- ИММУНИТЕТ И ИНФЕКЦИЯ ДИНАМИЧНОЕ ПРОТИВОСТОЯНИЕ ЖИВЫХ СИСТЕМ

Содержание
- 2. Как защититься? Многоклеточное в мире микробов и простейших
- 3. Встроенный в клеточные мембраны муцин – основной элемент гликокаликса – создает преграду, трудно преодолимую для любых
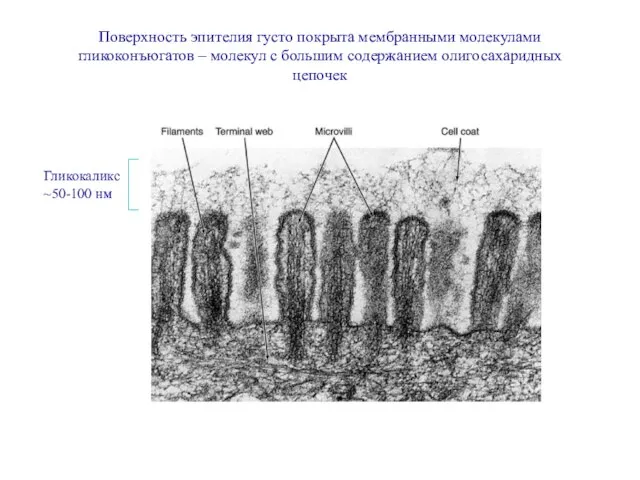
- 4. Гликокаликс ~50-100 нм Поверхность эпителия густо покрыта мембранными молекулами гликоконъюгатов – молекул с большим содержанием олигосахаридных
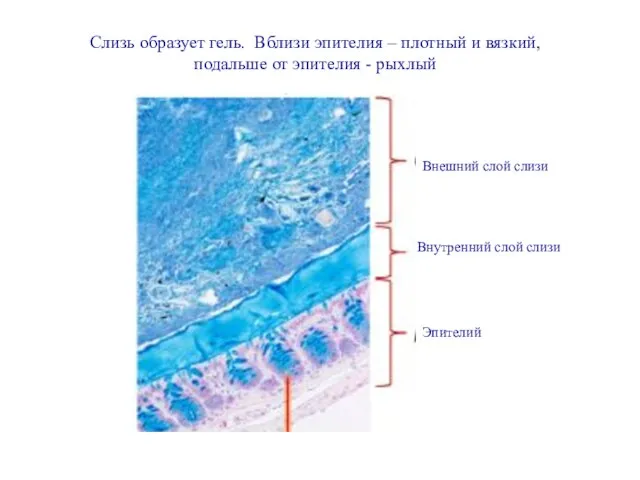
- 5. Внешний слой слизи Внутренний слой слизи Эпителий Слизь образует гель. Вблизи эпителия – плотный и вязкий,
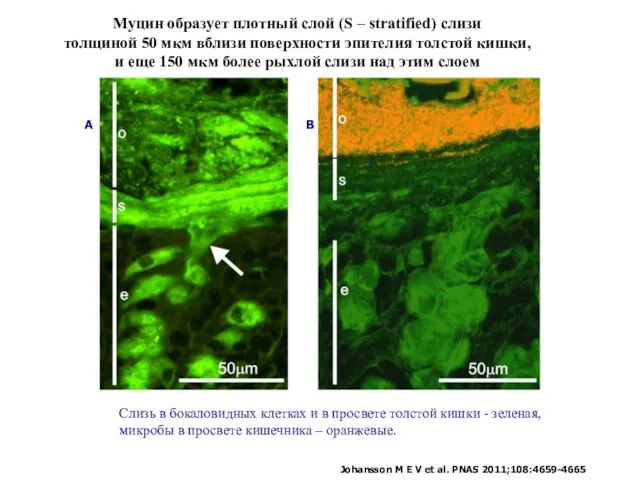
- 6. Муцин образует плотный слой (S – stratified) слизи толщиной 50 мкм вблизи поверхности эпителия толстой кишки,
- 7. Молекулярные сенсоры инфекций и повреждений Toll-like receptors NOD-like receptors RIG-like receptors (helicase, POL-III) C-type lectins Scavenger
- 8. Если инфекция обнаружена, наши клетки выбрасывают вещества, токсичные для микробов
- 9. Дефенсины Кателицидины Гистатины Лактоферрин Лизоцим Псориазин Дермцидин и многие другие… Эндогенные антибиотики – защитные вещества белковой
- 10. Дефенсин-α5 (красный) и лизоцим (зеленый) в слизистой кишки. Широкие стрелки указывают клетки, в которых обнаруживается только
- 11. Клетки эпителия, обнаружившие инфекцию, сообщают соседним клеткам (под эпителием) о том, что произошло вторжение инфекции Сообщения
- 12. IL-1α IL-1β IL-6 IL-8 GM-CSF Фибробласт Макрофаг Дендритная клетка Тучная клетка Сигналы SOS и Danger Бактерии,
- 13. В кишечнике – под эпителием находятся макрофаги и дендритные клетки 20 мкм 20 мкм 50 мкм
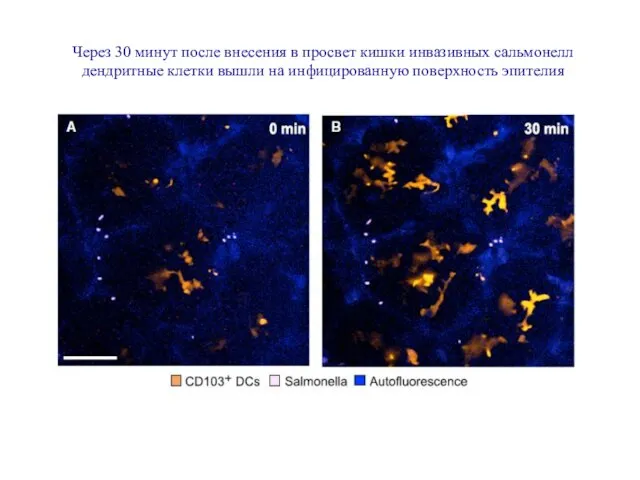
- 14. Через 30 минут после внесения в просвет кишки инвазивных сальмонелл дендритные клетки вышли на инфицированную поверхность
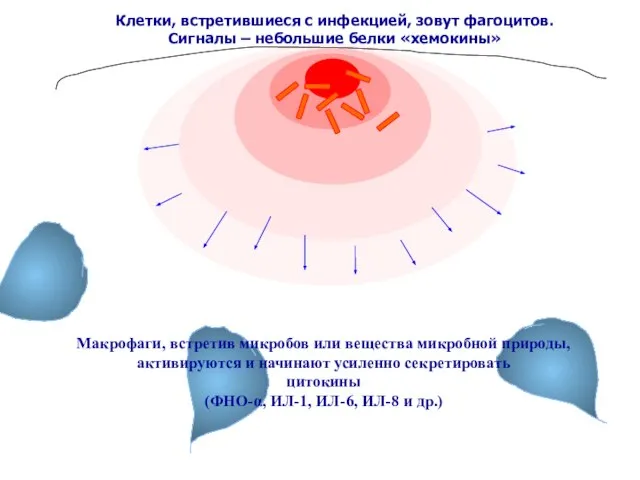
- 15. Макрофаги, встретив микробов или вещества микробной природы, активируются и начинают усиленно секретировать цитокины (ФНО-α, ИЛ-1, ИЛ-6,
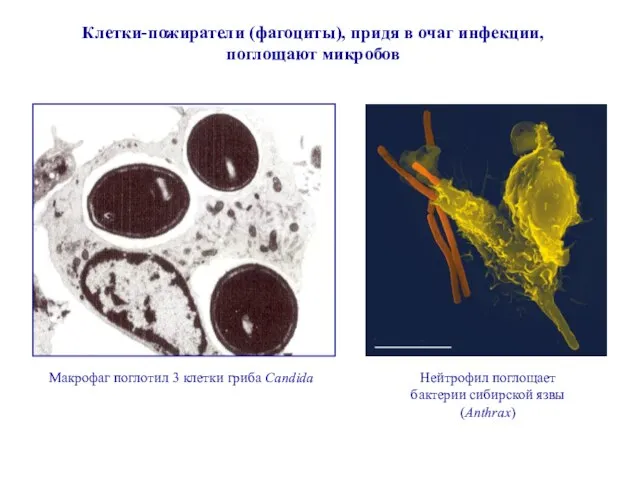
- 16. Клетки-пожиратели (фагоциты), придя в очаг инфекции, поглощают микробов Макрофаг поглотил 3 клетки гриба Candida Нейтрофил поглощает
- 17. Фагоциты, активированные при контакте с инфекцией, выделяют цитокины (ИЛ-1, ФНО, ИЛ-6, ИЛ-8). Эти цитокины действуют на
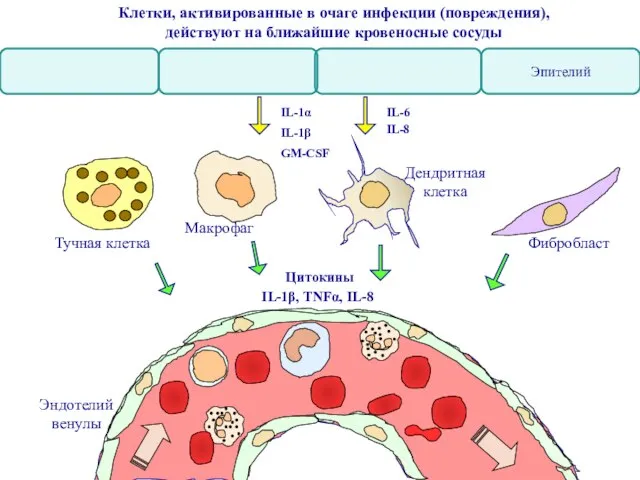
- 18. Эндотелий венулы IL-1α IL-1β IL-6 IL-8 GM-CSF Фибробласт Макрофаг Дендритная клетка Тучная клетка Клетки, активированные в
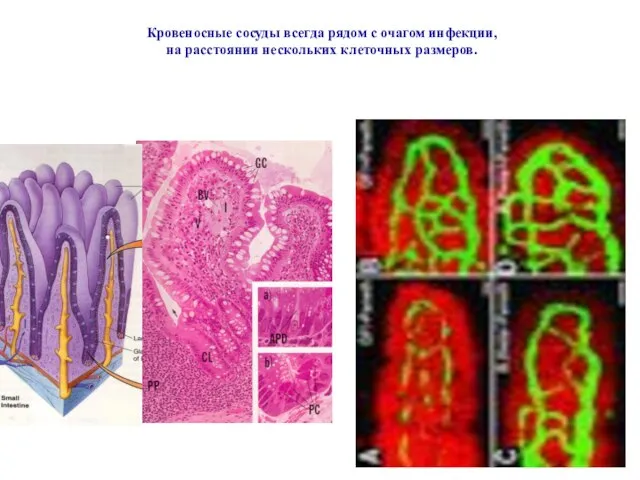
- 19. Кровеносные сосуды всегда рядом с очагом инфекции, на расстоянии нескольких клеточных размеров.
- 20. Два следствия активации эндотелия кровеносных сосудов вблизи очага инфекции: Повышение проницаемости стенки сосуда для крупных молекул
- 21. Вещества, которые выделяют тучные клетки (гистамин), повышают проницаемость стенки кровеносных сосудов. В течение нескольких минут через
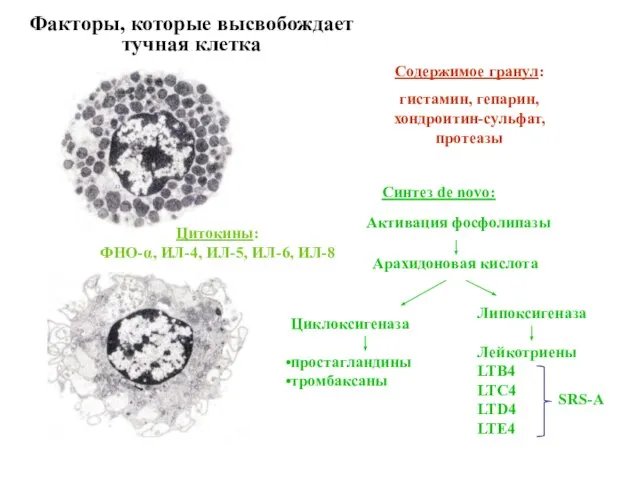
- 22. Факторы, которые высвобождает тучная клетка Содержимое гранул: гистамин, гепарин, хондроитин-сульфат, протеазы Синтез de novo: Активация фосфолипазы
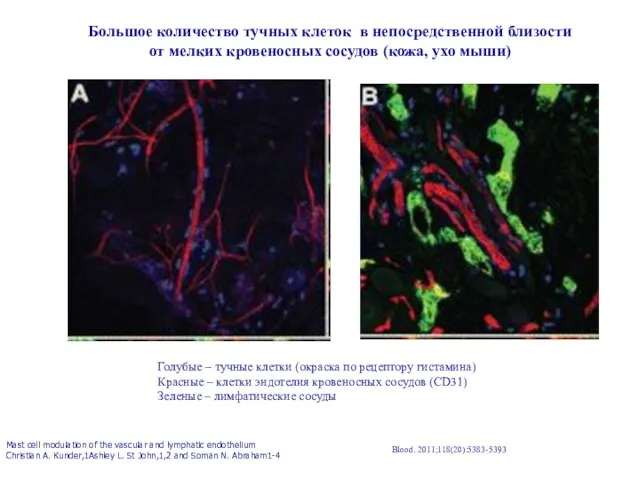
- 23. Большое количество тучных клеток в непосредственной близости от мелких кровеносных сосудов (кожа, ухо мыши) Голубые –
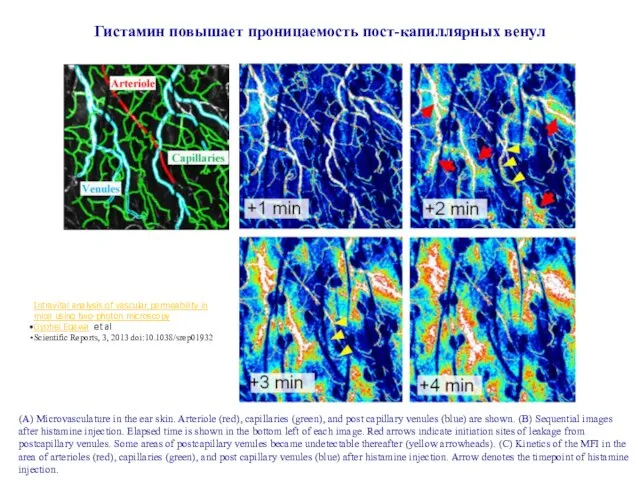
- 24. (A) Microvasculature in the ear skin. Arteriole (red), capillaries (green), and post capillary venules (blue) are
- 25. Крупные молекулы, выходя из кровотока в очаг инфекции, защищают нас от инфекции. Среди защитных молекул, имеющих
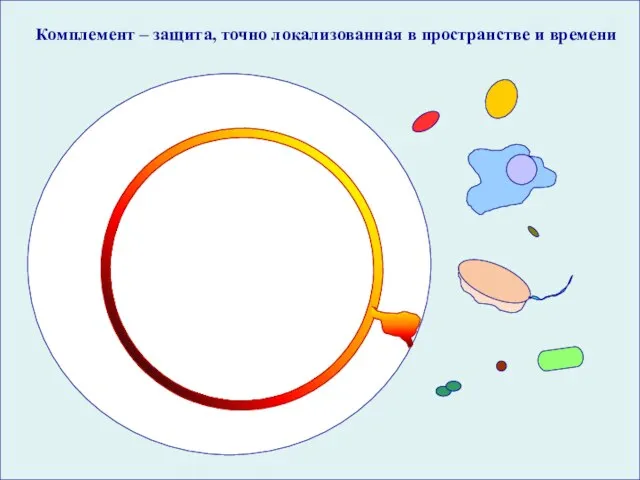
- 26. вирусов и Комплемент – защита, точно локализованная в пространстве и времени
- 27. С3 С1q С8 С2 С1r С6 С4 С1s С3b С2b С4b С5 С5a С5b С9 С7
- 28. При спонтанном гидролизе тиоэфирной группы домены TED и CUB отодвигаются от остальной части молекулы C3. Открывается
- 29. С3 С3b С3b-B D B С3b-Bb Размножение С3b реализуется на доступной поверхности Конвертаза С3 − это
