- Моделирование экологических взаимодействий биоценоза

Содержание
- 2. ДИНАМИКА ПОПУЛЯЦИЙ МЕЖВИДОВЫЕ ВЗАИМОДЕЙСТВИЯ КОМПОНЕНТОВ БИОЦЕНОЗА МОДЕЛЬ ПРОГНОЗА И УПРАВЛЕНИЯ ДИНАМИКОЙ ПОПУЛЯЦИЙ Лекция 9 МОДЕЛИРОВАНИЕ ЭКОЛОГИЧЕСКИХ
- 3. ДИНАМИКА ПОПУЛЯЦИЙ Системная экология изучает популяции как самостоятельные подсистемы в рамках экологической системы, объединяющей их с
- 4. Например, разбив популяцию на группы самок и самцов, получают состав популяции. При этом структура популяции будет
- 5. Изменение плотности популяции происходит в результате взаимодействия четырех процессов: размножение, гибель, иммиграция, эмиграция X(tk+1) = X(tk)+(Vb–Vd)+(Vi–Ve)Δt,
- 6. Однако, количественное описание функций скорости иммиграции и эмиграции связано с необходимостью учета свойств не только данной,
- 7. Однако, количественное описание функций скорости иммиграции и эмиграции связано с необходимостью учета свойств не только данной,
- 8. В полевых условиях определить влияние процессов размножения и гибели на динамику популяций весьма затруднительно, X(tk+1) =
- 9. для независимой от плотности популяции компоненты r существует оптимальная комбинация факторов внешней и внутренней среды, при
- 10. rm=1,0 rm=0,8 rm=0,6 rm=0,4 rm=0,2 Рис. 83. Возрастание плотности популяций с разными биотическими потенциалами.
- 11. Если бы условия среды сохранялись оптимальными постоянно, то прирост популяции шел бы в геометрической прогрессии непрерывно.
- 12. Но ресурсы среды ограничены, поэтому при отклонении экологических факторов от оптимальных рождаемость снижается, а смертность возрастает.
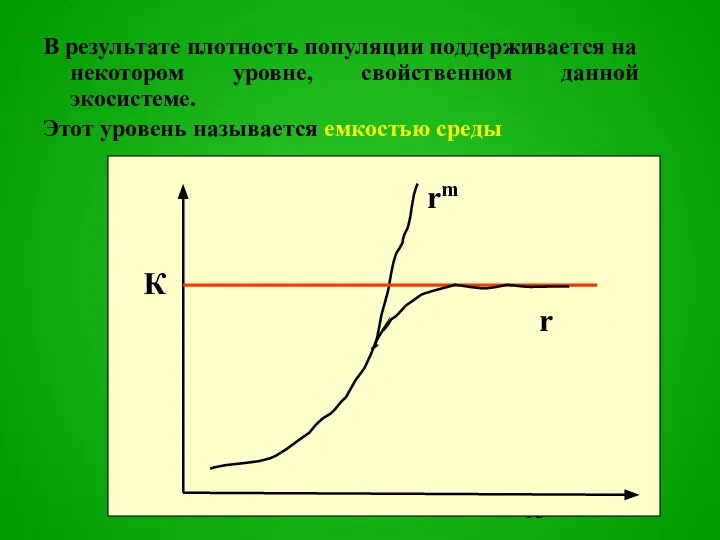
- 13. В результате плотность популяции поддерживается на некотором уровне, свойственном данной экосистеме. Этот уровень называется емкостью среды
- 14. Плотность популяции в среде с ограниченными ресурсами описывается уравнением, называемым логистическим K X(t1) = –––––––––––––––––––––––––––––, 1+(K–X(t0))/X(t0))
- 15. Для прогноза применить это уравнение можно, если предварительно выяснить зависимость Х от К при различном сочетании
- 16. В практике сельскохозяйственного производства агроном и консультант часто имеют дело с вредителями, болезнями и сорняками на
- 17. Динамика численности насекомых и пауков зависит от энтальпии среды. Если взять ее как параметр времени и
- 18. МЕЖВИДОВЫЕ ВЗАИМОДЕЙСТВИЯ КОМПОНЕНТОВ БИОЦЕНОЗА Прогноз динамики фитофагов гораздо точнее и реальнее, если ввести в него поправку
- 19. Модель, учитывающая взаимодействия между популяциями, называется биоценотической моделью динамики популяций X(tk+1) = X(tk)+(Vb–Vd)+(Vi–Ve)Δt Фитофаг Энтомофаг X(tk+1)
- 20. Если дополнительно ввести еще абиотические факторы, то получим экосистемную модель динамики популяций X(tk+1) = X(tk)+(Vb–Vd)+(Vi–Ve)Δt Фитофаг
- 21. Рождаемость (Vb) и смертность (Vd) зависят от многих факторов, основными из которых являются биотические взаимодействия. Последние
- 22. Частично преодолеть эти трудности оказалось возможным, когда взаимодействия стали классифицировать не по качественному разнообразию их механизмов,
- 23. КЛАССИФИКАЦИЯ ВЗАИМОДЕЙСТВИЯ ДВУХ ВИДОВ БИОЦЕНОЗА
- 24. Популяции двух видов не оказывают непосредственного воздействия друг на друга. Нейтрализм (0, 0) 0 0 Такой
- 25. один из видов взаимодействующей пары в биоценозе оказывает отрицательное воздействие на рост другого, хотя сам не
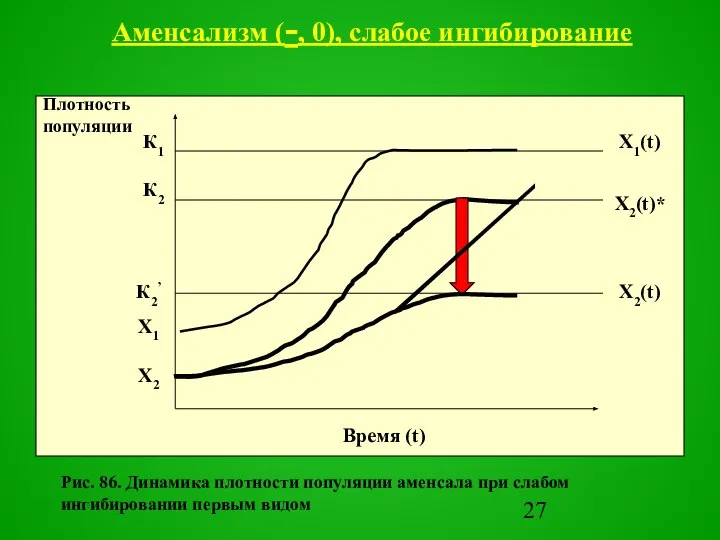
- 26. При слабом ингибировании система стремится к устойчивому стационарному состоянию, в котором численность аменсала будет меньше, чем
- 27. Рис. 86. Динамика плотности популяции аменсала при слабом ингибировании первым видом Аменсализм (-, 0), слабое ингибирование
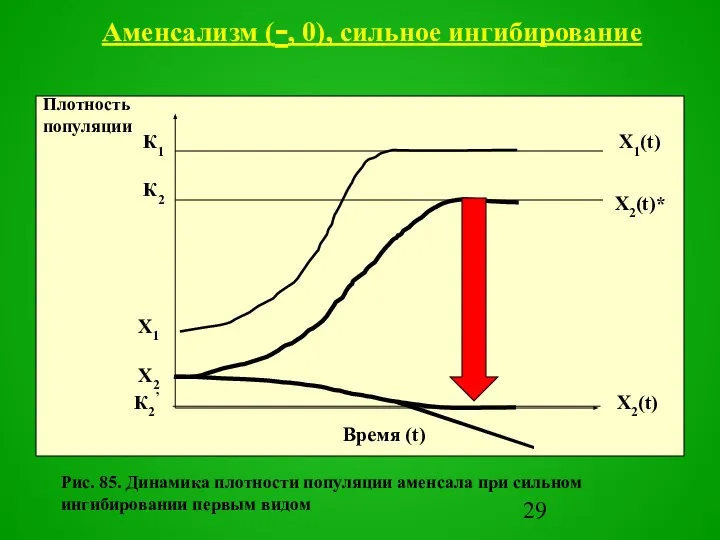
- 28. При сильном ингибировании популяция аменсала полностью погибает (элиминируется) Аменсализм (-, 0) сильное ингибирование
- 29. Рис. 85. Динамика плотности популяции аменсала при сильном ингибировании первым видом Аменсализм (-, 0), сильное ингибирование
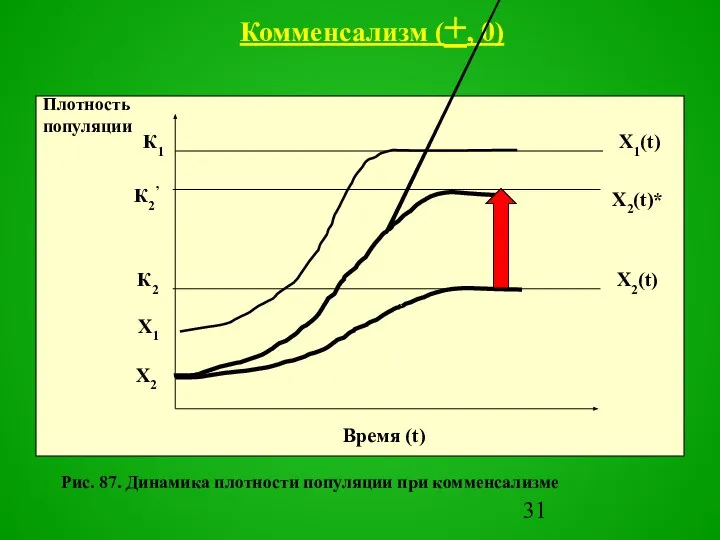
- 30. первый вид, называемый «хозяином», положительно воздействует на второй, называемый «комменсалом» («нахлебником»), а второй не влияет на
- 31. Рис. 87. Динамика плотности популяции при комменсализме Комменсализм (+, 0)
- 32. любое взаимно отрицательное отношение между видами. Конкуренция (интерференция (-, -) - - вид, получающий какое-либо преимущество
- 33. Частными случаями конкуренции являются: – конкуренция за тот или иной ограниченный ресурс (соперничество). Среди растений это
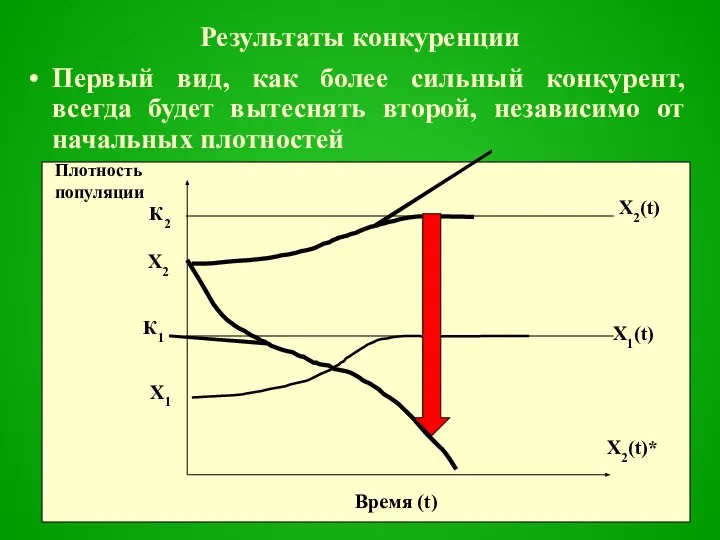
- 34. Результаты конкуренции Первый вид, как более сильный конкурент, всегда будет вытеснять второй, независимо от начальных плотностей
- 35. Результаты конкуренции Второй вид, будучи более сильным конкурентом, вытесняет первый независимо от начальных плотностей
- 36. Результаты конкуренции Если виды одинаково конкурентноспособны, то независимо от исходных плотностей система приходит в устойчивое состояние
- 37. Результаты конкуренции Если емкости внешней среды не лимитируют рост плотности популяций видов, то исход конкуренции определяется
- 38. Таким образом, для обеспечения устойчивости конкурентного равновесия в системе, если оно возможно, достаточно, чтобы самоингибирование каждой
