- Физиология возбудимых клеток. Мембранный потенциал

Содержание
- 2. Типы возбудимых клеток Нейроны Мышечные клетки Секреторные клетки Рецепторные клетки
- 3. Строение животной клетки
- 4. Особенности строения нейрона
- 5. Виды нейронов А — веретенообразный (кишечнополостные); Б — псевдоуниполярный (сенсорный нейрон позвоночных); В — мультиполярный (позвоночные);
- 6. Формирование трансмембранного потенциала А. в чашке Петри KCl K+ Cl- Градиент концентрации Градиент заряда равновесие
- 7. Рассчет заряда на мембране Равновесный потенциал для какого-либо иона Х можно рассчитать из уравнения, полученного в
- 8. Б. мицелла – синтетический прообраз клетки
- 9. Мембрана живой клетки К+ Na+ Са++
- 10. Равновесные потенциалы(Е) Движущая сила (V- Е) Cl- -89 - 47 Cl-каналы
- 11. Мембрана живой клетки полупроницаема -61 К+ Na+ = 0,023 рК Са++ рСа++ = 0 Cl-
- 12. Проницаемость обеспечена ионные каналами мембраны 1-1000 каналов на квадратный микрометр мембраны Центральная водная пора Устья канала:
- 13. Создание градиента концентрации: 1. Na-K АТФ-аза 2. ионные обменники Транспорт 3 Na/2K за счет энергии 1
- 14. Изменения мембранного потенциала покоя 1. Деполяризация- уменьшение (ее скорость определяется постоянной времени (τm=RmCm)) 2. Гиперполяризация- увеличение
- 15. Внутриклеточная регистрация мембранного потенциала покоя Внутриклеточная микроэлектродная регистрация Величина МПП в возбудимых клетках – от -60
