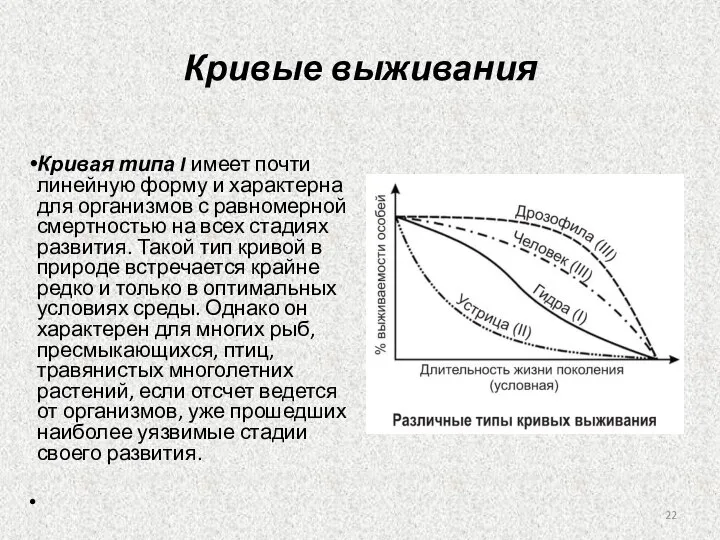
Слайд 23Кривая типа II Она характерна для организмов, которые в основном гибнут на начальных
Кривая типа II Она характерна для организмов, которые в основном гибнут на начальных

Слайд 24Экологические стратегии выживания
В 1938 г эколог А. Роменский выделил три основные стратегии
Экологические стратегии выживания
В 1938 г эколог А. Роменский выделил три основные стратегии

Слайд 25Динамика роста численности популяций
2 основных типа кривых роста численности
Экспоненциальный рост численности
Динамика роста численности популяций
2 основных типа кривых роста численности
Экспоненциальный рост численности

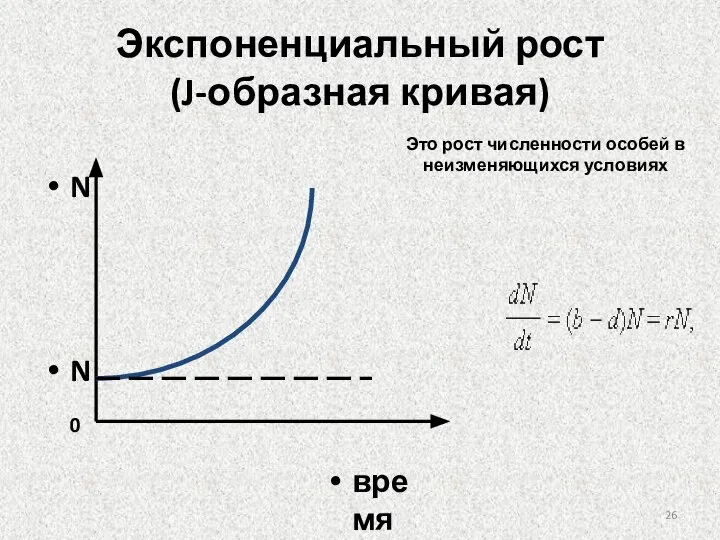
Слайд 26Экспоненциальный рост
(J-образная кривая)
Это рост численности особей в неизменяющихся условиях
Экспоненциальный рост
(J-образная кривая)
Это рост численности особей в неизменяющихся условиях
Слайд 27Воздействие экологических факторов на скорость роста популяции может довести численность популяции до
Воздействие экологических факторов на скорость роста популяции может довести численность популяции до

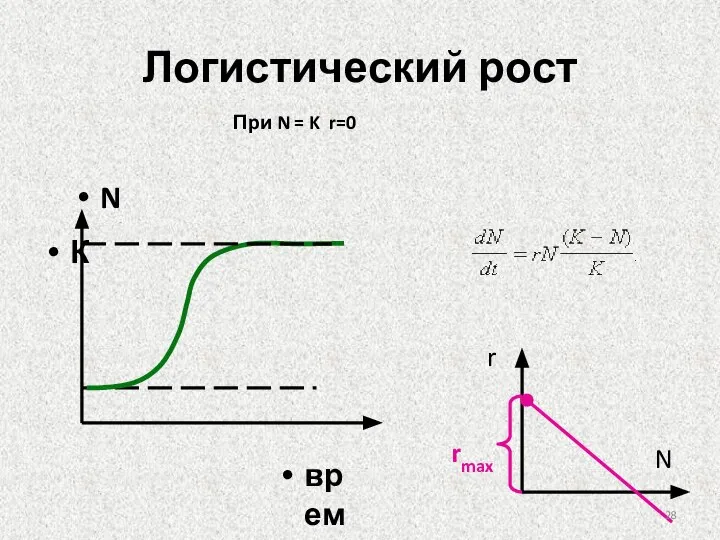
Слайд 28Логистический рост
При N = K r=0
Логистический рост
При N = K r=0
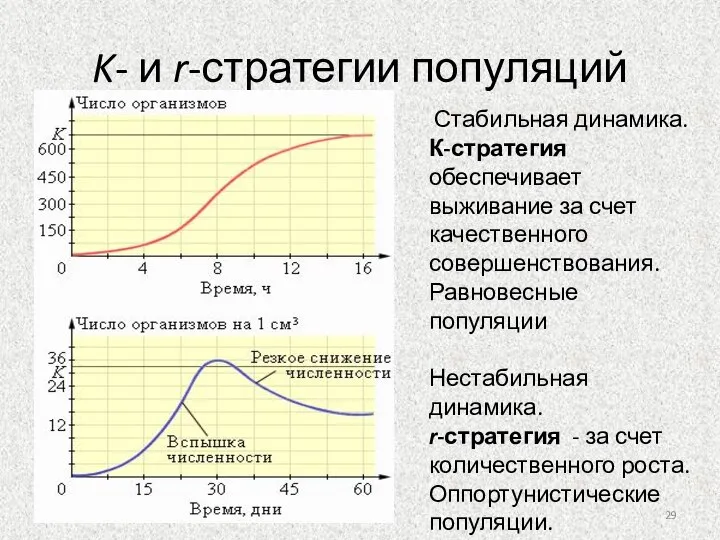
Слайд 29K- и r-стратегии популяций
Стабильная динамика.
К-стратегия обеспечивает выживание за счет качественного
K- и r-стратегии популяций
Стабильная динамика.
К-стратегия обеспечивает выживание за счет качественного