- 39. отношения между двумя видами, при которых увеличение (или уменьшение) плотности популяции первого («жертвы») влечет за собой
- 40. «Жертва-эксплуататор» (+, -) Вид-эксплуататор не способен прокормиться за счет данной жертвы и со временем вымирает, а
- 41. Типичный пример – применение биологического метода защиты растений, когда внедренный в экосистему хищник или паразит не
- 42. «Жертва-эксплуататор» (+, -) Вид-эксплуататор слишком эффективен в поисках жертвы и быстро размножается, в результате чего он
- 43. Ситуация “козел в огороде” характерна для монофагов, используемых как средство биологической защиты растений.
- 44. «Жертва-эксплуататор» (+, -) Существует единственная устойчивая комбинация плотностей жертвы и эксплуататора, и из любого начального состояния
- 45. Это ситуация, когда эксплуататор достаточно эффективен и может быстро снижать численность жертвы до низкого стационарного уровня,
- 46. «Жертва-эксплуататор» (+, -) Существует единственная устойчивая комбинация плотностей популяций жертвы и эксплуататора, при котором после переходного
- 47. Например, взаимодействие растений и копытных растительноядных животных, которые периодически мигрируют на другие участки. В агроэкосистемах –
- 48. «Жертва-эксплуататор» (+, -) Из любого начального состояния траектория системы стремится к единственно замкнутой траектории, движение по
- 49. Такие автоколебания характерны для ненарушенных внешними факторами систем, где высокая степень саморегуляции. Синусоидальная траектория характеризует внутренне
- 50. взаимно положительное влияние, которое в конечном счете проявляется в том, что увеличение (снижение) численности любого из
- 53. Все многообразие моделей прогнозов по шкале времени можно разделить на прогнозы МОДЕЛЬ ПРОГНОЗА И УПРАВЛЕНИЯ ДИНАМИКОЙ
- 54. Методы прогнозов
- 55. Климограмма это двумерная экологическая ниша, где учитываются температура воздуха
- 56. Климограмма
- 57. При помощи климограммы можно установить Обитает ли организм на данной территории? Осадки, мм
- 58. При помощи климограммы можно установить Будет ли обитать интродуцированный организм на данной территории? Осадки, мм
- 59. При помощи климограммы можно установить Если это фитофаг, будет ли он вредителем? Осадки, мм
- 60. При помощи климограммы можно установить Будет ли массовое размножение организма? Осадки, мм
- 61. Например, размножение тлей зависит от метеорологических условий и фазы разввития растения. Низкие температуры и обильные частые
- 62. В 1988 году в Костромской области наблюдалось массовое размножение тли. Воспользовавшись этим обстоятельством, мы составили климограмму
- 63. оптимальные для тли условия складываются при среднемесячной температуре воздуха –11…+19°С и сумме осадков за месяц от
- 64. Анализ климатических условий показывает, что ежегодному массовому размножению тли в Костромской области препятствуют пониженные температуры января
- 65. Методы прогнозов
- 66. Сезонный и оперативный прогнозы динамики составляются при помощи математических функций. Y=F(x) Для обеспечения возможности прогнозирования в
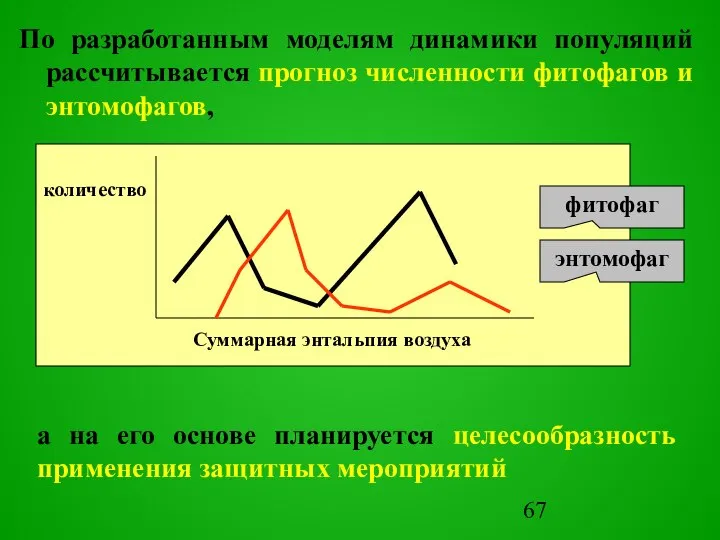
- 67. По разработанным моделям динамики популяций рассчитывается прогноз численности фитофагов и энтомофагов, количество Суммарная энтальпия воздуха а
- 68. Ta * qa * Рис. 99. Структурная схема блока управления энтомосанитарным состоянием агроэкосистемы. Энтомофаг:фитофаг на критическую
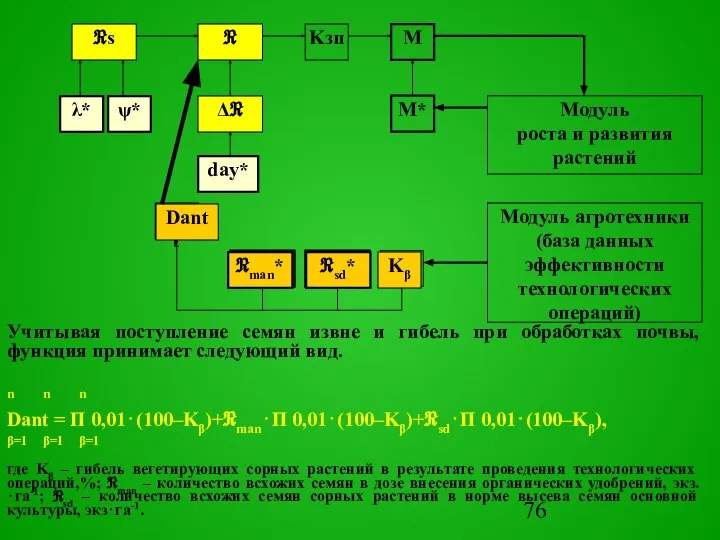
- 69. Прогноз засоренности посевов Прогноз засоренности посевов составить несколько проще по причине ограниченной способности семян и вегетирующих
- 70. М* М day* ψ* Kзп Модуль агротехники (база данных эффективности технологических операций) Модуль роста и развития
- 71. М* М day* ψ* Kзп Модуль агротехники (база данных эффективности технологических операций) Модуль роста и развития
- 72. М* М day* ψ* Kзп Модуль агротехники (база данных эффективности технологических операций) Модуль роста и развития
- 73. М* М Kзп Модуль агротехники (база данных эффективности технологических операций) Модуль роста и развития растений Dant
- 74. М* М Kзп Модуль агротехники (база данных эффективности технологических операций) Модуль роста и развития растений Dant
- 75. 20% 30% 80% 50%
- 76. М* М Kзп Модуль агротехники (база данных эффективности технологических операций) Модуль роста и развития растений Dant
- 77. М* М Kзп Модуль агротехники (база данных эффективности технологических операций) Модуль роста и развития растений λ*
- 78. М* М Kзп Модуль агротехники (база данных эффективности технологических операций) Модуль роста и развития растений λ*
- 79. Тогда малопараметрическая модель прогноза урожайности с учетом засоренности посевов запишется в виде Y(Q,W,t,NPK,pH,Зп) = Y(Q,W,t,NPK,pH)⋅Кзп
- 81. Скачать презентацию
Слайд 2ДИНАМИКА ПОПУЛЯЦИЙ
МЕЖВИДОВЫЕ ВЗАИМОДЕЙСТВИЯ КОМПОНЕНТОВ БИОЦЕНОЗА
МОДЕЛЬ ПРОГНОЗА И УПРАВЛЕНИЯ ДИНАМИКОЙ ПОПУЛЯЦИЙ
Лекция 9
МОДЕЛИРОВАНИЕ
ЭКОЛОГИЧЕСКИХ ВЗАИМОДЕЙСТВИЙ
БИОЦЕНОЗА
ДИНАМИКА ПОПУЛЯЦИЙ
МЕЖВИДОВЫЕ ВЗАИМОДЕЙСТВИЯ КОМПОНЕНТОВ БИОЦЕНОЗА
МОДЕЛЬ ПРОГНОЗА И УПРАВЛЕНИЯ ДИНАМИКОЙ ПОПУЛЯЦИЙ
Лекция 9
МОДЕЛИРОВАНИЕ
ЭКОЛОГИЧЕСКИХ ВЗАИМОДЕЙСТВИЙ
БИОЦЕНОЗА