- 16. Потенциал действия Фаза деполяризации Фаза реполяризации Раздражающий импульс
- 17. Временной ход ионных токов во время потенциала действия
- 18. Фармакологическое разделение ионных токов ядами контроль Калиевый ток Натриевый ток Выводы Входящий ток переносится ионами натрия,
- 19. Фазы потенциала действия 1- порог (около 50 мв, ток Na>K) 2- деполяризация 0,5 мс (вход Na)
- 20. Вызывается сверхпороговым раздражением Амплитуда не зависит от силы раздражения Распространяется по всей мембране не затухая Связан
- 21. Исследование отдельного канала Возможность исследовать отдельный канал Возможность менять потенциал на мембране Возможность менять ионный состав
- 22. Нобелевская премия 1991 года в области физиологии и медицины Эрвин Нейер и Берт Сакманн «за открытия
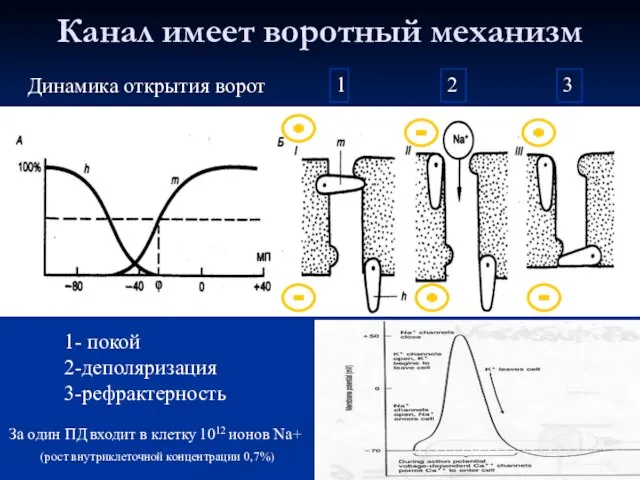
- 23. Канал имеет воротный механизм 1- покой 2-деполяризация 3-рефрактерность Динамика открытия ворот 1 2 3 За один
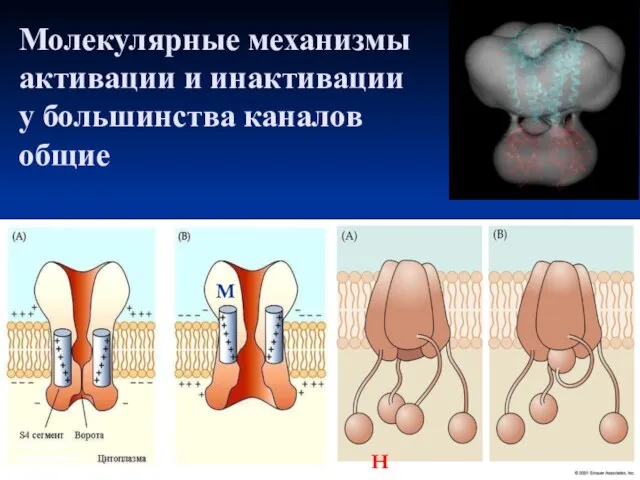
- 24. Молекулярные механизмы активации и инактивации у большинства каналов общие H M
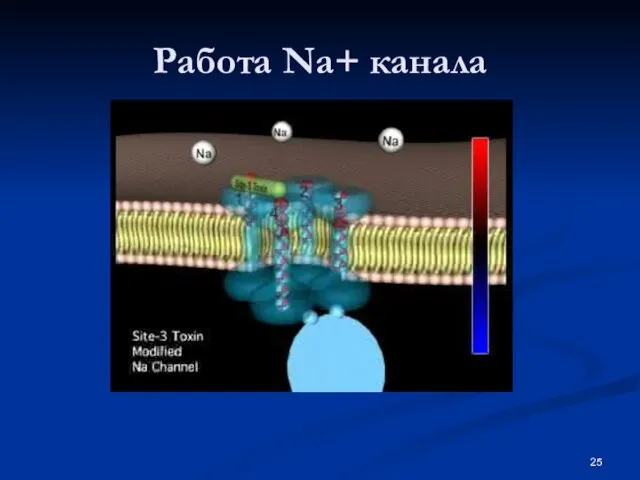
- 25. Работа Na+ канала
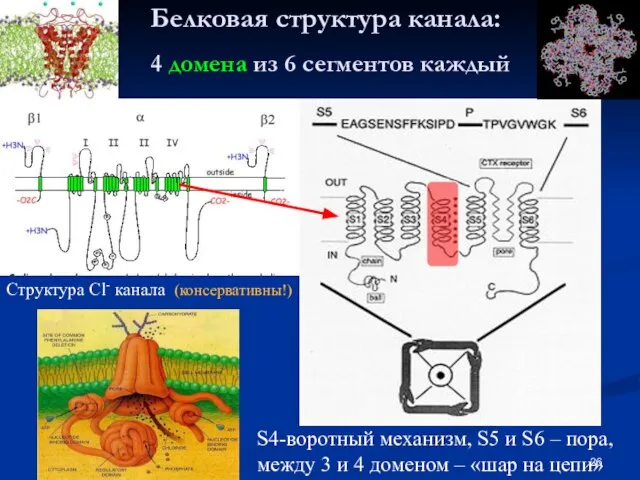
- 26. Белковая структура канала: 4 домена из 6 сегментов каждый Структура Cl- канала (консервативны!) S4-воротный механизм, S5