- 30. Три защитных действия комплемента: 1. ОПСОНИЗАЦИЯ ЧАСТИЦ (например, бактериальная частица), на которых активировался комплемент. Поверхность модифицируется
- 31. ФАГОЦИТОЗ ОПСОНИЗИРОВАННОЙ МИКРОБНОЙ ЧАСТИЦЫ Рецепторы С3b Фагоцит
- 32. Конвертаза C5 образует мембрану-атакующий комплекс и вскоре – пору, через которую из клетки вытекают соли и
- 33. Второе следствие активации эндотелия кровеносных сосудов вблизи очага инфекции – это выход лейкоцитов из кровотока
- 34. Активированные макрофаги Нейтрофил Эндотелий венулы Лимфоцит Моноцит Последовательные волны миграции клеток в очаг инфекции. Реакция воспаления.
- 35. вирусов и Организм направляет в очаг инфекции защитные молекулы и защитные клетки
- 36. Если микробы или вещества микробной природы с током тканевой жидкости проникли в лимфатический узел или еще
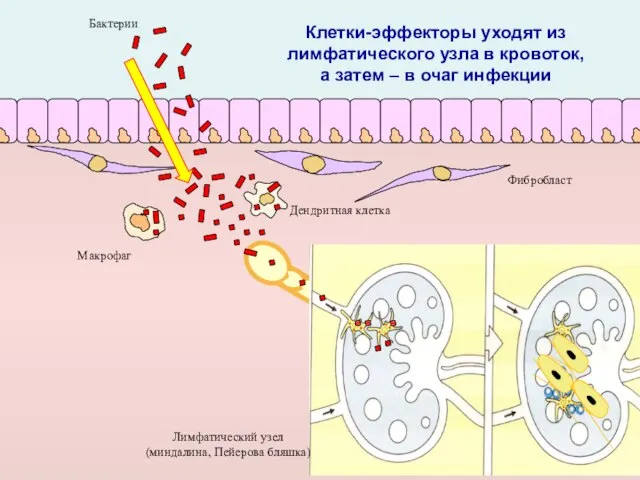
- 37. Внешняя среда Макрофаг Дендритная клетка Фибробласт Лимфатический узел (миндалина, Пейерова бляшка) Бактерии
- 38. В-клетки живут в фолликулах лимфатических узлов
- 39. В клетка Плазматическая клетка
- 40. Ig на клеточной Ig в цитоплазме мембране
- 41. Макрофаг Дендритная клетка Фибробласт Лимфатический узел (миндалина, Пейерова бляшка) Бактерии Клетки-эффекторы уходят из лимфатического узла в
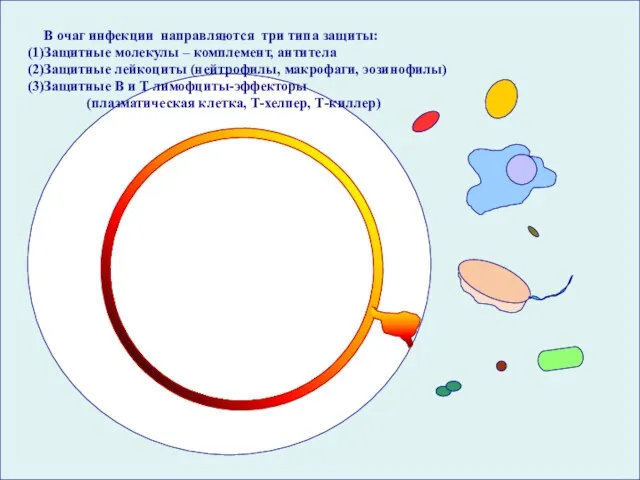
- 43. вирусов и В очаг инфекции направляются три типа защиты: Защитные молекулы – комплемент, антитела Защитные лейкоциты
- 44. Инфекция не уступает нам в оснащенности. Это – сильный, динамичный, умный и даже социально организованный противник
- 45. СТРАТЕГИЧЕСКИЕ ЦЕЛИ ИНФЕКЦИИ Использовать макроорганизм в качестве среды обитания Размножиться Инфицировать следующий макроорганизм
- 46. ТАКТИЧЕСКИЕ ЗАДАЧИ ИНФЕКЦИИ Проникнуть в макроорганизм Избежать ударов иммунной защиты или защититься от них Выжить и
- 47. СТРАТЕГИЧЕСКАЯ ЦЕЛЬ МАКРООРГАНИЗМА Не допустить вторжения инфекции Эффективно защититься от инфекции, если вторжение произошло
- 48. ЗАДАЧИ ИММУНИТЕТА В ОТНОШЕНИЯХ С ИНФЕКЦИЕЙ Не впустить инфекцию Убить инфекцию Удалить инфекцию из организма Залечить
- 49. АРСЕНАЛ ИНФЕКЦИИ Факторы, позволяющие проникать в клетки хозяина Факторы, способные убивать клетки хозяина Факторы, разрушающие тканевой
- 50. АРСЕНАЛ ИММУННОЙ ЗАЩИТЫ ОТ ИНФЕКЦИИ Эндогенные антибиотики (около 100 веществ) Комплемент Интерфероны Фагоциты Натуральные киллеры (NK)
- 51. ИММУННАЯ СИСТЕМА 1. Изобретает принципиальные виды защиты в процессе эволюции биологических видов 2. Нарабатывает огромное разнообразие
- 52. Инфекция тоже активно генерирует разнообразие факторов защиты, чтобы ускользнуть от иммунной атаки
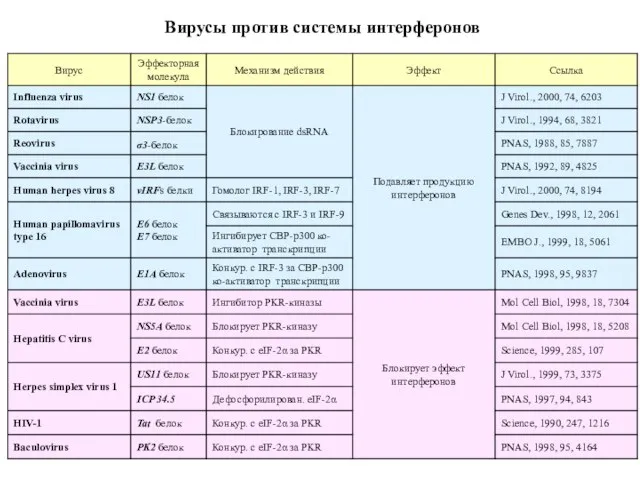
- 53. Вирусы против системы интерферонов
- 54. IFN a/b IRF-3 phosphorylation IRF-5 IRF-7 IRF-9 STAT1 STAT 2 STAT1 STAT 2 IRF-7 IRF-9 ISRE
- 55. БАКТЕРИИ УСКОЛЬЗАЮТ ОТ АНТИТЕЛ, ИЗМЕНЯЯ АНТИГЕНЫ СВОЕЙ ПОВЕРХНОСТИ Streptococcus pneumoniae имеет 84 антигенных варианта капсульных полисахаридов
- 56. Быстрая смена антигенных вариантов VSG-белка у трипаносом (более 1000 генов VSG)
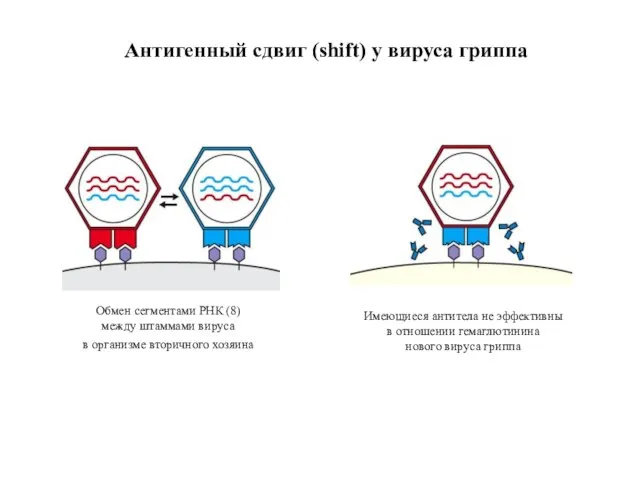
- 57. Антигенный дрейф (drift) у вируса гриппа Нейтрализующие антитела против гемаглютинина блокируют связывание вируса с клеткой Мутации
- 58. Антигенный сдвиг (shift) у вируса гриппа Обмен сегментами РНК (8) между штаммами вируса в организме вторичного
- 60. ИНФЕКЦИЯ И ИММУНИТЕТ ИСПОЛЬЗУЮТ ИДЕНТИЧНЫЙ ГЕНЕТИЧЕСКИЙ КОНСТРУКТОР ДЛЯ СОЗДАНИЯ СРЕДСТВ НАПАДЕНИЯ И ЗАЩИТЫ 1. Сегментарное устройство
- 61. Обе системы – иммунитет и инфекция – очень «интеллектуальны». Они гибко и адекватно используют имеющиеся средства
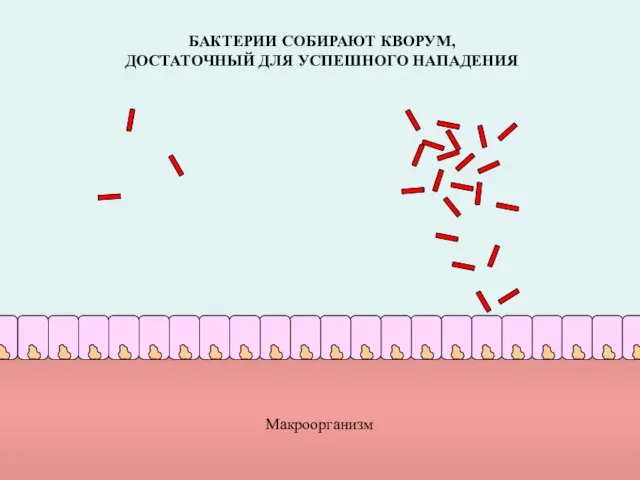
- 62. Бактерии, как люди, объединяются в коллективы для достижения своей цели
- 63. Макроорганизм БАКТЕРИИ СОБИРАЮТ КВОРУМ, ДОСТАТОЧНЫЙ ДЛЯ УСПЕШНОГО НАПАДЕНИЯ
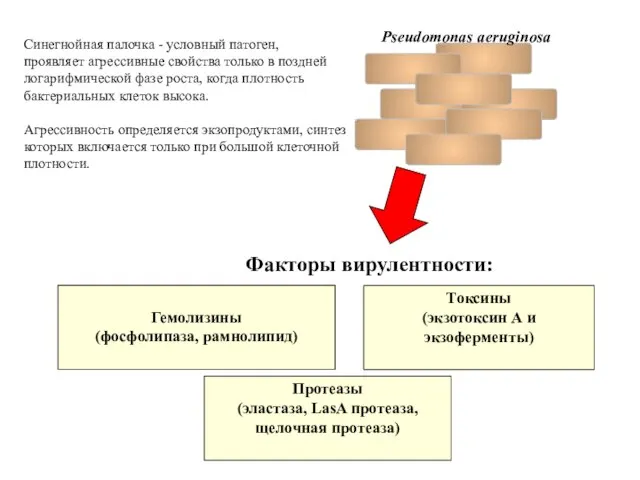
- 64. Синегнойная палочка - условный патоген, проявляет агрессивные свойства только в поздней логарифмической фазе роста, когда плотность
- 65. КАК БАКТЕРИИ ОЩУЩАЮТ КВОРУМ