Слайд 3ДИНАМИКА ПОПУЛЯЦИЙ
Системная экология изучает популяции как самостоятельные подсистемы в рамках экологической системы,
ДИНАМИКА ПОПУЛЯЦИЙ
Системная экология изучает популяции как самостоятельные подсистемы в рамках экологической системы,

Слайд 4Например, разбив популяцию на группы самок и самцов, получают состав популяции.
При этом
Например, разбив популяцию на группы самок и самцов, получают состав популяции.
При этом

Слайд 5Изменение плотности популяции происходит в результате взаимодействия четырех процессов: размножение, гибель, иммиграция,
Изменение плотности популяции происходит в результате взаимодействия четырех процессов: размножение, гибель, иммиграция,

Слайд 6Однако, количественное описание функций скорости иммиграции и эмиграции связано с необходимостью учета
Однако, количественное описание функций скорости иммиграции и эмиграции связано с необходимостью учета

Слайд 7Однако, количественное описание функций скорости иммиграции и эмиграции связано с необходимостью учета
Однако, количественное описание функций скорости иммиграции и эмиграции связано с необходимостью учета

Слайд 8В полевых условиях определить влияние процессов размножения и гибели на динамику популяций
В полевых условиях определить влияние процессов размножения и гибели на динамику популяций

Слайд 9для независимой от плотности популяции компоненты r существует оптимальная комбинация факторов внешней
для независимой от плотности популяции компоненты r существует оптимальная комбинация факторов внешней

Слайд 10rm=1,0
rm=0,8
rm=0,6
rm=0,4
rm=0,2
Рис. 83. Возрастание плотности популяций с разными биотическими потенциалами.
rm=1,0
rm=0,8
rm=0,6
rm=0,4
rm=0,2
Рис. 83. Возрастание плотности популяций с разными биотическими потенциалами.
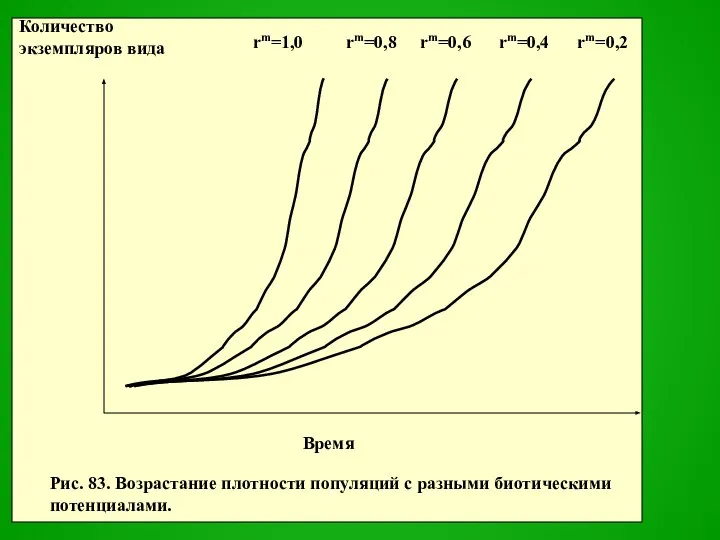
Слайд 11Если бы условия среды сохранялись оптимальными постоянно, то прирост популяции шел бы
Если бы условия среды сохранялись оптимальными постоянно, то прирост популяции шел бы
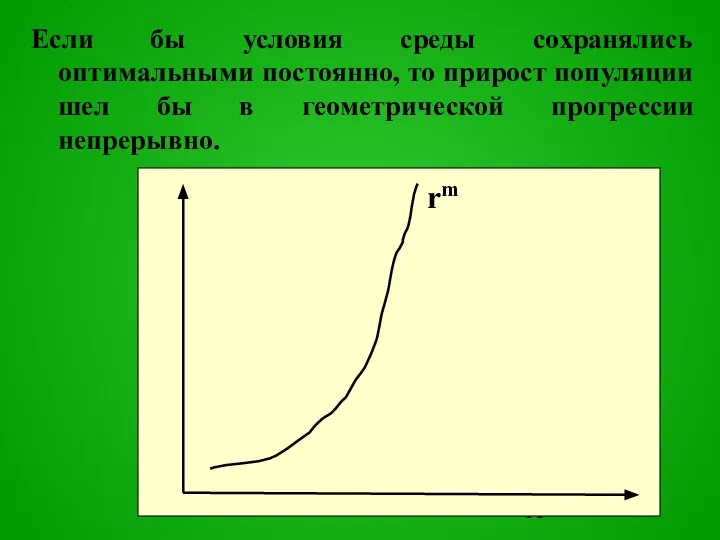
Слайд 12Но ресурсы среды ограничены, поэтому при отклонении экологических факторов от оптимальных рождаемость
Но ресурсы среды ограничены, поэтому при отклонении экологических факторов от оптимальных рождаемость
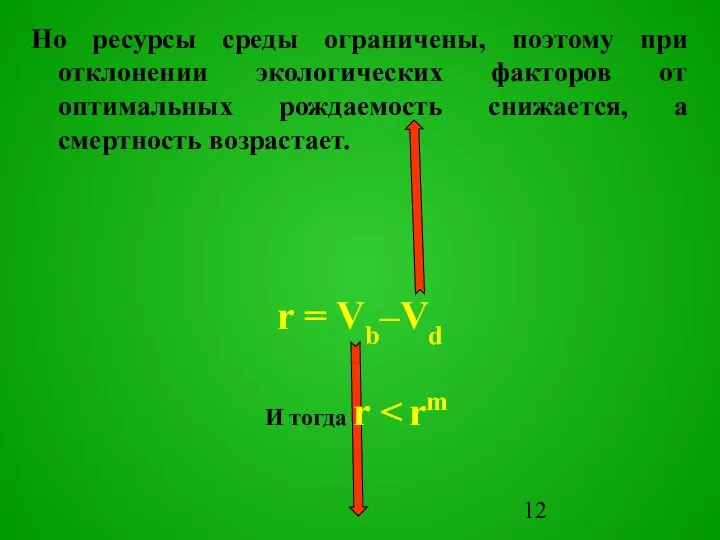
Слайд 13В результате плотность популяции поддерживается на некотором уровне, свойственном данной экосистеме.
Этот
В результате плотность популяции поддерживается на некотором уровне, свойственном данной экосистеме.
Этот
Слайд 14Плотность популяции в среде с ограниченными ресурсами описывается уравнением, называемым логистическим
K
X(t1)
Плотность популяции в среде с ограниченными ресурсами описывается уравнением, называемым логистическим
K
X(t1)

Слайд 15Для прогноза применить это уравнение можно, если предварительно выяснить зависимость Х от
Для прогноза применить это уравнение можно, если предварительно выяснить зависимость Х от

Слайд 16В практике сельскохозяйственного производства агроном и консультант часто имеют дело с вредителями,
В практике сельскохозяйственного производства агроном и консультант часто имеют дело с вредителями,

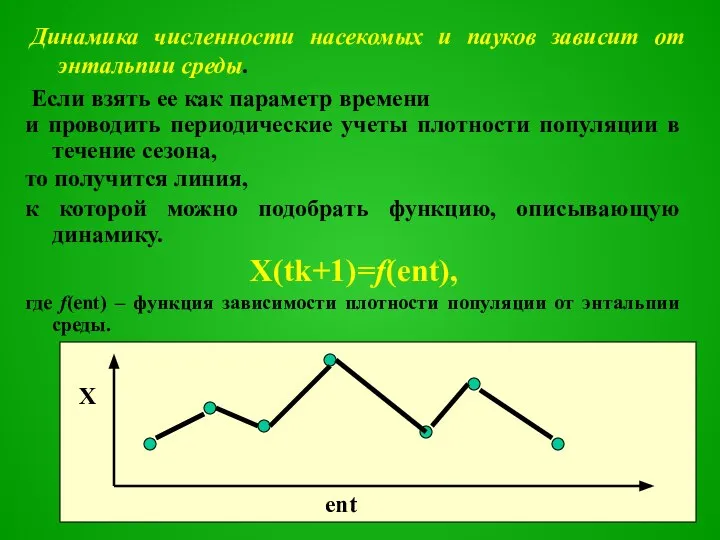
Слайд 17Динамика численности насекомых и пауков зависит от энтальпии среды.
Если взять ее как
Динамика численности насекомых и пауков зависит от энтальпии среды.
Если взять ее как
Слайд 18МЕЖВИДОВЫЕ ВЗАИМОДЕЙСТВИЯ КОМПОНЕНТОВ БИОЦЕНОЗА
Прогноз динамики фитофагов гораздо точнее и реальнее, если ввести
МЕЖВИДОВЫЕ ВЗАИМОДЕЙСТВИЯ КОМПОНЕНТОВ БИОЦЕНОЗА
Прогноз динамики фитофагов гораздо точнее и реальнее, если ввести