- 27. Рефрактерность - снижение способности клетки отвечать на раздражение в результате временной инактивации натриевых каналов Абсолютная рефрактерность
- 28. Распространение потенциала действия по волокну Увеличение диаметра волокна повышает скорость проведения: Постоянная длины волокна (от 0,1
- 29. Миелинизированные волокна Эстафетный (до 40 м/с) и сальтаторный (до 120м/с) механизмы распространения возбуждения
- 30. Скорость проведения ПД по разным типам волокон
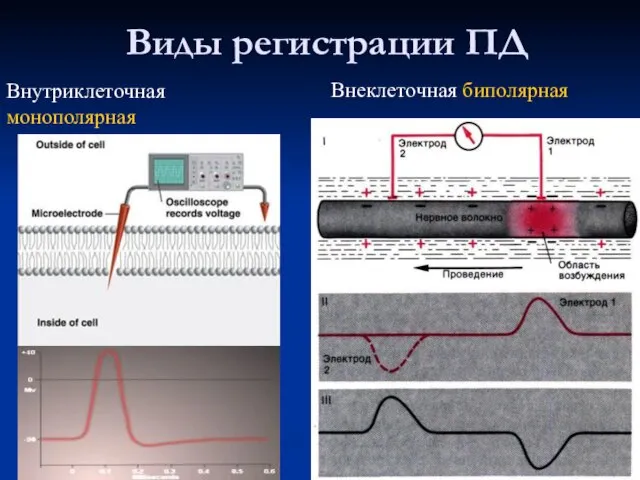
- 31. Виды регистрации ПД Внутриклеточная монополярная Внеклеточная биполярная
- 33. Скачать презентацию
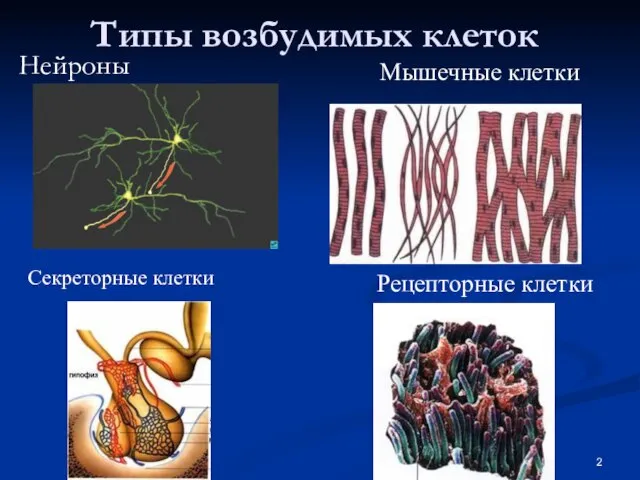
Слайд 3Строение животной клетки
Строение животной клетки
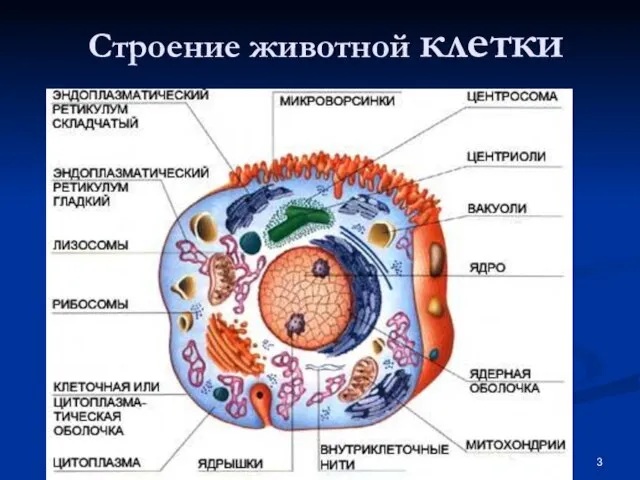
Слайд 4Особенности строения нейрона
Особенности строения нейрона
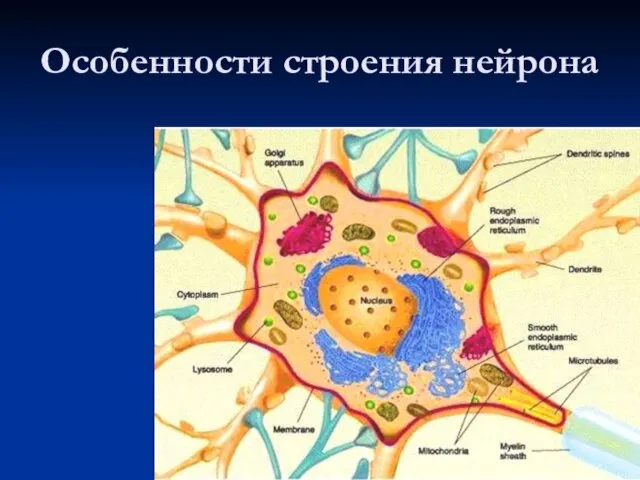
Слайд 5Виды нейронов
А — веретенообразный (кишечнополостные);
Б — псевдоуниполярный (сенсорный нейрон позвоночных);
В —
Виды нейронов
А — веретенообразный (кишечнополостные);
Б — псевдоуниполярный (сенсорный нейрон позвоночных);
В —
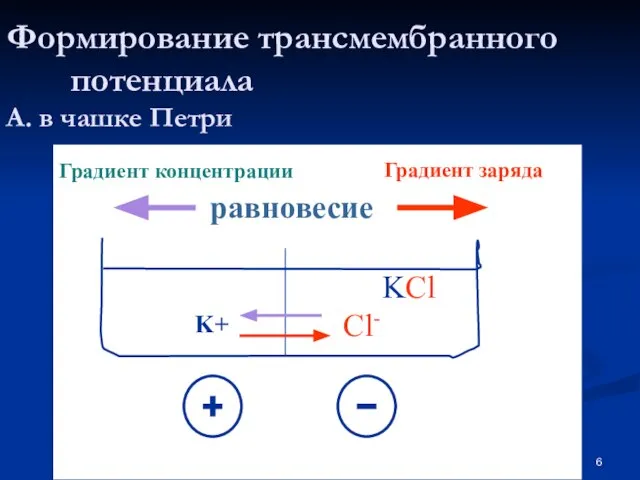
Слайд 6Формирование трансмембранного потенциала
А. в чашке Петри
KCl
K+
Cl-
Градиент концентрации
Градиент заряда
равновесие
Формирование трансмембранного потенциала
А. в чашке Петри
KCl
K+
Cl-
Градиент концентрации
Градиент заряда
равновесие
Слайд 7Рассчет заряда на мембране
Равновесный потенциал для какого-либо иона Х можно рассчитать из