- 66. Каждая бактерия умеет «говорить» и «слышать». Система Quorum sensing молчит при низкой плотности бактерий в среде
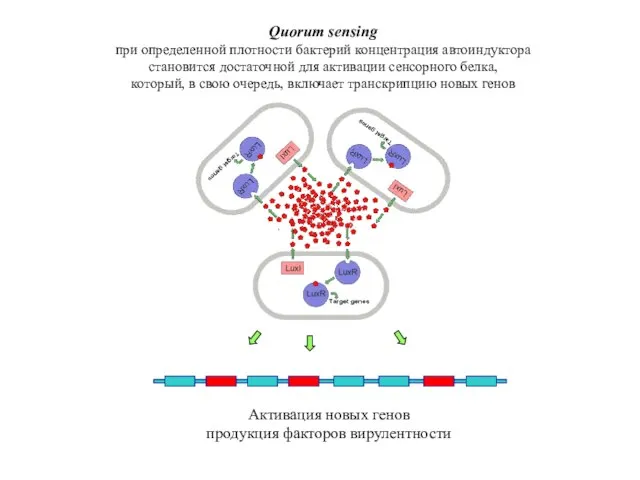
- 67. Активация новых генов продукция факторов вирулентности Quorum sensing при определенной плотности бактерий концентрация автоиндуктора становится достаточной
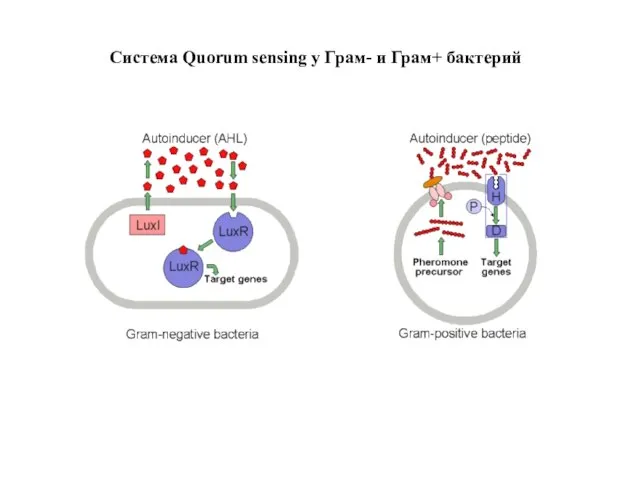
- 68. Система Quorum sensing у Грам- и Грам+ бактерий
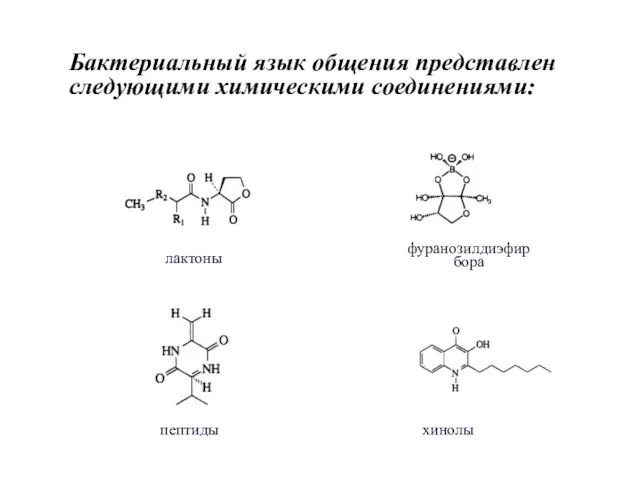
- 69. Бактериальный язык общения представлен следующими химическими соединениями: лактоны хинолы пептиды фуранозилдиэфир бора
- 70. «Механизмы QS позволяют патогенным бактериям очень рационально использовать свой болезнетворный потенциал. Бактрии не атакуют эукариотическую клетку
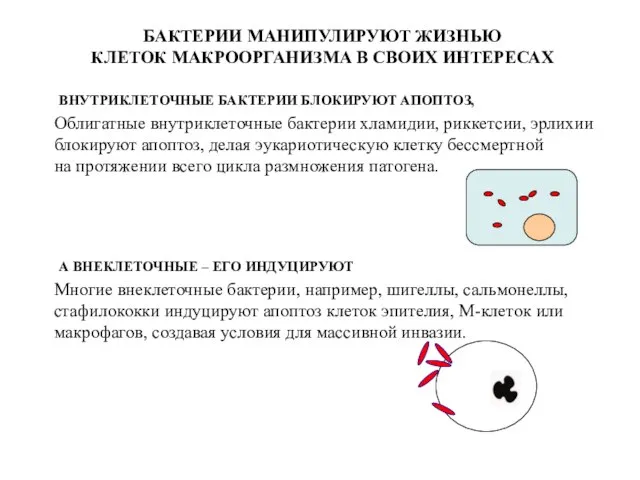
- 71. Облигатные внутриклеточные бактерии хламидии, риккетсии, эрлихии блокируют апоптоз, делая эукариотическую клетку бессмертной на протяжении всего цикла
- 72. Для манипуляций с нашими клетками бактерии используют молекулярный шприц (называется «секреторная система»)
- 74. Типы секреторных систем бактерий The structural biology of type IV secretion systems Rémi Fronzes, Peter J.
- 75. Просвет кишки Стенка кишки Salmonella enterica typhimurium Эпителиальная клетка ПРИНУДИТЕЛЬНЫЙ ЗАХВАТ САЛЬМОНЕЛЛ КЛЕТКАМИ ЭПИТЕЛИЯ КИШЕЧНИКА
- 76. Эпителиальная клетка Просвет кишки SopE SopE2 Actin Actin Salmonella interica typhimurium G-proteins (CDC42, Rac) SopE SopE2
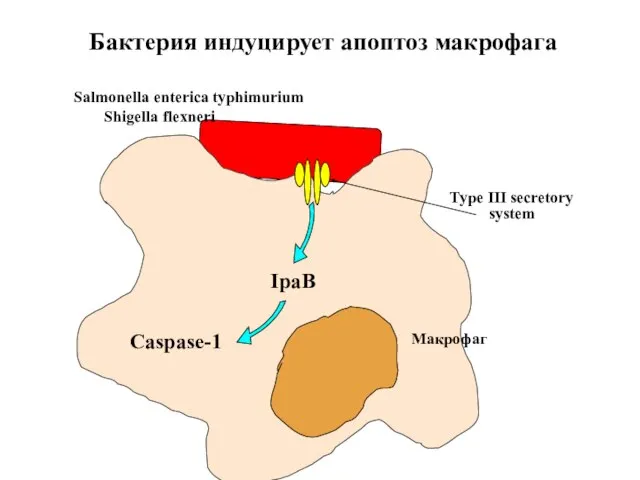
- 77. Макрофаг Salmonella enterica typhimurium Shigella flexneri IpaB Caspase-1 Бактерия индуцирует апоптоз макрофага Type III secretory system
- 78. Динамическое противостояние двух живых систем на примере взаимоотношений вирусной инфекции и иммунитета
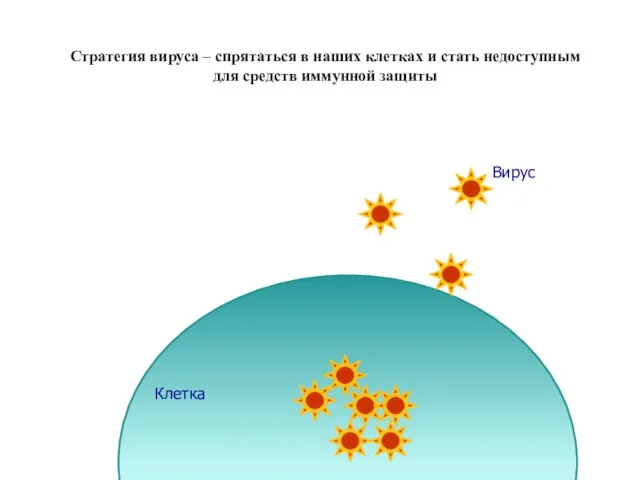
- 79. Стратегия вируса – спрятаться в наших клетках и стать недоступным для средств иммунной защиты Вирус Клетка
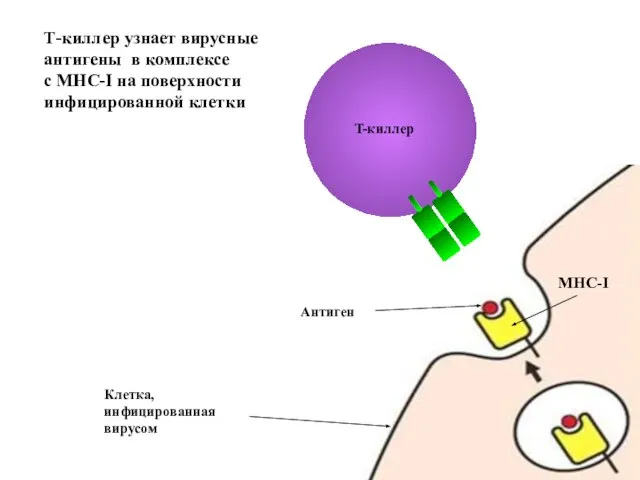
- 80. MHC-I Антиген Т-киллер узнает вирусные антигены в комплексе с МНС-I на поверхности инфицированной клетки Клетка, инфицированная
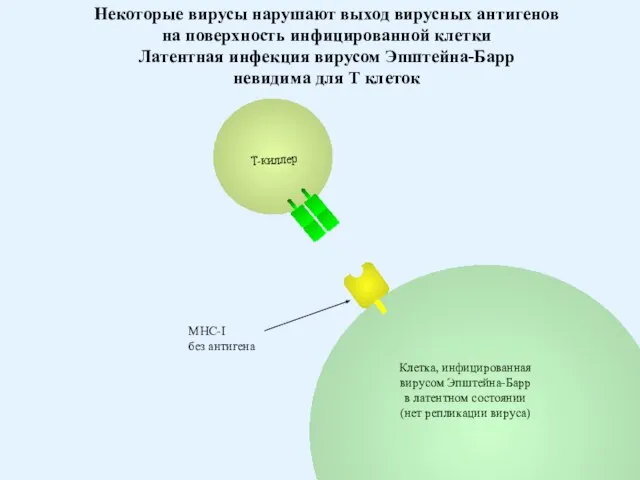
- 81. Клетка, инфицированная вирусом Эпштейна-Барр в латентном состоянии (нет репликации вируса) Некоторые вирусы нарушают выход вирусных антигенов
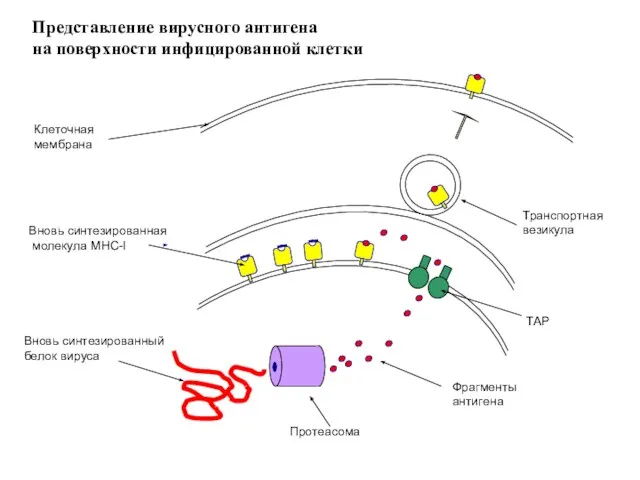
- 82. Вновь синтезированная молекула МНС-I Вновь синтезированный белок вируса Фрагменты антигена Транспортная везикула TAP Клеточная мембрана Представление
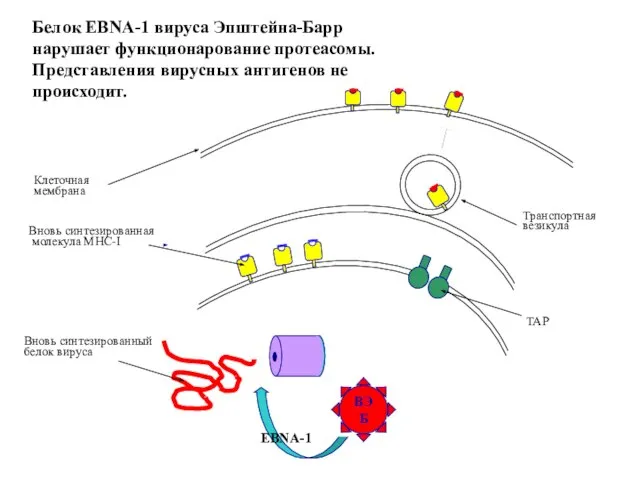
- 83. Вновь синтезированная молекула МНС-I Вновь синтезированный белок вируса Транспортная везикула TAP Клеточная мембрана Белок EBNA-1 вируса
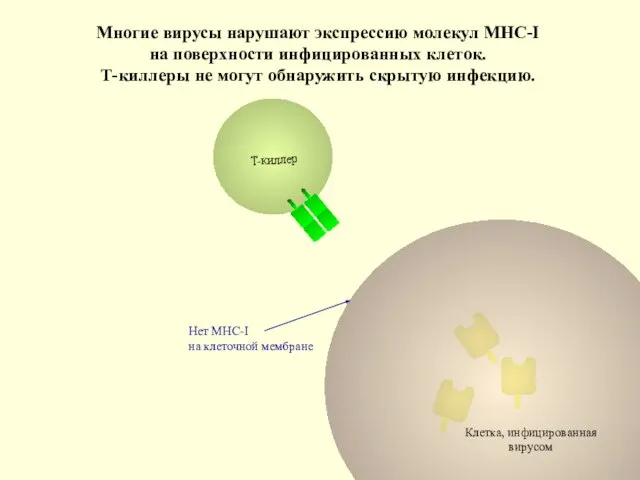
- 84. Клетка, инфицированная вирусом Нет МНС-I на клеточной мембране Многие вирусы нарушают экспрессию молекул МНС-I на поверхности
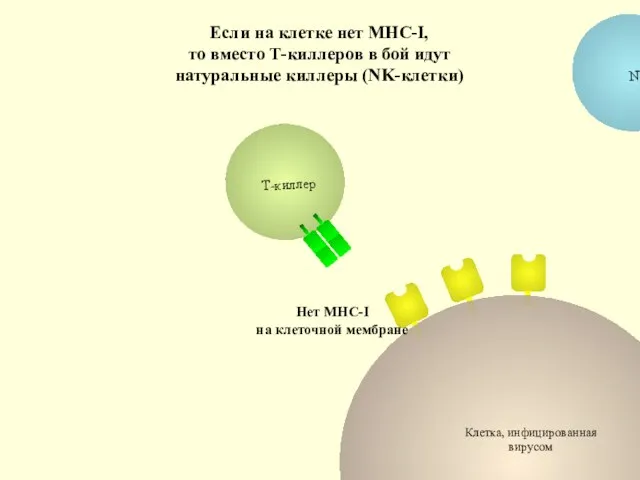
- 85. Клетка, инфицированная вирусом Нет МНС-I на клеточной мембране Если на клетке нет МНС-I, то вместо Т-киллеров
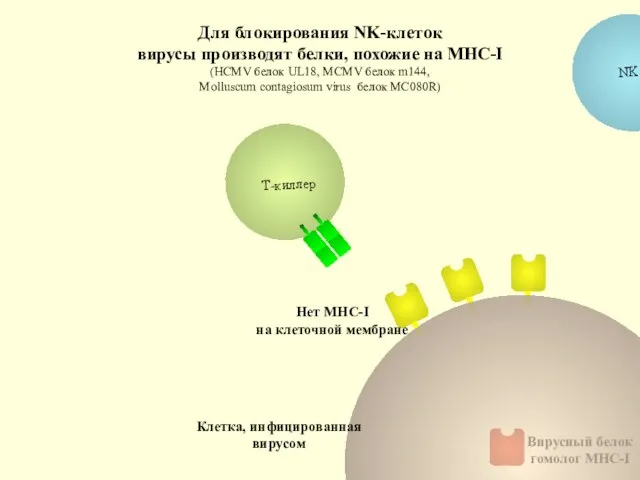
- 86. Для блокирования NK-клеток вирусы производят белки, похожие на МНС-I (HCMV белок UL18, MCMV белок m144, Molluscum
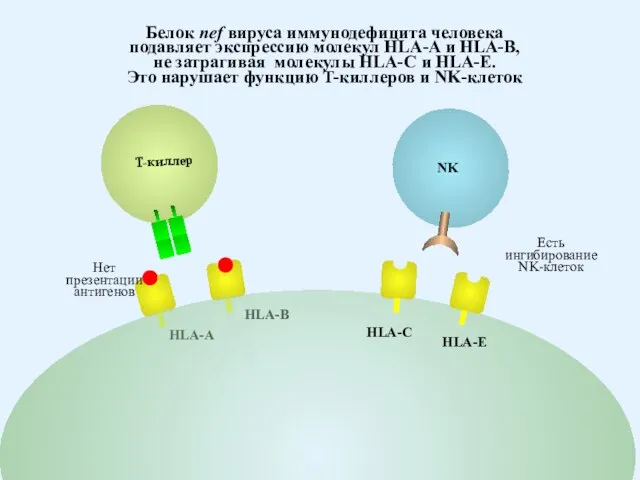
- 87. T-киллер Белок nef вируса иммунодефицита человека подавляет экспрессию молекул HLA-A и HLA-B, не затрагивая молекулы HLA-C
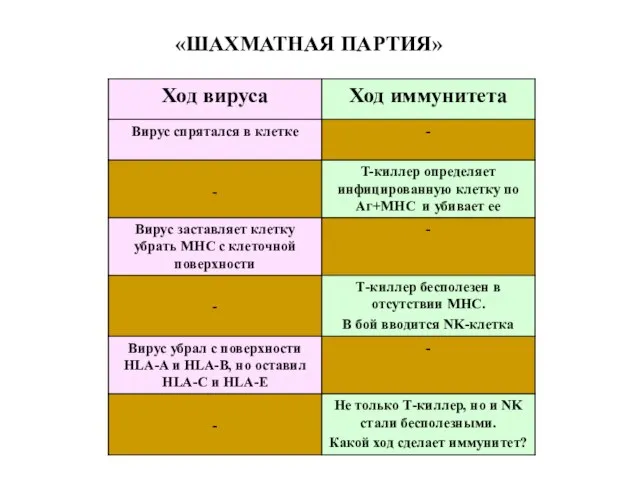
- 88. «ШАХМАТНАЯ ПАРТИЯ»
- 89. УРОКИ: ИММУНИТЕТ И ИНФЕКЦИЯ - ДИНАМИЧНЫЕ И РАЦИОНАЛЬНЫЕ ЖИВЫЕ СИСТЕМЫ, НЕПРЕРЫВНО ИЗМЕНЯЮЩИЕСЯ АДЕКВАТНО ВОЗНИКАЮЩИМ УСЛОВИЯМ. ИНФЕКЦИЯ
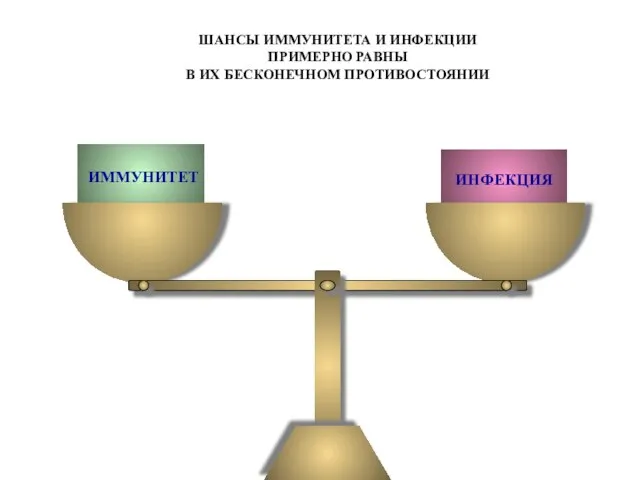
- 90. ИНФЕКЦИЯ ИММУНИТЕТ ШАНСЫ ИММУНИТЕТА И ИНФЕКЦИИ ПРИМЕРНО РАВНЫ В ИХ БЕСКОНЕЧНОМ ПРОТИВОСТОЯНИИ
- 91. ИНФЕКЦИЯ ИММУНИТЕТ ЭТИОТРОПНОЕ ЛЕЧЕНИЕ ДЕЗИНТОКСИКАЦИЯ ОСЛАБЛЕНИЕ ИММУНИТЕТА ВАКЦИНЫ
- 93. Скачать презентацию
Слайд 3Встроенный в клеточные мембраны муцин – основной элемент гликокаликса –
создает преграду,
Встроенный в клеточные мембраны муцин – основной элемент гликокаликса –
создает преграду,