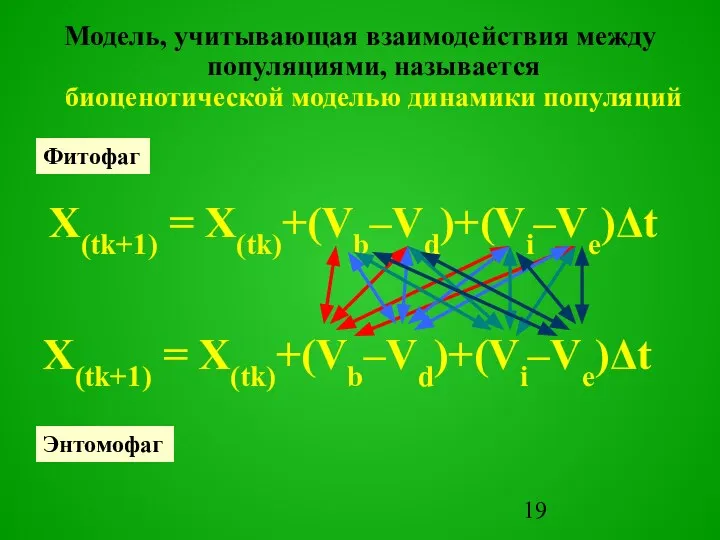
Слайд 19Модель, учитывающая взаимодействия между популяциями, называется
биоценотической моделью динамики популяций
X(tk+1) = X(tk)+(Vb–Vd)+(Vi–Ve)Δt
Фитофаг
Энтомофаг
X(tk+1) =
Модель, учитывающая взаимодействия между популяциями, называется
биоценотической моделью динамики популяций
X(tk+1) = X(tk)+(Vb–Vd)+(Vi–Ve)Δt
Фитофаг
Энтомофаг
X(tk+1) =
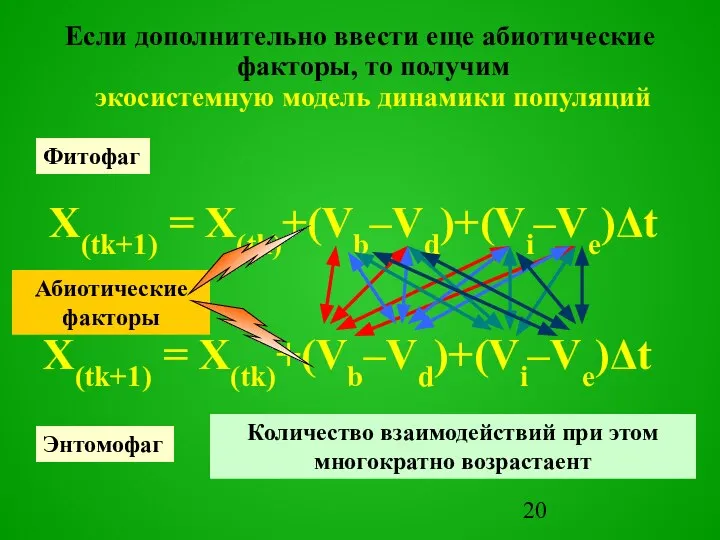
Слайд 20Если дополнительно ввести еще абиотические факторы, то получим
экосистемную модель динамики популяций
X(tk+1)
Если дополнительно ввести еще абиотические факторы, то получим
экосистемную модель динамики популяций
X(tk+1)
Слайд 21Рождаемость (Vb) и смертность (Vd) зависят от многих факторов, основными из которых
Рождаемость (Vb) и смертность (Vd) зависят от многих факторов, основными из которых

Слайд 22Частично преодолеть эти трудности оказалось возможным, когда взаимодействия стали классифицировать не по
Частично преодолеть эти трудности оказалось возможным, когда взаимодействия стали классифицировать не по

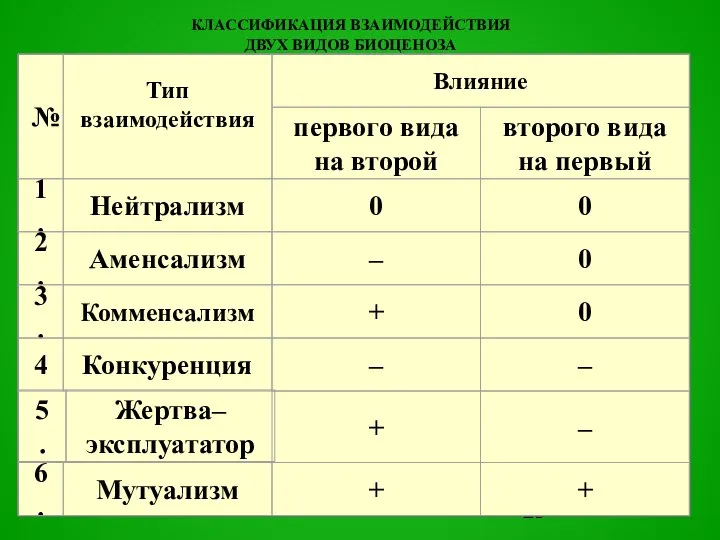
Слайд 23КЛАССИФИКАЦИЯ ВЗАИМОДЕЙСТВИЯ
ДВУХ ВИДОВ БИОЦЕНОЗА
КЛАССИФИКАЦИЯ ВЗАИМОДЕЙСТВИЯ
ДВУХ ВИДОВ БИОЦЕНОЗА
Слайд 24Популяции двух видов
не оказывают непосредственного воздействия друг на друга.
Нейтрализм (0, 0)
0
0
Такой тип
Популяции двух видов
не оказывают непосредственного воздействия друг на друга.
Нейтрализм (0, 0)
0
0
Такой тип

Слайд 25один из видов взаимодействующей пары в биоценозе оказывает отрицательное воздействие на рост
один из видов взаимодействующей пары в биоценозе оказывает отрицательное воздействие на рост

Слайд 26При слабом ингибировании система стремится к устойчивому стационарному состоянию, в котором численность
При слабом ингибировании система стремится к устойчивому стационарному состоянию, в котором численность

Слайд 27Рис. 86. Динамика плотности популяции аменсала при слабом ингибировании первым видом
Аменсализм (-,
Рис. 86. Динамика плотности популяции аменсала при слабом ингибировании первым видом
Аменсализм (-,
Слайд 28При сильном ингибировании популяция аменсала полностью погибает (элиминируется)
Аменсализм (-, 0) сильное
При сильном ингибировании популяция аменсала полностью погибает (элиминируется)
Аменсализм (-, 0) сильное

Слайд 29Рис. 85. Динамика плотности популяции аменсала при сильном ингибировании первым видом
Аменсализм (-,
Рис. 85. Динамика плотности популяции аменсала при сильном ингибировании первым видом
Аменсализм (-,
Слайд 30первый вид, называемый «хозяином», положительно воздействует на второй, называемый «комменсалом» («нахлебником»), а
первый вид, называемый «хозяином», положительно воздействует на второй, называемый «комменсалом» («нахлебником»), а

Слайд 31Рис. 87. Динамика плотности популяции при комменсализме
Комменсализм (+, 0)
Рис. 87. Динамика плотности популяции при комменсализме
Комменсализм (+, 0)
Слайд 32любое взаимно отрицательное отношение между видами.
Конкуренция (интерференция (-, -)
-
-
вид, получающий какое-либо преимущество
любое взаимно отрицательное отношение между видами.
Конкуренция (интерференция (-, -)
-
-
вид, получающий какое-либо преимущество

Слайд 33Частными случаями конкуренции являются:
– конкуренция за тот или иной ограниченный ресурс (соперничество).
Частными случаями конкуренции являются:
– конкуренция за тот или иной ограниченный ресурс (соперничество).

Слайд 34Результаты конкуренции
Первый вид, как более сильный конкурент, всегда будет вытеснять второй, независимо
Результаты конкуренции
Первый вид, как более сильный конкурент, всегда будет вытеснять второй, независимо
Слайд 35Результаты конкуренции
Второй вид, будучи более сильным конкурентом, вытесняет первый независимо от начальных
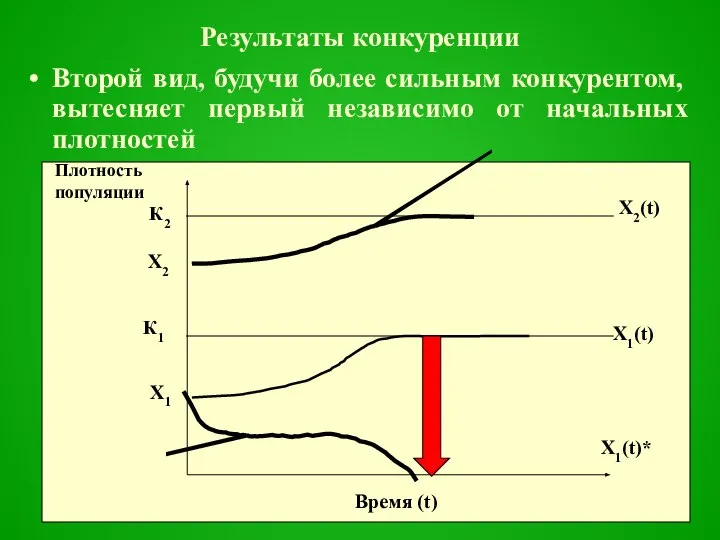
Результаты конкуренции
Второй вид, будучи более сильным конкурентом, вытесняет первый независимо от начальных
Слайд 36Результаты конкуренции
Если виды одинаково конкурентноспособны, то независимо от исходных плотностей система приходит
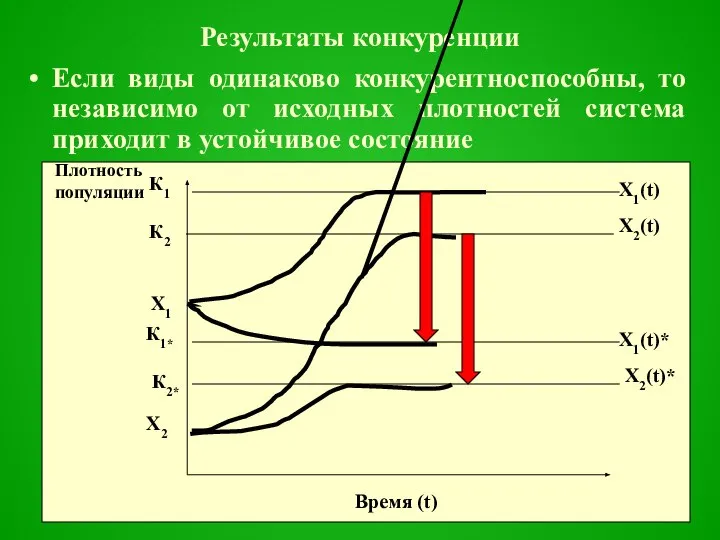
Результаты конкуренции
Если виды одинаково конкурентноспособны, то независимо от исходных плотностей система приходит
Слайд 37Результаты конкуренции
Если емкости внешней среды не лимитируют рост плотности популяций видов, то
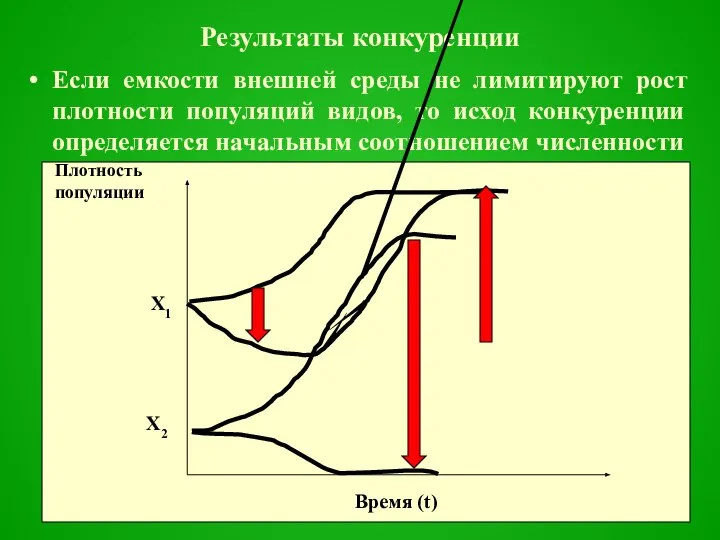
Результаты конкуренции
Если емкости внешней среды не лимитируют рост плотности популяций видов, то
Слайд 38Таким образом, для обеспечения устойчивости конкурентного равновесия в системе, если оно возможно,
Таким образом, для обеспечения устойчивости конкурентного равновесия в системе, если оно возможно,