Рассчет заряда на мембране
Равновесный потенциал для какого-либо иона Х можно рассчитать из

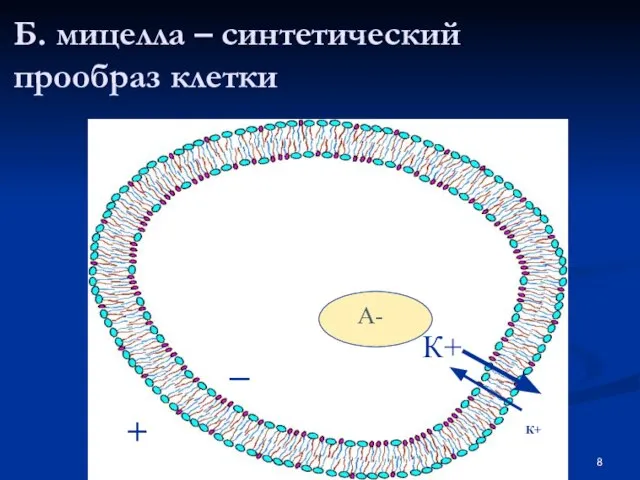
Слайд 8Б. мицелла – синтетический прообраз клетки
Б. мицелла – синтетический прообраз клетки
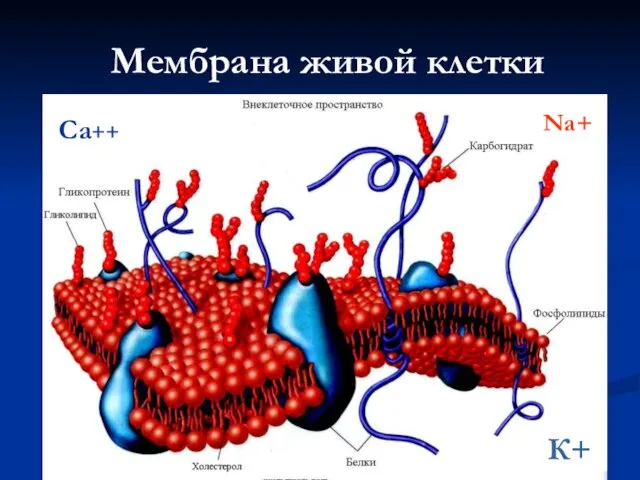
Слайд 9Мембрана живой клетки
К+
Na+
Са++
Мембрана живой клетки
К+
Na+
Са++
Слайд 10Равновесные потенциалы(Е) Движущая сила (V- Е)
Cl-
-89 - 47
Cl-каналы
Равновесные потенциалы(Е) Движущая сила (V- Е)
Cl-
-89 - 47
Cl-каналы

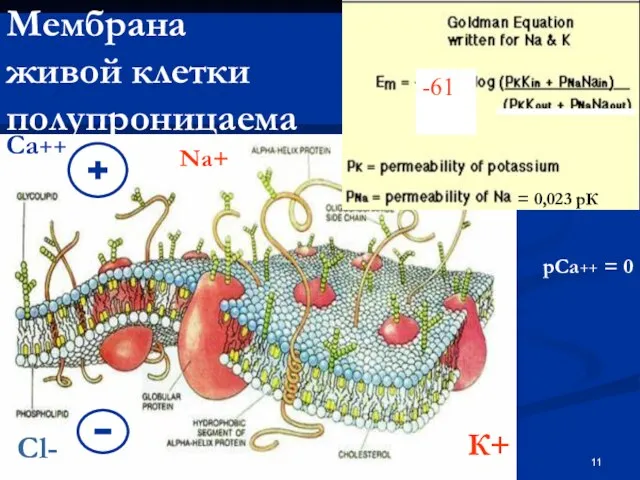
Слайд 11Мембрана
живой клетки полупроницаема
-61
К+
Na+
= 0,023 рК
Са++
рСа++ = 0
Cl-
Мембрана
живой клетки полупроницаема
-61
К+
Na+
= 0,023 рК
Са++
рСа++ = 0
Cl-
Слайд 12Проницаемость обеспечена ионные каналами мембраны
1-1000 каналов на квадратный микрометр мембраны
Центральная водная пора
Устья
Проницаемость обеспечена ионные каналами мембраны
1-1000 каналов на квадратный микрометр мембраны
Центральная водная пора
Устья

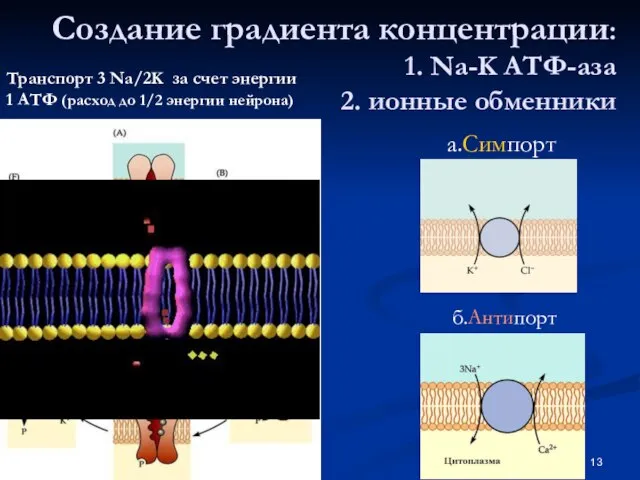
Слайд 13Создание градиента концентрации:
1. Na-K АТФ-аза
2. ионные обменники
Транспорт 3 Na/2K за
Создание градиента концентрации:
1. Na-K АТФ-аза
2. ионные обменники
Транспорт 3 Na/2K за
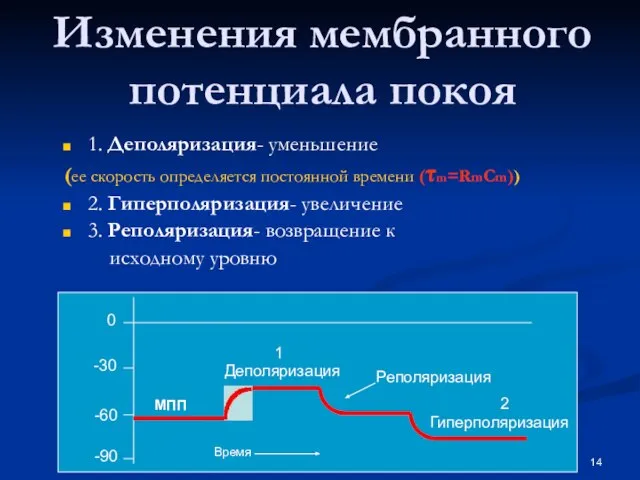
Слайд 14Изменения мембранного потенциала покоя
1. Деполяризация- уменьшение
(ее скорость определяется постоянной времени (τm=RmCm))
2.