Слайд 4Гликокаликс
~50-100 нм
Поверхность эпителия густо покрыта мембранными молекулами гликоконъюгатов – молекул с
Гликокаликс
~50-100 нм
Поверхность эпителия густо покрыта мембранными молекулами гликоконъюгатов – молекул с
Слайд 5Внешний слой слизи
Внутренний слой слизи
Эпителий
Слизь образует гель. Вблизи эпителия – плотный и
Внешний слой слизи
Внутренний слой слизи
Эпителий
Слизь образует гель. Вблизи эпителия – плотный и
Слайд 6Муцин образует плотный слой (S – stratified) слизи
толщиной 50 мкм вблизи
Муцин образует плотный слой (S – stratified) слизи
толщиной 50 мкм вблизи
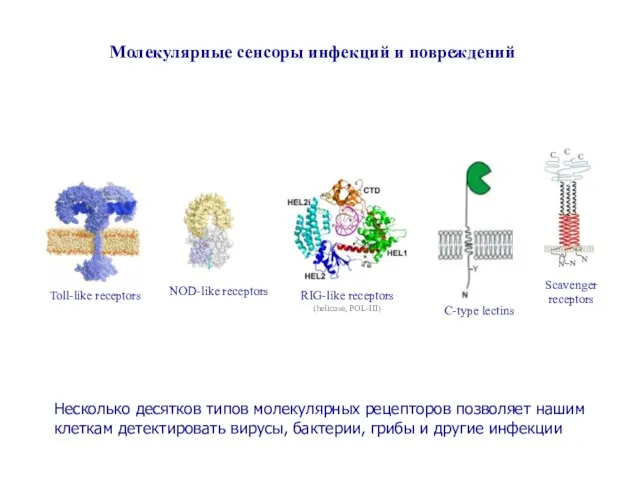
Слайд 7Молекулярные сенсоры инфекций и повреждений
Toll-like receptors
NOD-like receptors
RIG-like receptors
(helicase, POL-III)
C-type lectins
Scavenger
receptors
Несколько десятков
Молекулярные сенсоры инфекций и повреждений
Toll-like receptors
NOD-like receptors
RIG-like receptors
(helicase, POL-III)
C-type lectins
Scavenger
receptors
Несколько десятков
Слайд 8Если инфекция обнаружена, наши клетки выбрасывают вещества, токсичные для микробов
Если инфекция обнаружена, наши клетки выбрасывают вещества, токсичные для микробов