Слайд 39отношения между двумя видами, при которых увеличение (или уменьшение) плотности популяции первого
отношения между двумя видами, при которых увеличение (или уменьшение) плотности популяции первого

Слайд 40«Жертва-эксплуататор» (+, -)
Вид-эксплуататор не способен прокормиться за счет данной жертвы и со
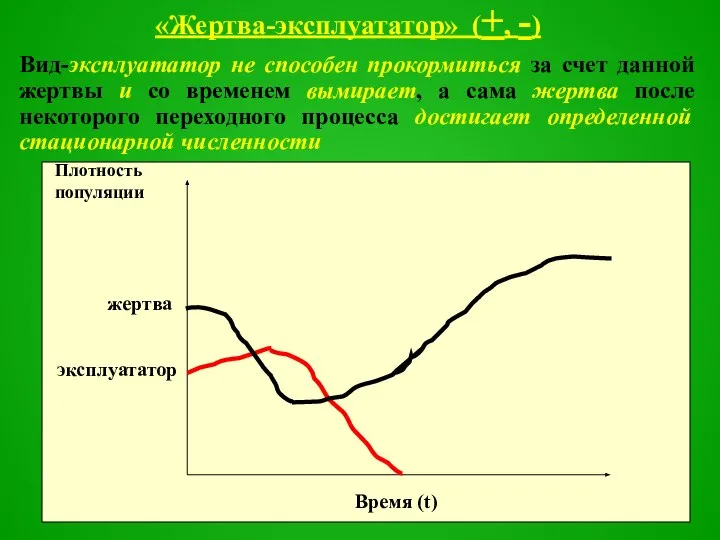
«Жертва-эксплуататор» (+, -)
Вид-эксплуататор не способен прокормиться за счет данной жертвы и со
Слайд 41Типичный пример – применение биологического метода защиты растений, когда внедренный в экосистему
Типичный пример – применение биологического метода защиты растений, когда внедренный в экосистему

Слайд 42«Жертва-эксплуататор» (+, -)
Вид-эксплуататор слишком эффективен в поисках жертвы и быстро размножается, в
«Жертва-эксплуататор» (+, -)
Вид-эксплуататор слишком эффективен в поисках жертвы и быстро размножается, в
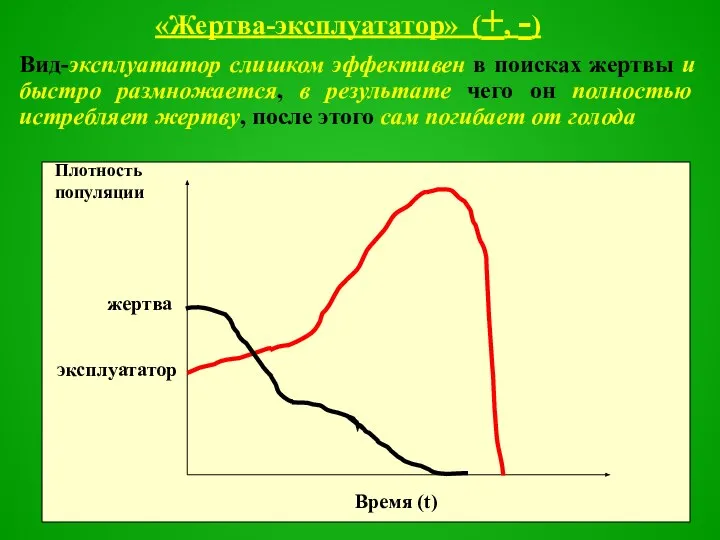
Слайд 43Ситуация “козел в огороде” характерна для монофагов, используемых как средство биологической защиты
Ситуация “козел в огороде” характерна для монофагов, используемых как средство биологической защиты

Слайд 44«Жертва-эксплуататор» (+, -)
Существует единственная устойчивая комбинация плотностей жертвы и эксплуататора, и из
«Жертва-эксплуататор» (+, -)
Существует единственная устойчивая комбинация плотностей жертвы и эксплуататора, и из
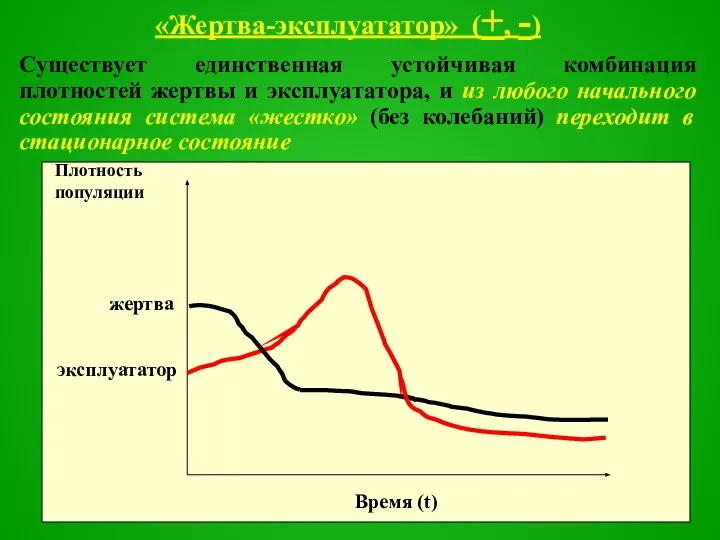
Слайд 45Это ситуация, когда эксплуататор достаточно эффективен и может быстро снижать численность жертвы
Это ситуация, когда эксплуататор достаточно эффективен и может быстро снижать численность жертвы

Слайд 46«Жертва-эксплуататор» (+, -)
Существует единственная устойчивая комбинация плотностей популяций жертвы и эксплуататора, при
«Жертва-эксплуататор» (+, -)
Существует единственная устойчивая комбинация плотностей популяций жертвы и эксплуататора, при
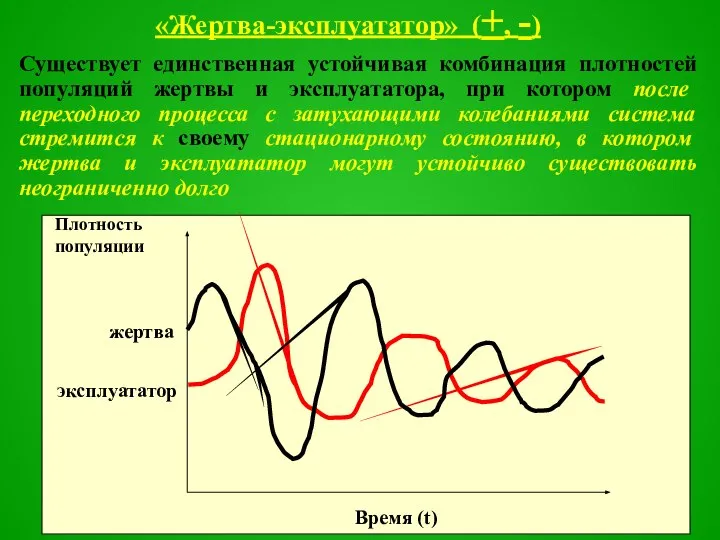
Слайд 47Например, взаимодействие растений и копытных растительноядных животных, которые периодически мигрируют на другие
Например, взаимодействие растений и копытных растительноядных животных, которые периодически мигрируют на другие

Слайд 48«Жертва-эксплуататор» (+, -)
Из любого начального состояния траектория системы стремится к единственно замкнутой
«Жертва-эксплуататор» (+, -)
Из любого начального состояния траектория системы стремится к единственно замкнутой
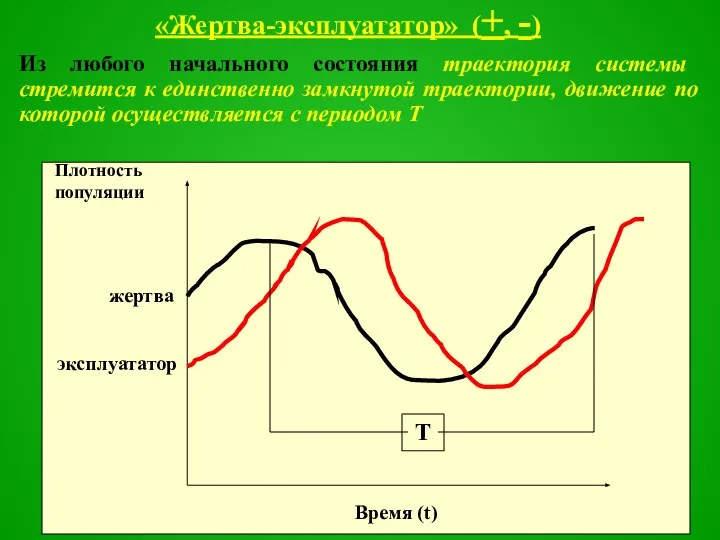
Слайд 49Такие автоколебания характерны для ненарушенных внешними факторами систем, где высокая степень саморегуляции.
Синусоидальная
Синусоидальная

Слайд 50взаимно положительное влияние, которое в конечном счете проявляется в том, что увеличение
взаимно положительное влияние, которое в конечном счете проявляется в том, что увеличение

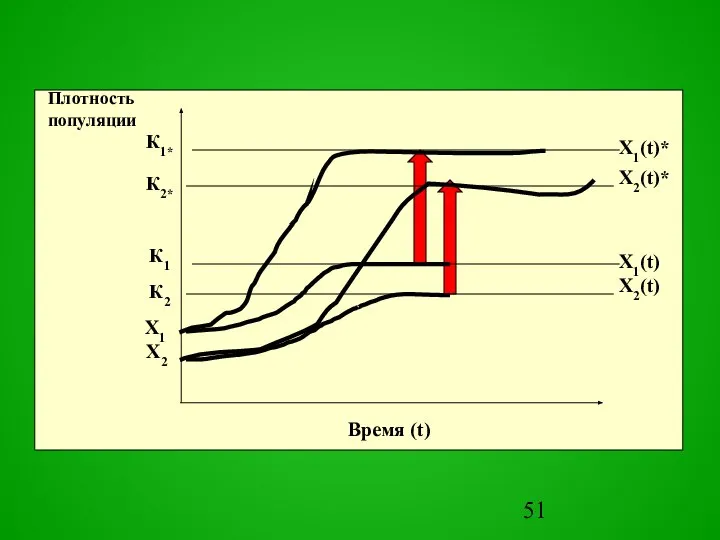
Слайд 53Все многообразие моделей прогнозов по шкале времени можно разделить на прогнозы
МОДЕЛЬ
Все многообразие моделей прогнозов по шкале времени можно разделить на прогнозы
МОДЕЛЬ