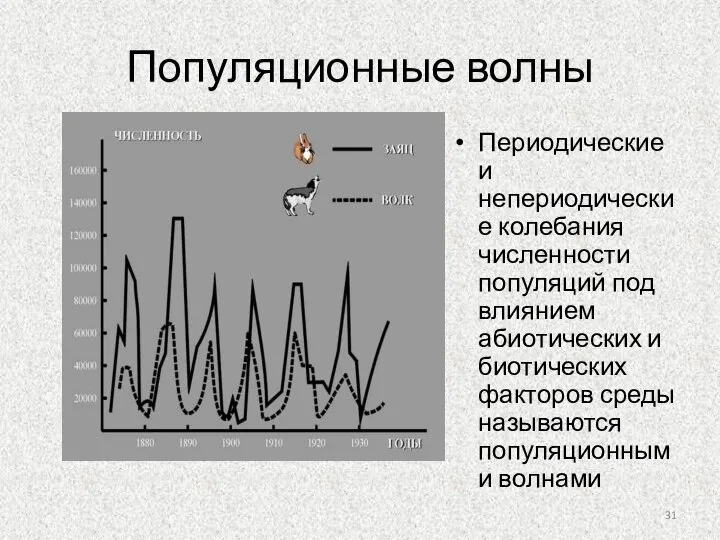
Слайд 31Популяционные волны
Периодические и непериодические колебания численности популяций под влиянием абиотических и биотических
Популяционные волны
Периодические и непериодические колебания численности популяций под влиянием абиотических и биотических
 Итоги экогода в учебном заведении
Итоги экогода в учебном заведении Тайны Арчекаса. Экологическая тропа
Тайны Арчекаса. Экологическая тропа Болота и луга
Болота и луга Лесной питомник Дедовичи, Санкт-Петербург
Лесной питомник Дедовичи, Санкт-Петербург Лекция 1 Основы
Лекция 1 Основы Мастер’ОК. Трудовой отряд
Мастер’ОК. Трудовой отряд Вторичная переработка мусора
Вторичная переработка мусора Air Pollution
Air Pollution Человек и экология. Брейн-ринг
Человек и экология. Брейн-ринг Экологическая ферма
Экологическая ферма Межпопуляционные отношения в экосистемах
Межпопуляционные отношения в экосистемах Общество и природа
Общество и природа Энергоэффективность и самочувствие людей в зелёном офисе
Энергоэффективность и самочувствие людей в зелёном офисе Кейс Развитие территорий. Направление: Берегу природу
Кейс Развитие территорий. Направление: Берегу природу Экопарк на Гребном канале, г. Нижний Новгород. Проект
Экопарк на Гребном канале, г. Нижний Новгород. Проект Экология. 1 класс
Экология. 1 класс Мир без мусора
Мир без мусора Человек в биосфере
Человек в биосфере Петербург: от столицы культурной – к столице экологической
Петербург: от столицы культурной – к столице экологической Восстановление флоры и фауны рек и озер при
Восстановление флоры и фауны рек и озер при Архитектурная экология
Архитектурная экология Эколого-культурная тропа Селенитовый путь с. Красный Ясыл
Эколого-культурная тропа Селенитовый путь с. Красный Ясыл компост-мобиль. Умная пятница
компост-мобиль. Умная пятница Введение в ландшафтную экологию
Введение в ландшафтную экологию Экологические проблемы
Экологические проблемы Das planet erde ist in gefahr
Das planet erde ist in gefahr Правовой режим зон чрезвычайной экологической ситуации и зон экологического бедствия
Правовой режим зон чрезвычайной экологической ситуации и зон экологического бедствия Повышение эффективности расширенной ответственности производителя
Повышение эффективности расширенной ответственности производителя