Слайд 9Дефенсины
Кателицидины
Гистатины
Лактоферрин
Лизоцим
Псориазин
Дермцидин
и многие другие…
Эндогенные антибиотики –
защитные вещества белковой и пептидной природы
наиболее активно
Дефенсины
Кателицидины
Гистатины
Лактоферрин
Лизоцим
Псориазин
Дермцидин
и многие другие…
Эндогенные антибиотики –
защитные вещества белковой и пептидной природы
наиболее активно

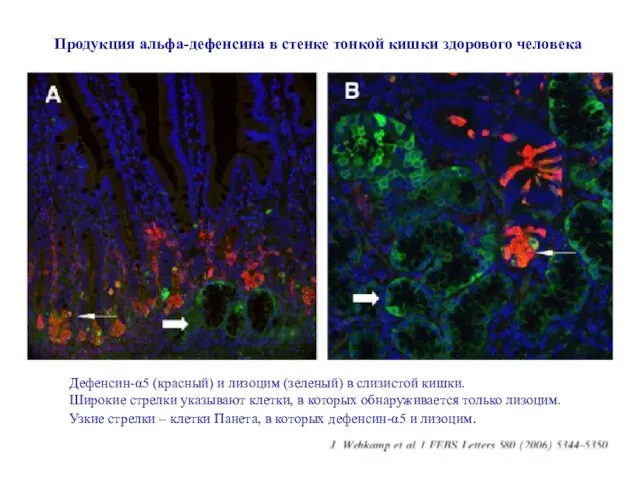
Слайд 10Дефенсин-α5 (красный) и лизоцим (зеленый) в слизистой кишки.
Широкие стрелки указывают клетки,
Дефенсин-α5 (красный) и лизоцим (зеленый) в слизистой кишки.
Широкие стрелки указывают клетки,
Слайд 11Клетки эпителия, обнаружившие инфекцию,
сообщают соседним клеткам (под эпителием)
о том, что произошло вторжение
Клетки эпителия, обнаружившие инфекцию,
сообщают соседним клеткам (под эпителием)
о том, что произошло вторжение

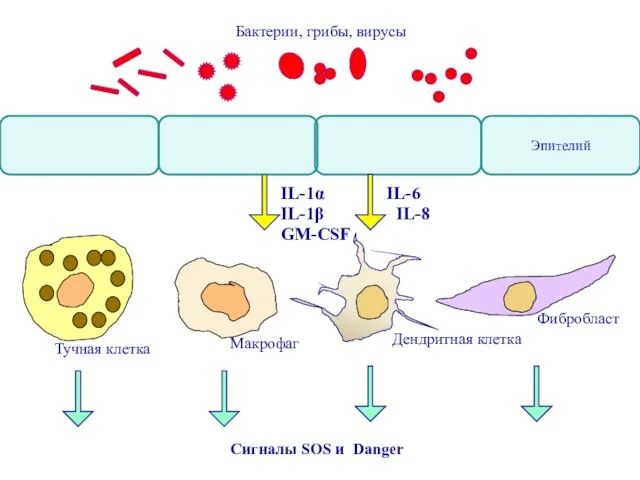
Слайд 12IL-1α
IL-1β
IL-6
IL-8
GM-CSF
Фибробласт
Макрофаг
Дендритная клетка
Тучная клетка
Сигналы SOS и Danger
Бактерии, грибы, вирусы
IL-1α
IL-1β
IL-6
IL-8
GM-CSF
Фибробласт
Макрофаг
Дендритная клетка
Тучная клетка
Сигналы SOS и Danger
Бактерии, грибы, вирусы
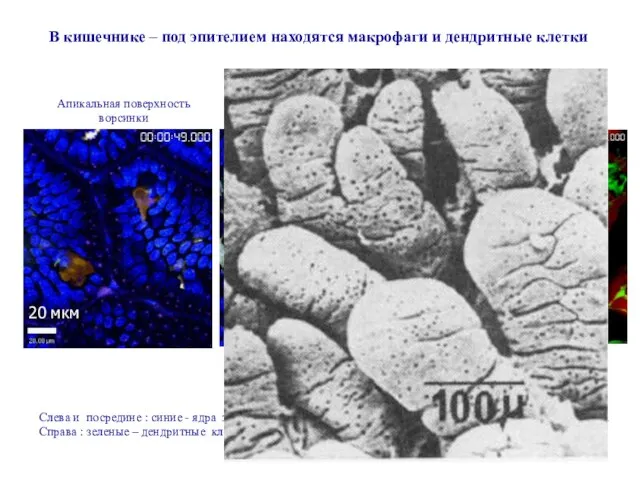
Слайд 13В кишечнике – под эпителием находятся макрофаги и дендритные клетки
20 мкм
20 мкм
50
В кишечнике – под эпителием находятся макрофаги и дендритные клетки
20 мкм
20 мкм
50
Слайд 14Через 30 минут после внесения в просвет кишки инвазивных сальмонелл
дендритные клетки вышли
Через 30 минут после внесения в просвет кишки инвазивных сальмонелл
дендритные клетки вышли
Слайд 15Макрофаги, встретив микробов или вещества микробной природы,
активируются и начинают усиленно секретировать
Макрофаги, встретив микробов или вещества микробной природы,
активируются и начинают усиленно секретировать
Слайд 16Клетки-пожиратели (фагоциты), придя в очаг инфекции,
поглощают микробов
Макрофаг поглотил 3 клетки гриба Candida
Нейтрофил
Клетки-пожиратели (фагоциты), придя в очаг инфекции,
поглощают микробов
Макрофаг поглотил 3 клетки гриба Candida
Нейтрофил
Слайд 17Фагоциты, активированные при контакте с инфекцией, выделяют цитокины (ИЛ-1, ФНО, ИЛ-6, ИЛ-8).
Эти
Фагоциты, активированные при контакте с инфекцией, выделяют цитокины (ИЛ-1, ФНО, ИЛ-6, ИЛ-8).
Эти