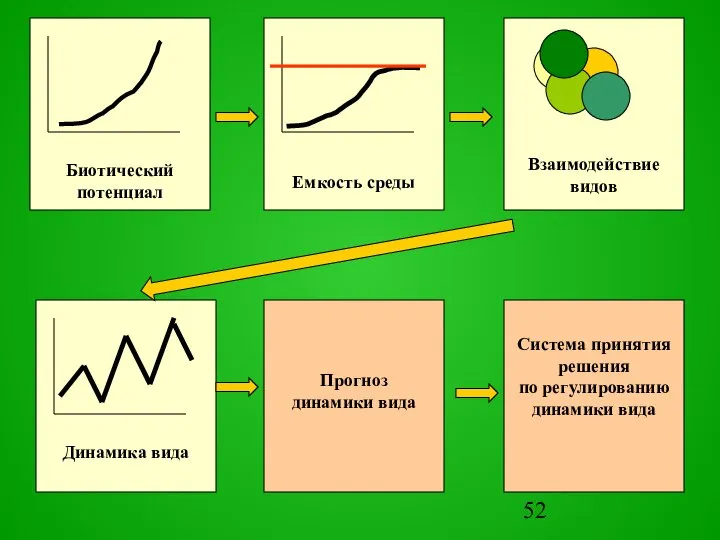
Слайд 54Методы прогнозов
Методы прогнозов
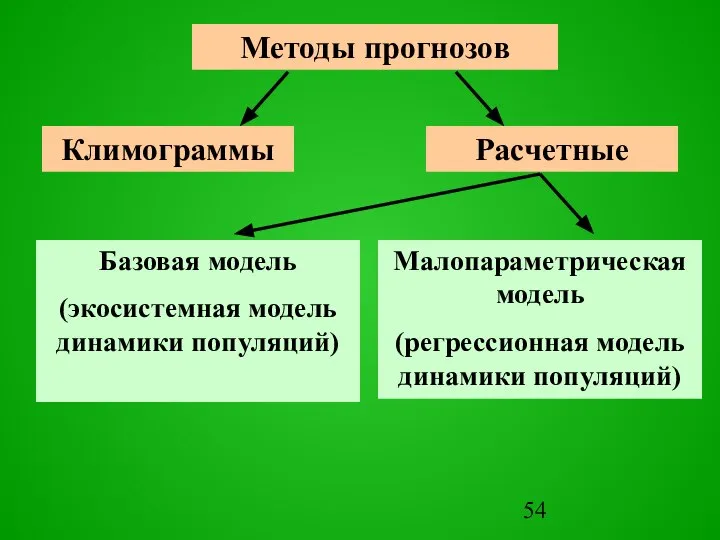
Слайд 55Климограмма
это двумерная экологическая ниша, где учитываются температура воздуха
Климограмма
это двумерная экологическая ниша, где учитываются температура воздуха
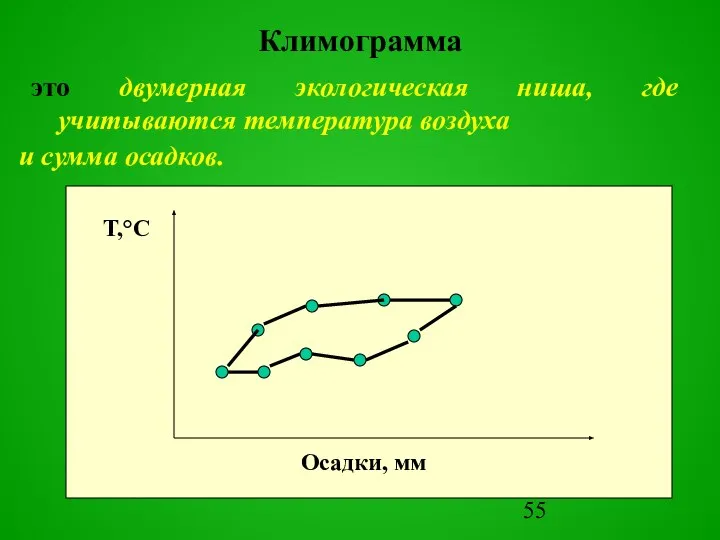
Слайд 56Климограмма
Климограмма
Слайд 57При помощи климограммы можно установить
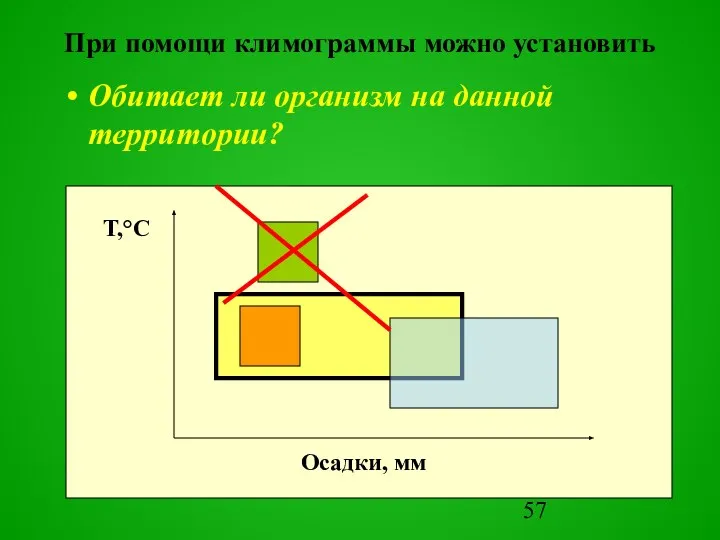
Обитает ли организм на данной территории?
Осадки, мм
При помощи климограммы можно установить
Обитает ли организм на данной территории?
Осадки, мм
Слайд 58При помощи климограммы можно установить
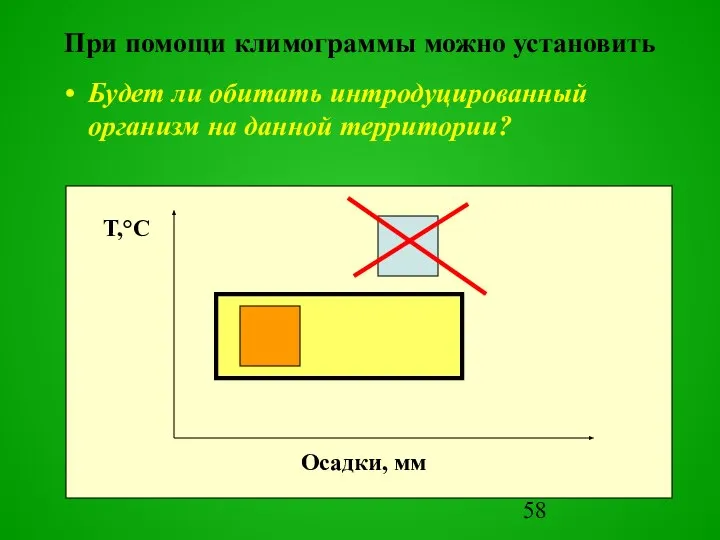
Будет ли обитать интродуцированный организм на данной территории?
Осадки,
При помощи климограммы можно установить
Будет ли обитать интродуцированный организм на данной территории?
Осадки,
Слайд 59При помощи климограммы можно установить
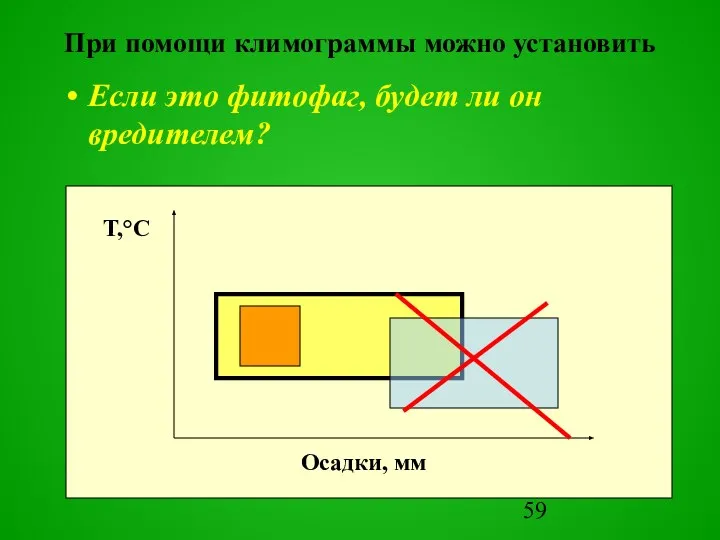
Если это фитофаг, будет ли он вредителем?
Осадки, мм
При помощи климограммы можно установить
Если это фитофаг, будет ли он вредителем?
Осадки, мм
Слайд 60При помощи климограммы можно установить
Будет ли массовое размножение организма?
Осадки, мм
При помощи климограммы можно установить
Будет ли массовое размножение организма?
Осадки, мм
Слайд 61Например, размножение тлей зависит от метеорологических условий и фазы разввития растения.
Низкие температуры
Например, размножение тлей зависит от метеорологических условий и фазы разввития растения.
Низкие температуры

Слайд 62В 1988 году в Костромской области наблюдалось массовое размножение тли.
Воспользовавшись этим
В 1988 году в Костромской области наблюдалось массовое размножение тли.
Воспользовавшись этим