Изменения мембранного потенциала покоя
1. Деполяризация- уменьшение
(ее скорость определяется постоянной времени (τm=RmCm))
2.
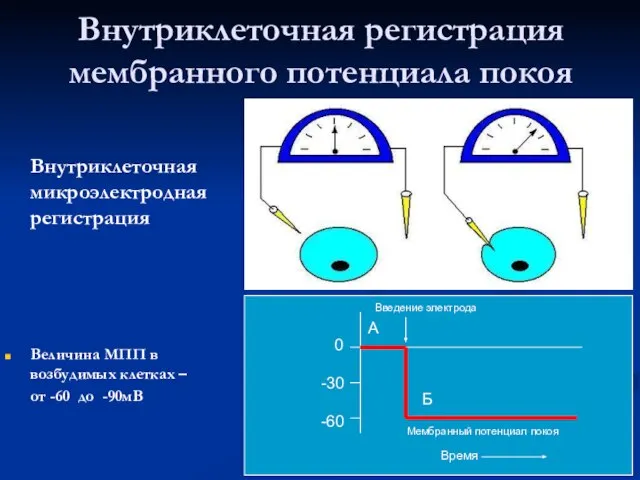
Слайд 15Внутриклеточная регистрация мембранного потенциала покоя
Внутриклеточная микроэлектродная регистрация
Величина МПП в возбудимых клетках
Внутриклеточная регистрация мембранного потенциала покоя
Внутриклеточная микроэлектродная регистрация
Величина МПП в возбудимых клетках
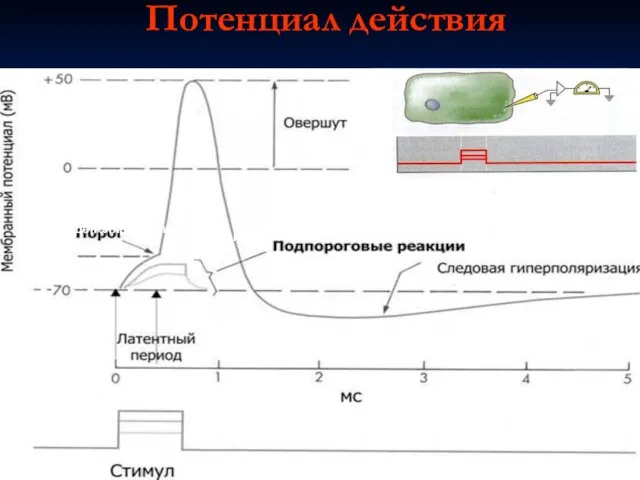
Слайд 16Потенциал действия
Фаза
деполяризации
Фаза
реполяризации
Раздражающий
импульс
Потенциал действия
Фаза
деполяризации
Фаза
реполяризации
Раздражающий
импульс
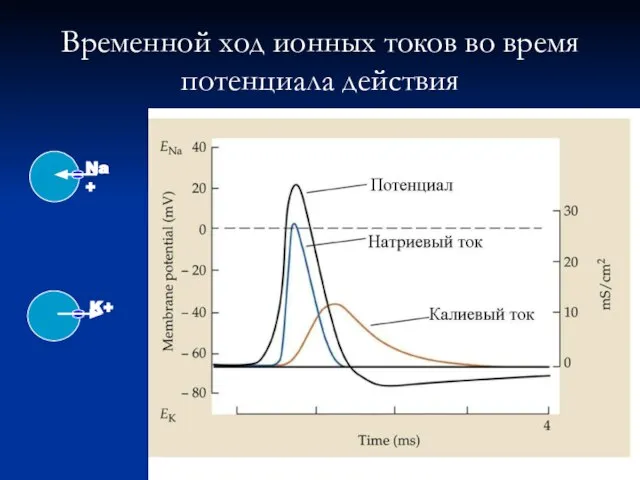
Слайд 17Временной ход ионных токов во время
потенциала действия
Временной ход ионных токов во время
потенциала действия
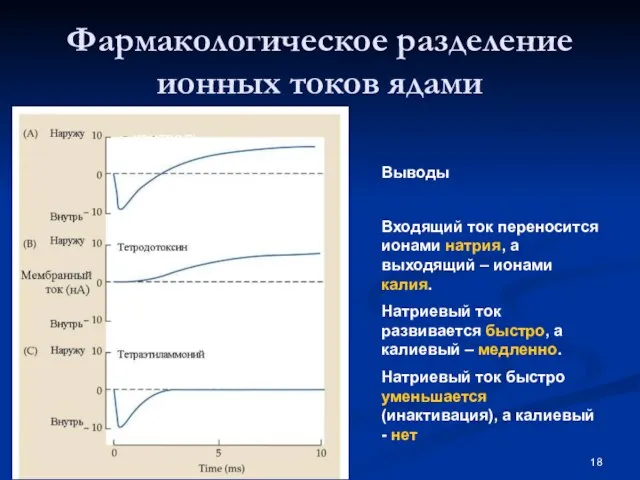
Слайд 18Фармакологическое разделение ионных токов ядами
контроль
Калиевый ток
Натриевый ток
Выводы
Входящий ток переносится ионами натрия, а
Фармакологическое разделение ионных токов ядами
контроль
Калиевый ток
Натриевый ток
Выводы
Входящий ток переносится ионами натрия, а
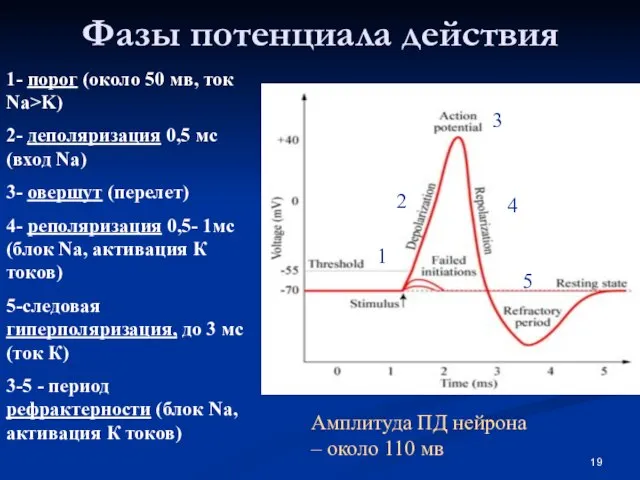
Слайд 19Фазы потенциала действия
1- порог (около 50 мв, ток Na>K)
2- деполяризация 0,5 мс
Фазы потенциала действия
1- порог (около 50 мв, ток Na>K)