Слайд 18Эндотелий
венулы
IL-1α
IL-1β
IL-6
IL-8
GM-CSF
Фибробласт
Макрофаг
Дендритная
клетка
Тучная клетка
Клетки, активированные в очаге инфекции (повреждения),
действуют на ближайшие кровеносные сосуды
Цитокины
IL-1β,
Эндотелий
венулы
IL-1α
IL-1β
IL-6
IL-8
GM-CSF
Фибробласт
Макрофаг
Дендритная
клетка
Тучная клетка
Клетки, активированные в очаге инфекции (повреждения),
действуют на ближайшие кровеносные сосуды
Цитокины
IL-1β,
Слайд 19Кровеносные сосуды всегда рядом с очагом инфекции,
на расстоянии нескольких клеточных размеров.
Кровеносные сосуды всегда рядом с очагом инфекции, на расстоянии нескольких клеточных размеров.
Слайд 20Два следствия активации эндотелия кровеносных
сосудов вблизи очага инфекции:
Повышение проницаемости стенки сосуда
Два следствия активации эндотелия кровеносных
сосудов вблизи очага инфекции:
Повышение проницаемости стенки сосуда

Слайд 21Вещества, которые выделяют тучные клетки (гистамин), повышают проницаемость стенки кровеносных сосудов.
В течение
Вещества, которые выделяют тучные клетки (гистамин), повышают проницаемость стенки кровеносных сосудов.
В течение

Слайд 22Факторы, которые высвобождает тучная клетка
Содержимое гранул:
гистамин, гепарин,
хондроитин-сульфат,
протеазы
Синтез de novo:
Активация фосфолипазы
Арахидоновая кислота
Циклоксигеназа
простагландины
тромбаксаны
Липоксигеназа
Лейкотриены
LTB4
LTC4
LTD4
LTE4
SRS-A
Цитокины:
Факторы, которые высвобождает тучная клетка
Содержимое гранул:
гистамин, гепарин,
хондроитин-сульфат,
протеазы
Синтез de novo:
Активация фосфолипазы
Арахидоновая кислота
Циклоксигеназа
простагландины
тромбаксаны
Липоксигеназа
Лейкотриены
LTB4
LTC4
LTD4
LTE4
SRS-A
Цитокины:
Слайд 23Большое количество тучных клеток в непосредственной близости от мелких кровеносных сосудов (кожа,
Большое количество тучных клеток в непосредственной близости от мелких кровеносных сосудов (кожа,
Слайд 24(A) Microvasculature in the ear skin. Arteriole (red), capillaries (green), and post
(A) Microvasculature in the ear skin. Arteriole (red), capillaries (green), and post
Слайд 25Крупные молекулы, выходя из кровотока в очаг инфекции,
защищают нас от инфекции.
Среди защитных
Крупные молекулы, выходя из кровотока в очаг инфекции,
защищают нас от инфекции.
Среди защитных

Слайд 26вирусов и
Комплемент – защита, точно локализованная в пространстве и времени
вирусов и
Комплемент – защита, точно локализованная в пространстве и времени
Слайд 27С3
С1q
С8
С2
С1r
С6
С4
С1s
С3b
С2b
С4b
С5
С5a
С5b
С9
С7
I
С4bp
H
P
DAF
D
Bb
MCP
СD59
MASP-
1
MASP-
2
CR1
CR2
H
CR4
H
H
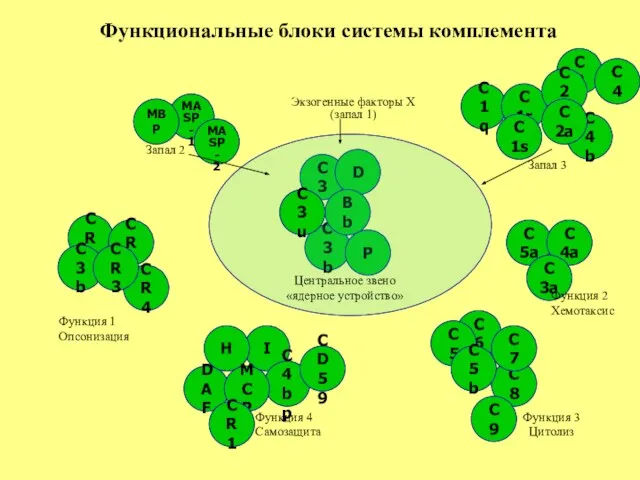
Функциональные блоки системы комплемента
С2a
MBP
CR1
С4a
С3a
С3u
Центральное звено
«ядерное устройство»
Функция 2
Хемотаксис
Запал 3
Функция 3
Цитолиз
Функция 4
Самозащита
Функция
С3
С1q
С8
С2
С1r
С6
С4
С1s
С3b
С2b
С4b
С5
С5a
С5b
С9
С7
I
С4bp
H
P
DAF
D
Bb
MCP
СD59
MASP-
1
MASP-
2
CR1
CR2
H
CR4
H
H
Функциональные блоки системы комплемента
С2a
MBP
CR1
С4a
С3a
С3u
Центральное звено
«ядерное устройство»
Функция 2
Хемотаксис
Запал 3
Функция 3
Цитолиз
Функция 4
Самозащита
Функция
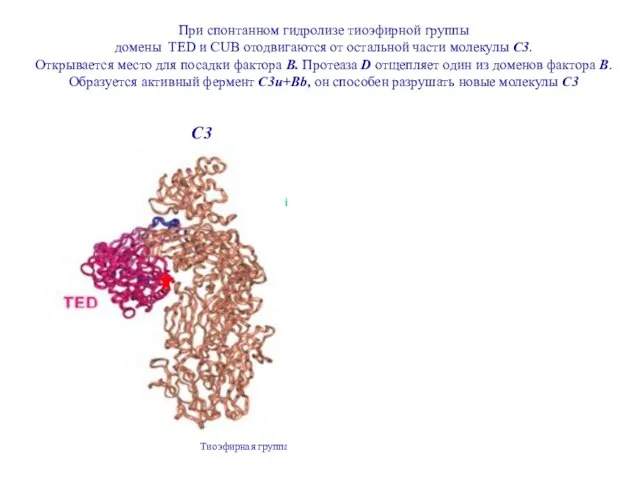
Слайд 28При спонтанном гидролизе тиоэфирной группы
домены TED и CUB отодвигаются от остальной части
При спонтанном гидролизе тиоэфирной группы
домены TED и CUB отодвигаются от остальной части
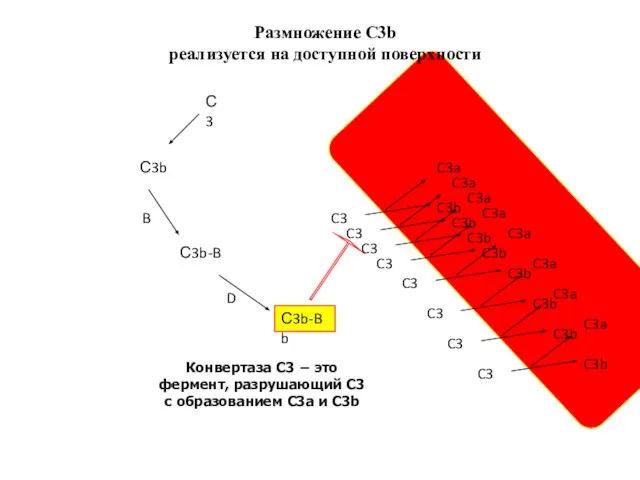
Слайд 29С3
С3b
С3b-B
D
B
С3b-Bb
Размножение С3b
реализуется на доступной поверхности
Конвертаза С3 − это фермент, разрушающий С3
с
С3
С3b
С3b-B
D
B
С3b-Bb
Размножение С3b
реализуется на доступной поверхности
Конвертаза С3 − это фермент, разрушающий С3
с
Слайд 30Три защитных действия комплемента:
1. ОПСОНИЗАЦИЯ ЧАСТИЦ (например, бактериальная частица), на которых активировался
Три защитных действия комплемента:
1. ОПСОНИЗАЦИЯ ЧАСТИЦ (например, бактериальная частица), на которых активировался

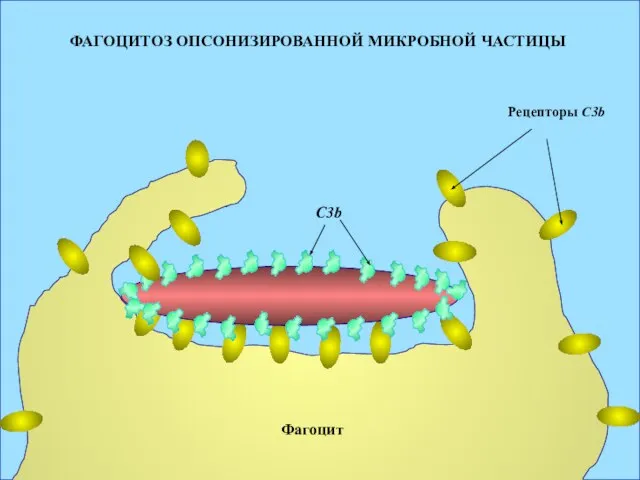
Слайд 31ФАГОЦИТОЗ ОПСОНИЗИРОВАННОЙ МИКРОБНОЙ ЧАСТИЦЫ
Рецепторы С3b
Фагоцит
ФАГОЦИТОЗ ОПСОНИЗИРОВАННОЙ МИКРОБНОЙ ЧАСТИЦЫ
Рецепторы С3b
Фагоцит
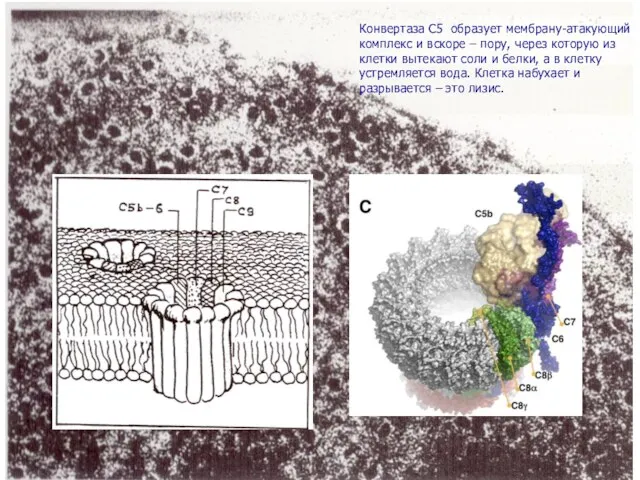
Слайд 32Конвертаза C5 образует мембрану-атакующий комплекс и вскоре – пору, через которую из
Конвертаза C5 образует мембрану-атакующий комплекс и вскоре – пору, через которую из
Слайд 33Второе следствие активации эндотелия кровеносных сосудов вблизи очага инфекции – это выход
Второе следствие активации эндотелия кровеносных сосудов вблизи очага инфекции – это выход