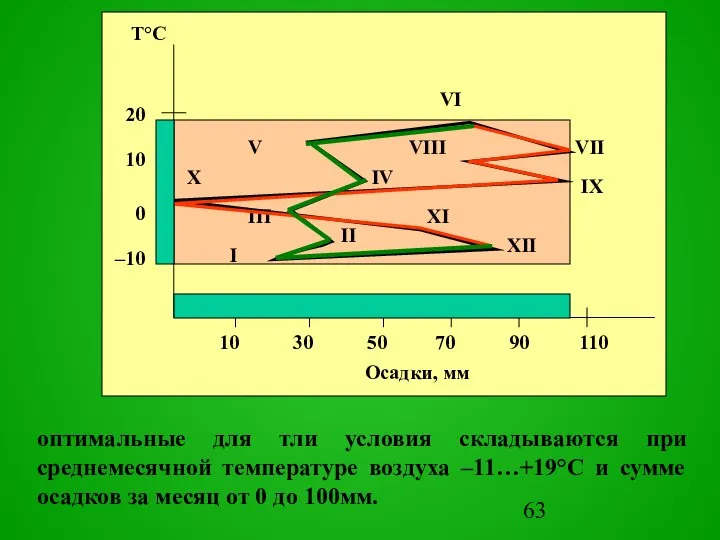
Слайд 63оптимальные для тли условия складываются при среднемесячной температуре воздуха –11…+19°С и сумме
оптимальные для тли условия складываются при среднемесячной температуре воздуха –11…+19°С и сумме
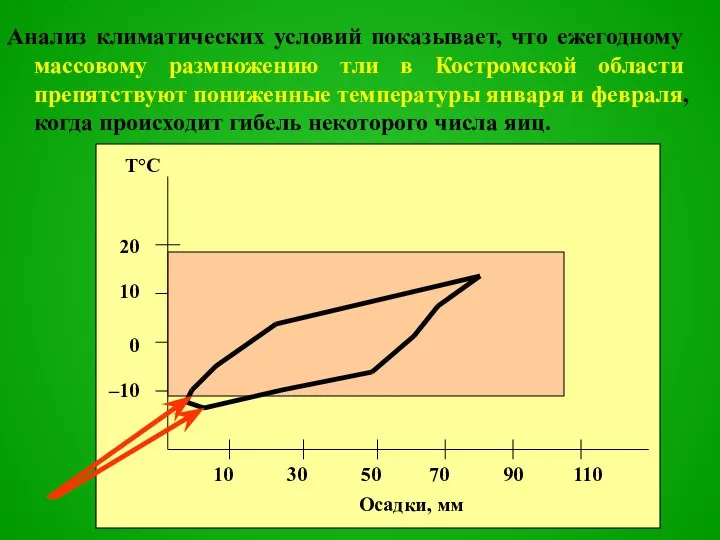
Слайд 64Анализ климатических условий показывает, что ежегодному массовому размножению тли в Костромской области
Анализ климатических условий показывает, что ежегодному массовому размножению тли в Костромской области
Слайд 65Методы прогнозов
Методы прогнозов
Слайд 66Сезонный и оперативный прогнозы динамики составляются при помощи математических функций.
Y=F(x)
Для обеспечения возможности
Сезонный и оперативный прогнозы динамики составляются при помощи математических функций.
Y=F(x)
Для обеспечения возможности

Слайд 67По разработанным моделям динамики популяций рассчитывается прогноз численности фитофагов и энтомофагов,
количество
Суммарная энтальпия
По разработанным моделям динамики популяций рассчитывается прогноз численности фитофагов и энтомофагов,
количество
Суммарная энтальпия
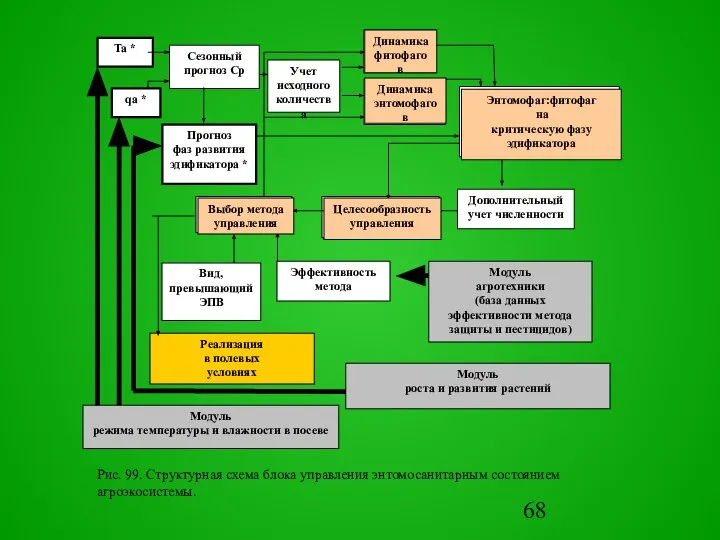
Слайд 68Ta *
qa *
Рис. 99. Структурная схема блока управления энтомосанитарным состоянием агроэкосистемы.
Энтомофаг:фитофаг
на
критическую
Ta *
qa *
Рис. 99. Структурная схема блока управления энтомосанитарным состоянием агроэкосистемы.
Энтомофаг:фитофаг на критическую
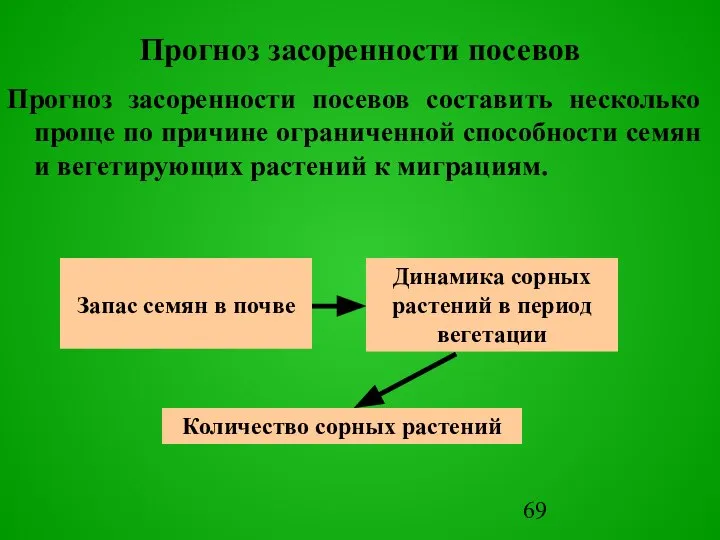
Слайд 69Прогноз засоренности посевов
Прогноз засоренности посевов составить несколько проще по причине ограниченной способности
Прогноз засоренности посевов
Прогноз засоренности посевов составить несколько проще по причине ограниченной способности
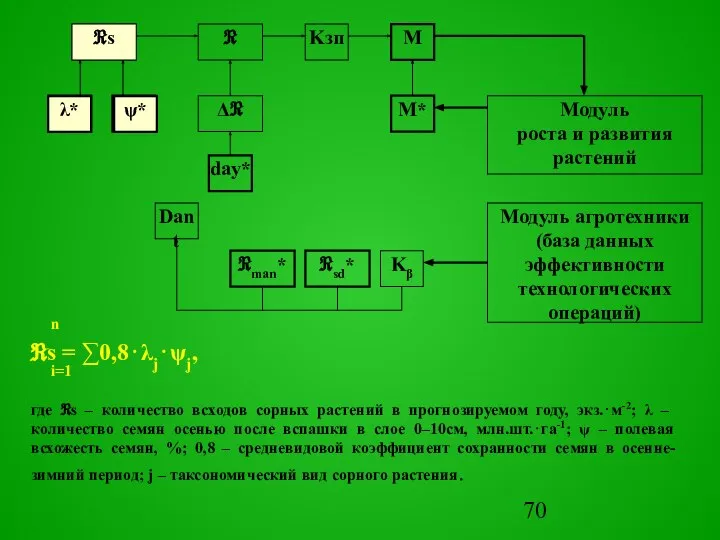
Слайд 70М*
М
day*
ψ*
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
Dant
Kβ
λ*
Δℜ
ℜsd*
ℜman*
ℜ
ℜs
n
ℜs = ∑0,8⋅λj⋅ψj,
М*
М
day*
ψ*
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
Dant
Kβ
λ*
Δℜ
ℜsd*
ℜman*
ℜ
ℜs
n
ℜs = ∑0,8⋅λj⋅ψj,
Слайд 71М*
М
day*
ψ*
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
Dant
Kβ
λ*
Δℜ
ℜsd*
ℜman*
ℜ
ℜs
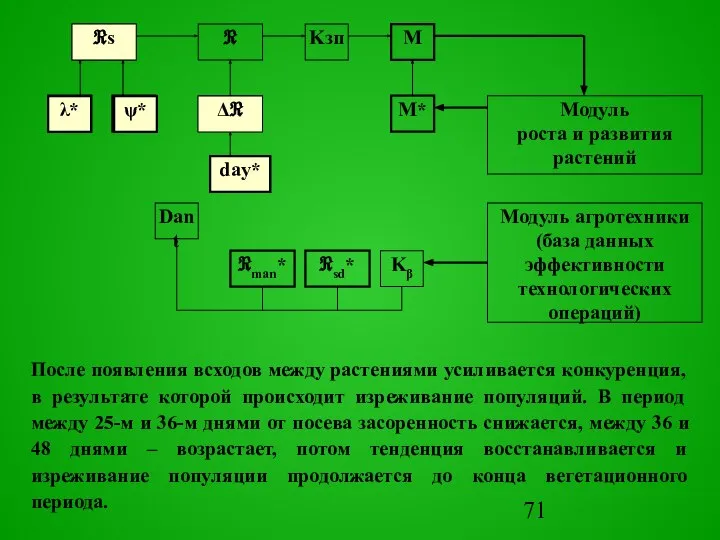
После появления всходов между
М*
М
day*
ψ*
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
Dant
Kβ
λ*
Δℜ
ℜsd*
ℜman*
ℜ
ℜs
После появления всходов между
Слайд 72М*
М
day*
ψ*
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
Dant
Kβ
λ*
Δℜ
ℜsd*
ℜman*
ℜ
ℜs
λ*
ψ*
ℜs
day*
Δℜ
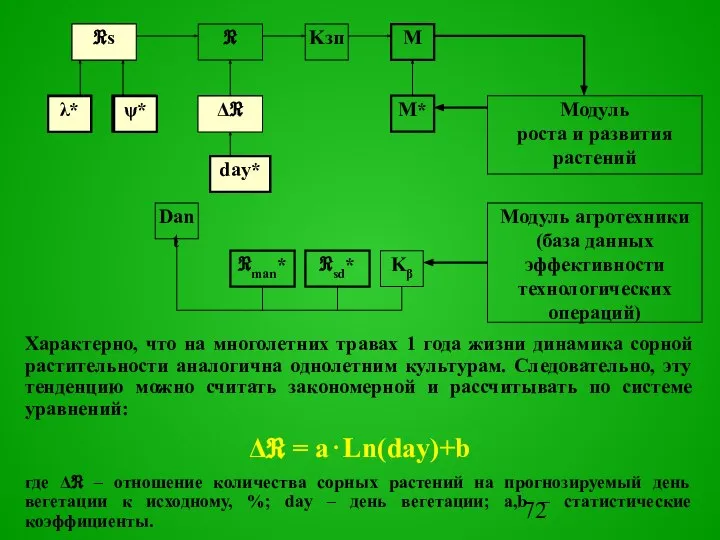
Характерно, что на многолетних
М*
М
day*
ψ*
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
Dant
Kβ
λ*
Δℜ
ℜsd*
ℜman*
ℜ
ℜs
λ*
ψ*
ℜs
day*
Δℜ
Характерно, что на многолетних
Слайд 73М*
М
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
Dant
Kβ
ℜsd*
ℜman*
ℜ
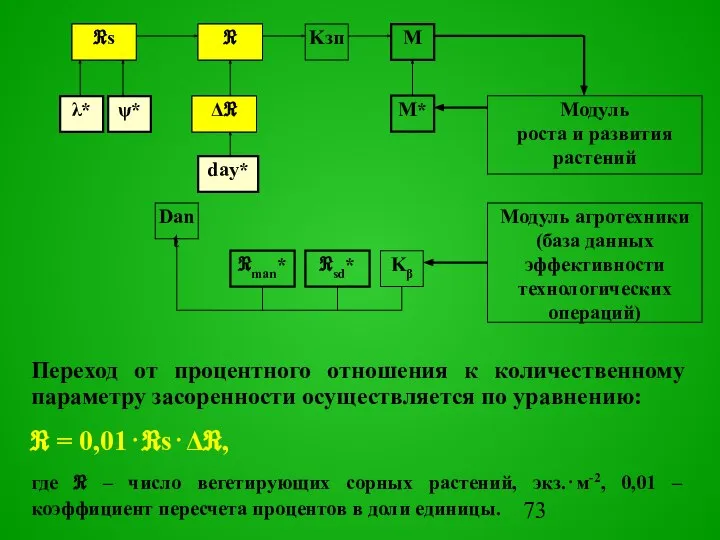
Переход от процентного отношения
М*
М
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
Dant
Kβ
ℜsd*
ℜman*
ℜ
Переход от процентного отношения
Слайд 74М*
М
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
Dant
Kβ
ℜsd*
ℜman*
λ*
ψ*
day*
ℜ
ℜs
Δℜ
М*
М
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
Dant
Kβ
ℜsd*
ℜman*
λ*
ψ*
day*
ℜ
ℜs
Δℜ
Слайд 7520%
30%
80%
50%
20%
30%
80%
50%