2- деполяризация 0,5 мс
Слайд 20Вызывается сверхпороговым раздражением
Амплитуда не зависит от силы раздражения
Распространяется по всей мембране не
Амплитуда не зависит от силы раздражения
Распространяется по всей мембране не

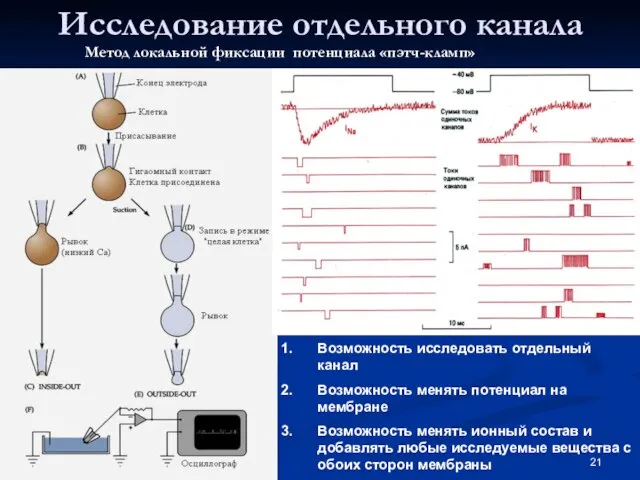
Слайд 21Исследование отдельного канала
Возможность исследовать отдельный канал
Возможность менять потенциал на мембране
Возможность менять ионный
Исследование отдельного канала
Возможность исследовать отдельный канал
Возможность менять потенциал на мембране
Возможность менять ионный
Слайд 22Нобелевская премия 1991 года в области физиологии и медицины
Эрвин Нейер и Берт
Нобелевская премия 1991 года в области физиологии и медицины
Эрвин Нейер и Берт

Слайд 23Канал имеет воротный механизм
1- покой
2-деполяризация
3-рефрактерность
Динамика открытия ворот
1
2
3
За один ПД входит в клетку
Канал имеет воротный механизм
1- покой
2-деполяризация
3-рефрактерность
Динамика открытия ворот
1
2
3
За один ПД входит в клетку
Слайд 24Молекулярные механизмы активации и инактивации
у большинства каналов
общие
H
M
Молекулярные механизмы активации и инактивации
у большинства каналов
общие
H
M
Слайд 25Работа Na+ канала
Работа Na+ канала
Слайд 26Белковая структура канала:
4 домена из 6 сегментов каждый
Структура Cl- канала (консервативны!)