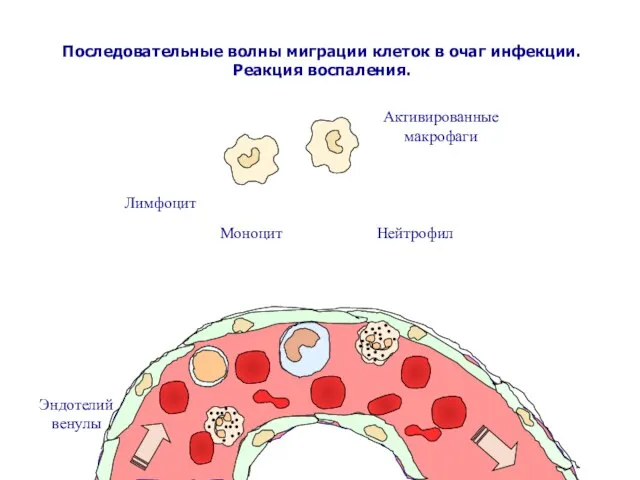
Слайд 34Активированные макрофаги
Нейтрофил
Эндотелий
венулы
Лимфоцит
Моноцит
Последовательные волны миграции клеток в очаг инфекции.
Реакция воспаления.
Активированные макрофаги
Нейтрофил
Эндотелий
венулы
Лимфоцит
Моноцит
Последовательные волны миграции клеток в очаг инфекции.
Реакция воспаления.
Слайд 35вирусов и
Организм направляет в очаг инфекции
защитные молекулы и защитные клетки
вирусов и
Организм направляет в очаг инфекции
защитные молекулы и защитные клетки
Слайд 36Если микробы или вещества микробной природы с током тканевой жидкости проникли в
Если микробы или вещества микробной природы с током тканевой жидкости проникли в

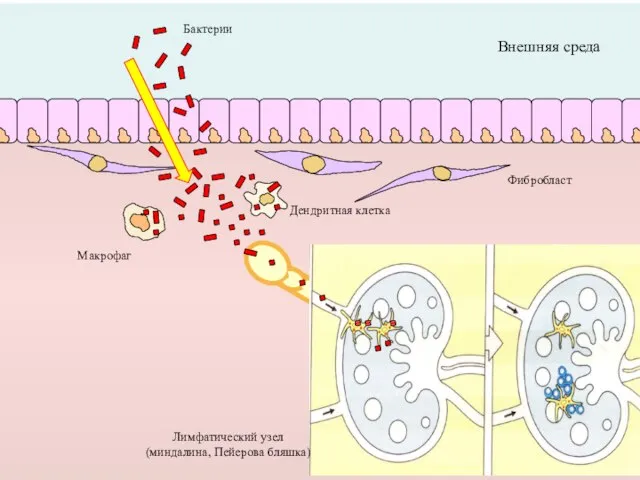
Слайд 37Внешняя среда
Макрофаг
Дендритная клетка
Фибробласт
Лимфатический узел
(миндалина, Пейерова бляшка)
Бактерии
Внешняя среда
Макрофаг
Дендритная клетка
Фибробласт
Лимфатический узел
(миндалина, Пейерова бляшка)
Бактерии
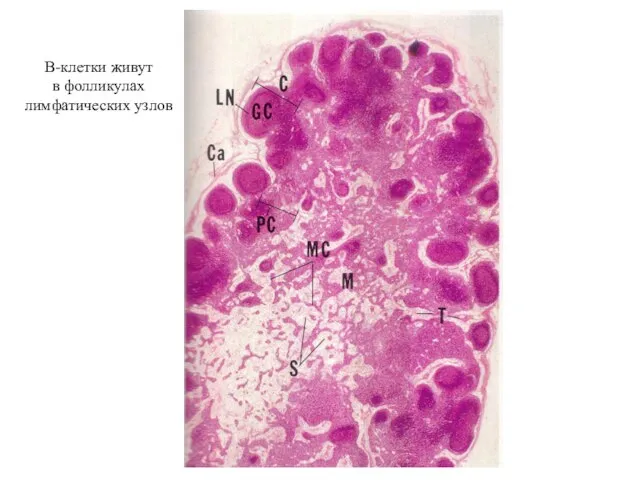
Слайд 38В-клетки живут
в фолликулах
лимфатических узлов
В-клетки живут
в фолликулах
лимфатических узлов
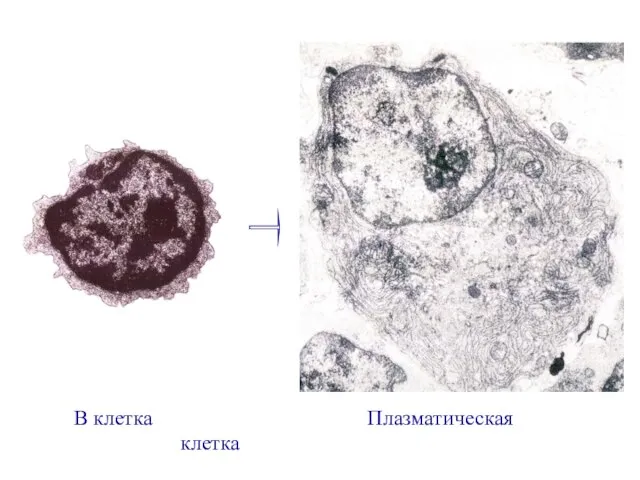
Слайд 39В клетка Плазматическая клетка
В клетка Плазматическая клетка
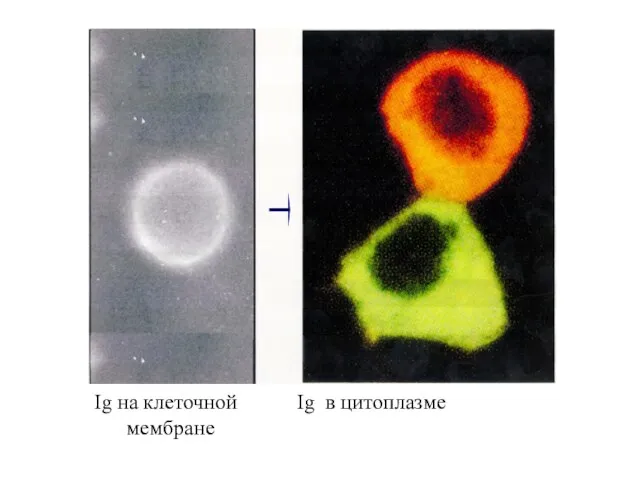
Слайд 40Ig на клеточной Ig в цитоплазме
мембране
Ig на клеточной Ig в цитоплазме
мембране
Слайд 41Макрофаг
Дендритная клетка
Фибробласт
Лимфатический узел
(миндалина, Пейерова бляшка)
Бактерии
Клетки-эффекторы уходят из лимфатического узла в кровоток,
Макрофаг
Дендритная клетка
Фибробласт
Лимфатический узел
(миндалина, Пейерова бляшка)
Бактерии
Клетки-эффекторы уходят из лимфатического узла в кровоток,
Слайд 43вирусов и
В очаг инфекции направляются три типа защиты:
Защитные молекулы – комплемент, антитела
Защитные
вирусов и
В очаг инфекции направляются три типа защиты:
Защитные молекулы – комплемент, антитела
Защитные
Слайд 44Инфекция не уступает нам в оснащенности.
Это – сильный, динамичный, умный и даже
Инфекция не уступает нам в оснащенности.
Это – сильный, динамичный, умный и даже

Слайд 45СТРАТЕГИЧЕСКИЕ ЦЕЛИ ИНФЕКЦИИ
Использовать макроорганизм
в качестве среды обитания
Размножиться
Инфицировать следующий макроорганизм
СТРАТЕГИЧЕСКИЕ ЦЕЛИ ИНФЕКЦИИ
Использовать макроорганизм
в качестве среды обитания
Размножиться
Инфицировать следующий макроорганизм

Слайд 46ТАКТИЧЕСКИЕ ЗАДАЧИ ИНФЕКЦИИ
Проникнуть в макроорганизм
Избежать ударов иммунной защиты или защититься от них
ТАКТИЧЕСКИЕ ЗАДАЧИ ИНФЕКЦИИ
Проникнуть в макроорганизм
Избежать ударов иммунной защиты или защититься от них

Слайд 47СТРАТЕГИЧЕСКАЯ ЦЕЛЬ МАКРООРГАНИЗМА
Не допустить вторжения инфекции
Эффективно защититься от инфекции,
если вторжение произошло
СТРАТЕГИЧЕСКАЯ ЦЕЛЬ МАКРООРГАНИЗМА
Не допустить вторжения инфекции
Эффективно защититься от инфекции,
если вторжение произошло

Слайд 48ЗАДАЧИ ИММУНИТЕТА
В ОТНОШЕНИЯХ С ИНФЕКЦИЕЙ
Не впустить инфекцию
Убить инфекцию
Удалить инфекцию из организма
Залечить
ЗАДАЧИ ИММУНИТЕТА
В ОТНОШЕНИЯХ С ИНФЕКЦИЕЙ
Не впустить инфекцию
Убить инфекцию
Удалить инфекцию из организма
Залечить

Слайд 49АРСЕНАЛ ИНФЕКЦИИ
Факторы, позволяющие проникать в клетки хозяина
Факторы, способные убивать клетки хозяина
Факторы, разрушающие
АРСЕНАЛ ИНФЕКЦИИ
Факторы, позволяющие проникать в клетки хозяина
Факторы, способные убивать клетки хозяина
Факторы, разрушающие

Слайд 50АРСЕНАЛ ИММУННОЙ ЗАЩИТЫ
ОТ ИНФЕКЦИИ
Эндогенные антибиотики (около 100 веществ)
Комплемент
Интерфероны
Фагоциты
Натуральные киллеры (NK)
Т клетки-киллеры
АРСЕНАЛ ИММУННОЙ ЗАЩИТЫ
ОТ ИНФЕКЦИИ
Эндогенные антибиотики (около 100 веществ)
Комплемент
Интерфероны
Фагоциты
Натуральные киллеры (NK)
Т клетки-киллеры

Слайд 51ИММУННАЯ СИСТЕМА
1. Изобретает принципиальные виды защиты
в процессе эволюции биологических видов
2. Нарабатывает огромное
ИММУННАЯ СИСТЕМА
1. Изобретает принципиальные виды защиты
в процессе эволюции биологических видов
2. Нарабатывает огромное

Слайд 52Инфекция тоже активно генерирует
разнообразие факторов защиты,
чтобы ускользнуть от иммунной атаки
Инфекция тоже активно генерирует
разнообразие факторов защиты,
чтобы ускользнуть от иммунной атаки