Слайд 76М*
М
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
Dant
Kβ
ℜsd*
ℜman*
λ*
ψ*
day*
ℜ
ℜs
Δℜ
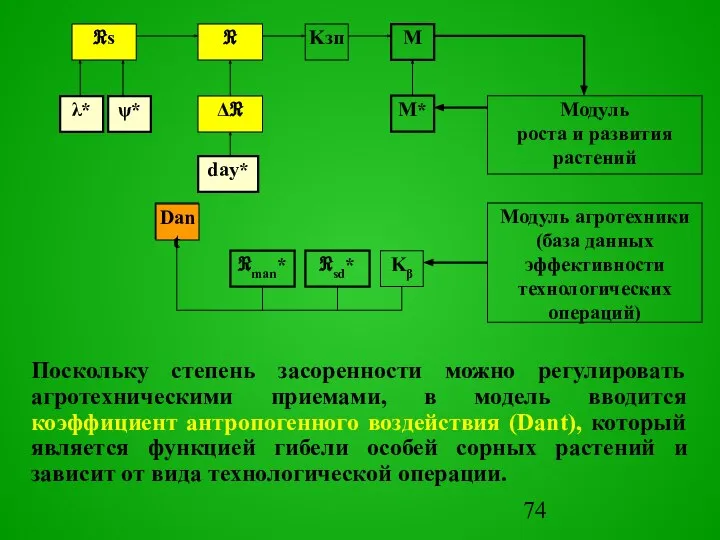
Учитывая поступление семян извне
М*
М
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
Dant
Kβ
ℜsd*
ℜman*
λ*
ψ*
day*
ℜ
ℜs
Δℜ
Учитывая поступление семян извне
Слайд 77М*
М
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
λ*
ψ*
day*
ℜ
ℜs
Δℜ
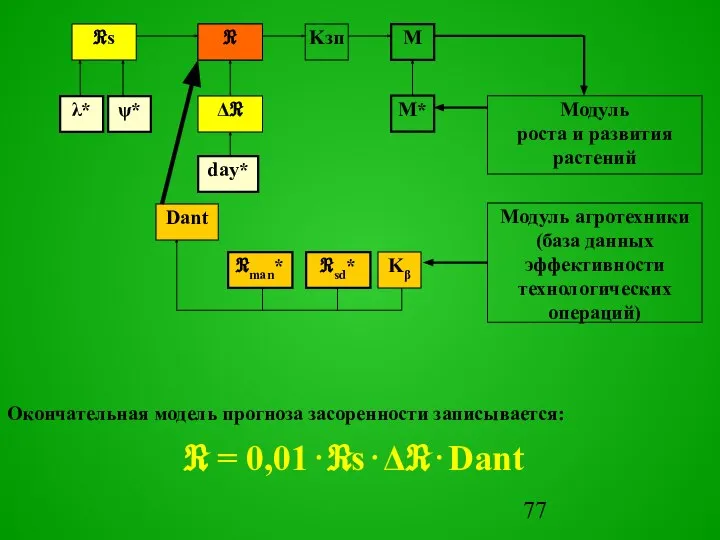
Окончательная модель прогноза засоренности
М*
М
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
λ*
ψ*
day*
ℜ
ℜs
Δℜ
Окончательная модель прогноза засоренности
Слайд 78М*
М
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
λ*
ψ*
day*
ℜ
ℜs
Δℜ
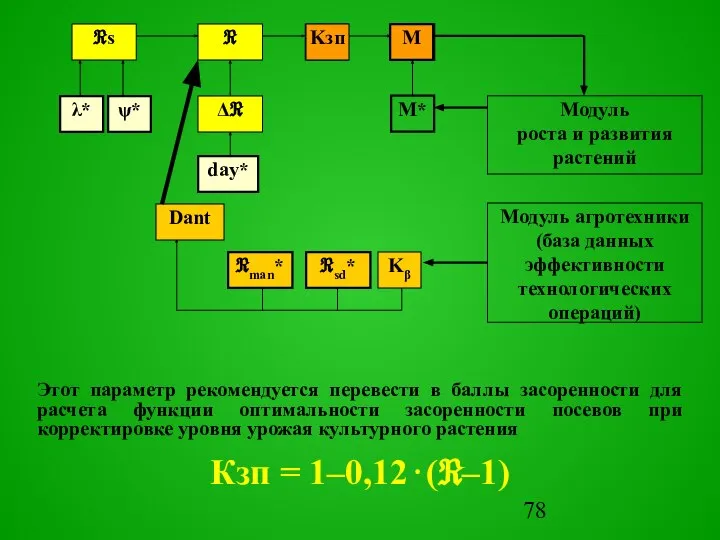
Этот параметр рекомендуется перевести
М*
М
Kзп
Модуль агротехники
(база данных эффективности технологических операций)
Модуль
роста и развития растений
λ*
ψ*
day*
ℜ
ℜs
Δℜ
Этот параметр рекомендуется перевести
Слайд 79Тогда малопараметрическая модель прогноза урожайности с учетом засоренности посевов запишется в виде
Y(Q,W,t,NPK,pH,Зп)
Тогда малопараметрическая модель прогноза урожайности с учетом засоренности посевов запишется в виде
Y(Q,W,t,NPK,pH,Зп)

 Экологический клуб Экос
Экологический клуб Экос Летнее чтение. Лицом к лицу с природой
Летнее чтение. Лицом к лицу с природой Мусорное дело
Мусорное дело Роль промышленных предприятий в загрязнении атмосферы
Роль промышленных предприятий в загрязнении атмосферы Уровень комфортности классной комнаты и наше самочувствие
Уровень комфортности классной комнаты и наше самочувствие Важнейшие экологические факторы в водной среде
Важнейшие экологические факторы в водной среде Научно-исследовательский интегрированный проект по экологии и химии
Научно-исследовательский интегрированный проект по экологии и химии Берегите природу
Берегите природу Берегите природу
Берегите природу Основные параметры и понятия экологического мониторинга. Понятие об экологической опасности
Основные параметры и понятия экологического мониторинга. Понятие об экологической опасности Экологическая характеристика популяции
Экологическая характеристика популяции Природа Крыма
Природа Крыма Цените красоту природы
Цените красоту природы Як ми наносимо шкоду власній планеті
Як ми наносимо шкоду власній планеті Мероприятия по повышению эффективности работы предприятия при осуществлении уборки города, предприятие СпецАвтоТранс
Мероприятия по повышению эффективности работы предприятия при осуществлении уборки города, предприятие СпецАвтоТранс Природа в городе. Угадай растения
Природа в городе. Угадай растения Чрезвычайные ситуации экологического характера
Чрезвычайные ситуации экологического характера Экологическое право. Человек в природе
Экологическое право. Человек в природе ресурсосбережение.
ресурсосбережение. Зеленые глаза. Итоги работы с экочемоданчиком
Зеленые глаза. Итоги работы с экочемоданчиком Экологический мониторинг окружающей среды с. Мултаново
Экологический мониторинг окружающей среды с. Мултаново Экологическая деятельность ПАО “Мосэнерго”
Экологическая деятельность ПАО “Мосэнерго” Петро био. Устойчивые энергоэффективные решения
Петро био. Устойчивые энергоэффективные решения Анализ ТБО в Санкт-петербурге
Анализ ТБО в Санкт-петербурге Plog-марафон
Plog-марафон Есть ли в России возможность использовать альтернативную энергетику?
Есть ли в России возможность использовать альтернативную энергетику? Загрязнение Мирового океана Аникеенко Артемий 11 А
Загрязнение Мирового океана Аникеенко Артемий 11 А Экологическое сообщество Муравейник
Экологическое сообщество Муравейник