S4-воротный
Белковая структура канала:
4 домена из 6 сегментов каждый
Структура Cl- канала (консервативны!)
S4-воротный
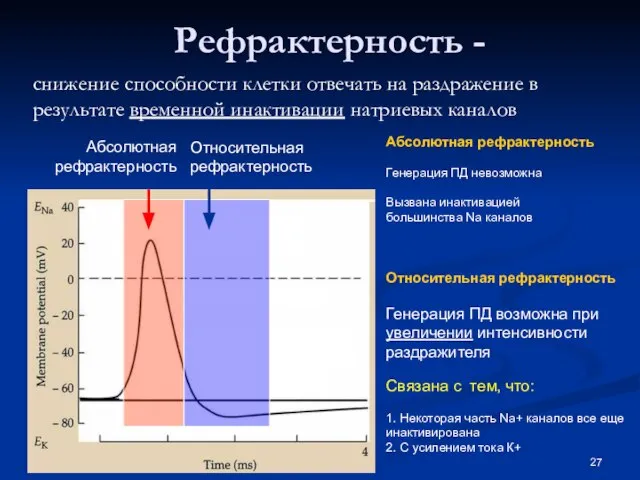
Слайд 27 Рефрактерность -
снижение способности клетки отвечать на раздражение в результате временной инактивации
Рефрактерность -
снижение способности клетки отвечать на раздражение в результате временной инактивации
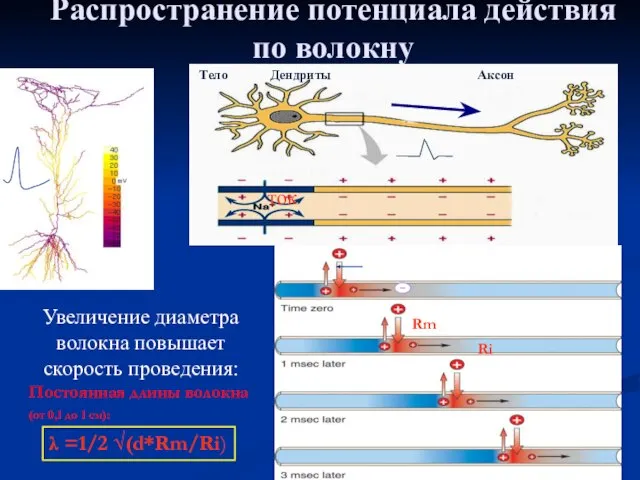
Слайд 28Распространение потенциала действия по волокну
Увеличение диаметра волокна повышает скорость проведения:
Постоянная длины волокна
Распространение потенциала действия по волокну
Увеличение диаметра волокна повышает скорость проведения:
Постоянная длины волокна
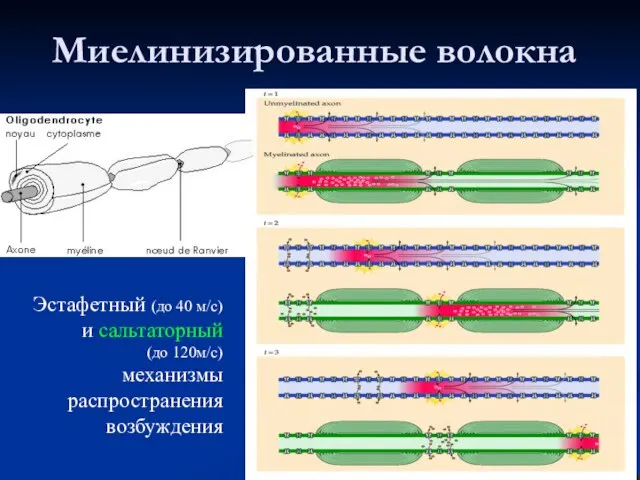
Слайд 29Миелинизированные волокна
Эстафетный (до 40 м/с) и сальтаторный
(до 120м/с)
механизмы распространения возбуждения
Миелинизированные волокна
Эстафетный (до 40 м/с) и сальтаторный
(до 120м/с)
механизмы распространения возбуждения
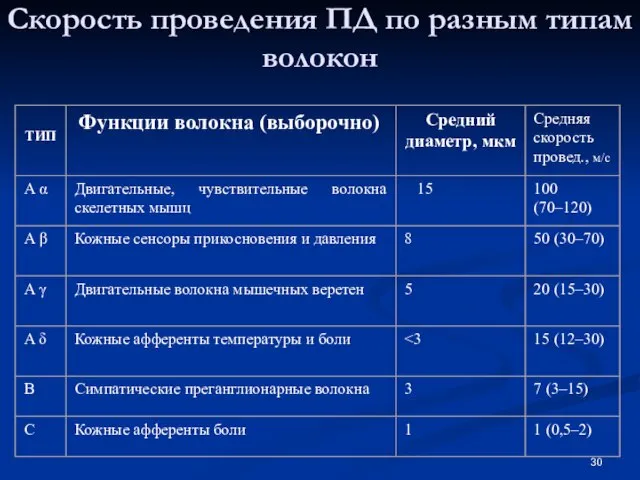
Слайд 30Скорость проведения ПД по разным типам волокон
Скорость проведения ПД по разным типам волокон
Слайд 31Виды регистрации ПД
Внутриклеточная монополярная
Внеклеточная биполярная
Виды регистрации ПД
Внутриклеточная монополярная
Внеклеточная биполярная
 Моу лицей №34
Моу лицей №34 ко дню матери
ко дню матери Урок презентация на тему Мировое хозяйство (4 класс)
Урок презентация на тему Мировое хозяйство (4 класс) Образование будущего в СГЭУ
Образование будущего в СГЭУ Презентация на тему Почему табак называют ядом
Презентация на тему Почему табак называют ядом Оновлені дизайни масла Селянське
Оновлені дизайни масла Селянське Презентация на тему Художник
Презентация на тему Художник workflow
workflow Коммерческое предложение по установке КМС-400/1
Коммерческое предложение по установке КМС-400/1 Презентация на тему Новейшая история
Презентация на тему Новейшая история  Xxiv сессия комитета Бетон, железобетон, преднапряжнный железобетон международной организации по стандартизации ИСО
Xxiv сессия комитета Бетон, железобетон, преднапряжнный железобетон международной организации по стандартизации ИСО Новая Боровая, 1 квартал. Классы жилья
Новая Боровая, 1 квартал. Классы жилья ПРИМЕНЕНИЕ ИТОГОВ С.ПЕТЕРБУРГСКОЙ КОНФЕРЕНЦИИ МЕЖДУНАРОДНЫМ ОБЩЕСТВОМ
ПРИМЕНЕНИЕ ИТОГОВ С.ПЕТЕРБУРГСКОЙ КОНФЕРЕНЦИИ МЕЖДУНАРОДНЫМ ОБЩЕСТВОМ Древняя Спарта
Древняя Спарта We are the world. We are the children
We are the world. We are the children ПЕРСПЕКТИВЫ РАЗВИТИЯ ПЕРЕСТРАХОВОЧНОГО РЫНКА РОССИИ КАК ОДНОГО ИЗ МЕЖДУНАРОДНЫХ ПЕРЕСТРАХОВОЧНЫХ ЦЕНТРОВ
ПЕРСПЕКТИВЫ РАЗВИТИЯ ПЕРЕСТРАХОВОЧНОГО РЫНКА РОССИИ КАК ОДНОГО ИЗ МЕЖДУНАРОДНЫХ ПЕРЕСТРАХОВОЧНЫХ ЦЕНТРОВ Основы построения процесса спортивной тренировки
Основы построения процесса спортивной тренировки Солнечные батареи
Солнечные батареи Сущность и принципы построения организационной структуры управления. Горизонтальное и вертикальное разделение труда
Сущность и принципы построения организационной структуры управления. Горизонтальное и вертикальное разделение труда Корней Чуковский.
Корней Чуковский. Что изучает курс «География материков и океанов»
Что изучает курс «География материков и океанов» Изображение одного и того же предмета в различных цветовых гаммах. Холодная гамма
Изображение одного и того же предмета в различных цветовых гаммах. Холодная гамма Мхи
Мхи Презентация на тему Сатира в начале XX века
Презентация на тему Сатира в начале XX века  Право на труд
Право на труд Технологии Flexus Balasystem для мусорных полигонов современного города
Технологии Flexus Balasystem для мусорных полигонов современного города Вопросы местного значения поселений в сфере жилищных отношений Закон № 131-ФЗ, ст. 14 (поселения), 16 (гор. округа): 1. К вопросам местно
Вопросы местного значения поселений в сфере жилищных отношений Закон № 131-ФЗ, ст. 14 (поселения), 16 (гор. округа): 1. К вопросам местно Итоги работы отрасли и основные направления инновационного развития легкой промышленности
Итоги работы отрасли и основные направления инновационного развития легкой промышленности