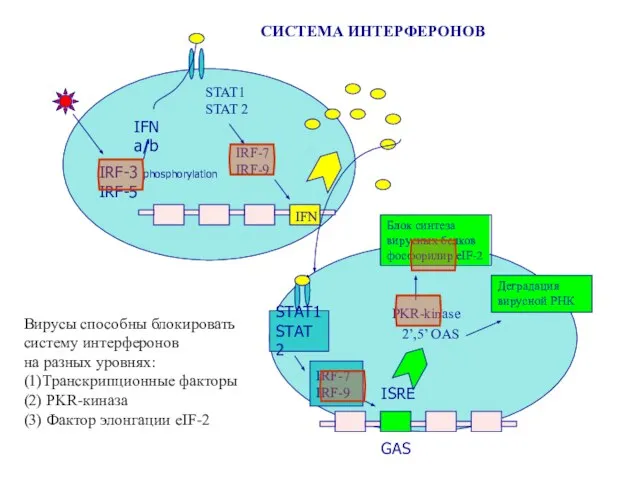
Слайд 53Вирусы против системы интерферонов
Вирусы против системы интерферонов
Слайд 54IFN a/b
IRF-3 phosphorylation
IRF-5
IRF-7
IRF-9
STAT1
STAT 2
STAT1
STAT 2
IRF-7
IRF-9
ISRE
GAS
PKR-kinase
2’,5’ OAS
Блок синтеза
вирусных белков
фосфорилир eIF-2
Деградация
вирусной РНК
IFN
СИСТЕМА
IFN a/b
IRF-3 phosphorylation
IRF-5
IRF-7
IRF-9
STAT1
STAT 2
STAT1
STAT 2
IRF-7
IRF-9
ISRE
GAS
PKR-kinase
2’,5’ OAS
Блок синтеза
вирусных белков
фосфорилир eIF-2
Деградация
вирусной РНК
IFN
СИСТЕМА
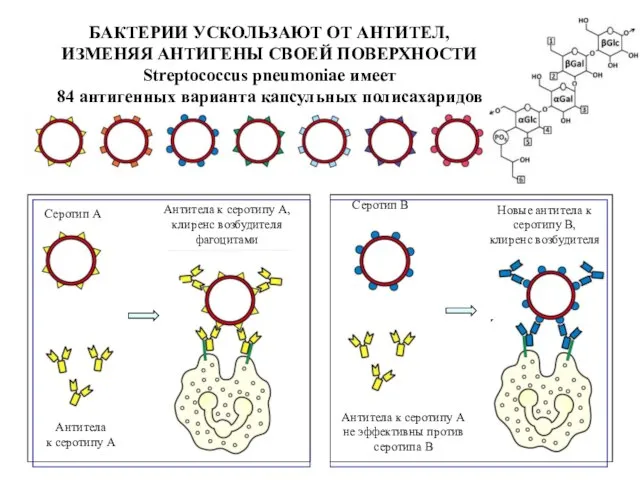
Слайд 55БАКТЕРИИ УСКОЛЬЗАЮТ ОТ АНТИТЕЛ,
ИЗМЕНЯЯ АНТИГЕНЫ СВОЕЙ ПОВЕРХНОСТИ
Streptococcus pneumoniae имеет
84 антигенных варианта капсульных
БАКТЕРИИ УСКОЛЬЗАЮТ ОТ АНТИТЕЛ,
ИЗМЕНЯЯ АНТИГЕНЫ СВОЕЙ ПОВЕРХНОСТИ
Streptococcus pneumoniae имеет
84 антигенных варианта капсульных
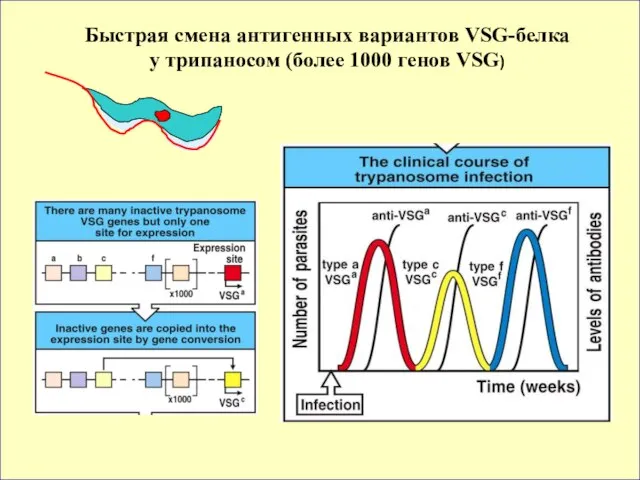
Слайд 56Быстрая смена антигенных вариантов VSG-белка
у трипаносом (более 1000 генов VSG)
Быстрая смена антигенных вариантов VSG-белка
у трипаносом (более 1000 генов VSG)
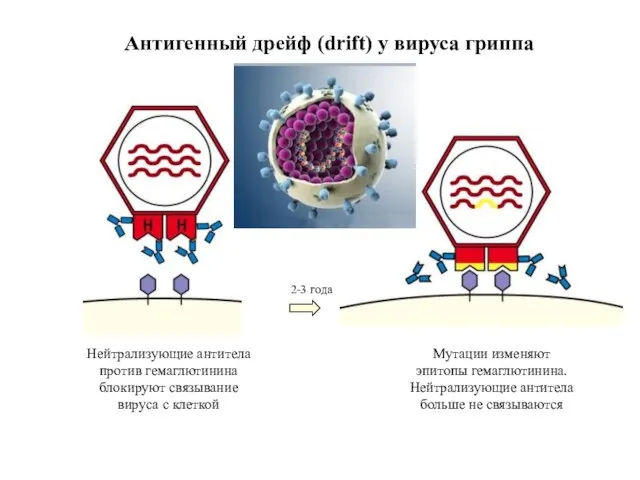
Слайд 57Антигенный дрейф (drift) у вируса гриппа
Нейтрализующие антитела
против гемаглютинина
блокируют связывание
вируса
Антигенный дрейф (drift) у вируса гриппа
Нейтрализующие антитела
против гемаглютинина
блокируют связывание
вируса
Слайд 58Антигенный сдвиг (shift) у вируса гриппа
Обмен сегментами РНК (8)
между штаммами вируса
в
Антигенный сдвиг (shift) у вируса гриппа
Обмен сегментами РНК (8)
между штаммами вируса
в

Слайд 60ИНФЕКЦИЯ И ИММУНИТЕТ
ИСПОЛЬЗУЮТ ИДЕНТИЧНЫЙ ГЕНЕТИЧЕСКИЙ КОНСТРУКТОР
ДЛЯ СОЗДАНИЯ СРЕДСТВ НАПАДЕНИЯ И
ИНФЕКЦИЯ И ИММУНИТЕТ
ИСПОЛЬЗУЮТ ИДЕНТИЧНЫЙ ГЕНЕТИЧЕСКИЙ КОНСТРУКТОР
ДЛЯ СОЗДАНИЯ СРЕДСТВ НАПАДЕНИЯ И

Слайд 61Обе системы – иммунитет и инфекция – очень «интеллектуальны».
Они гибко и адекватно
Обе системы – иммунитет и инфекция – очень «интеллектуальны».
Они гибко и адекватно

Слайд 62Бактерии, как люди, объединяются в коллективы
для достижения своей цели
Бактерии, как люди, объединяются в коллективы
для достижения своей цели

Слайд 63Макроорганизм
БАКТЕРИИ СОБИРАЮТ КВОРУМ,
ДОСТАТОЧНЫЙ ДЛЯ УСПЕШНОГО НАПАДЕНИЯ
Макроорганизм
БАКТЕРИИ СОБИРАЮТ КВОРУМ,
ДОСТАТОЧНЫЙ ДЛЯ УСПЕШНОГО НАПАДЕНИЯ
Слайд 64Синегнойная палочка - условный патоген,
проявляет агрессивные свойства только в поздней логарифмической фазе
Синегнойная палочка - условный патоген,
проявляет агрессивные свойства только в поздней логарифмической фазе
Слайд 65КАК БАКТЕРИИ ОЩУЩАЮТ КВОРУМ
КАК БАКТЕРИИ ОЩУЩАЮТ КВОРУМ

Слайд 66Каждая бактерия умеет «говорить» и «слышать».
Система Quorum sensing молчит при низкой плотности
Система Quorum sensing молчит при низкой плотности
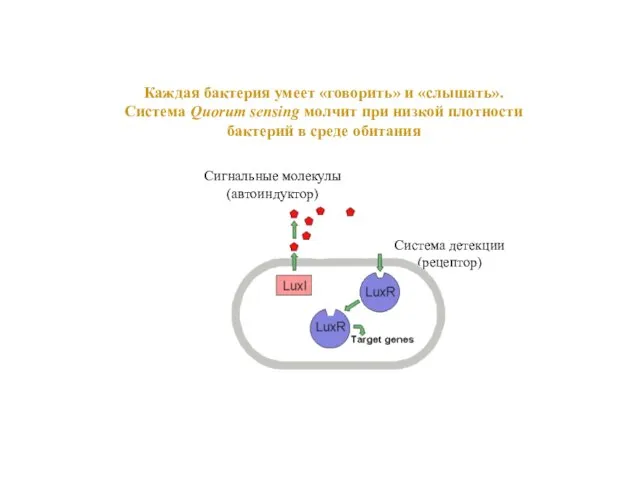
Слайд 67Активация новых генов
продукция факторов вирулентности
Quorum sensing
при определенной плотности бактерий концентрация автоиндуктора
становится
Активация новых генов
продукция факторов вирулентности
Quorum sensing
при определенной плотности бактерий концентрация автоиндуктора
становится
Слайд 68Система Quorum sensing у Грам- и Грам+ бактерий
Система Quorum sensing у Грам- и Грам+ бактерий
Слайд 69Бактериальный язык общения представлен следующими химическими соединениями:
лактоны
хинолы
пептиды
фуранозилдиэфир
бора
Бактериальный язык общения представлен следующими химическими соединениями:
лактоны
хинолы
пептиды
фуранозилдиэфир
бора
Слайд 70 «Механизмы QS позволяют патогенным бактериям очень рационально использовать свой болезнетворный потенциал.
«Механизмы QS позволяют патогенным бактериям очень рационально использовать свой болезнетворный потенциал.

Слайд 71Облигатные внутриклеточные бактерии хламидии, риккетсии, эрлихии
блокируют апоптоз, делая эукариотическую клетку бессмертной
Облигатные внутриклеточные бактерии хламидии, риккетсии, эрлихии
блокируют апоптоз, делая эукариотическую клетку бессмертной
Слайд 72Для манипуляций с нашими клетками бактерии используют молекулярный шприц
(называется «секреторная система»)
Для манипуляций с нашими клетками бактерии используют молекулярный шприц
(называется «секреторная система»)

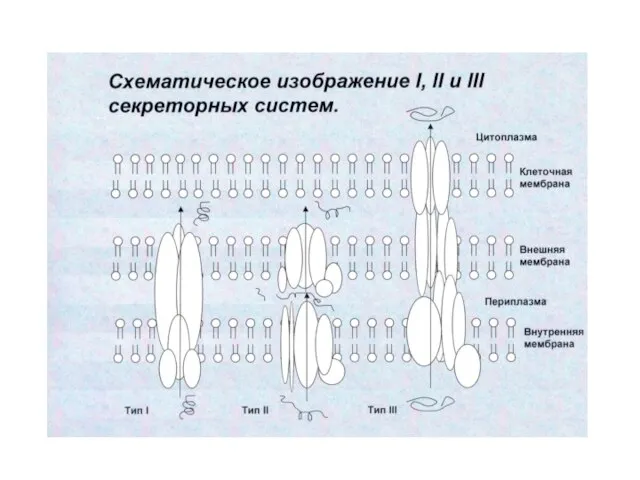
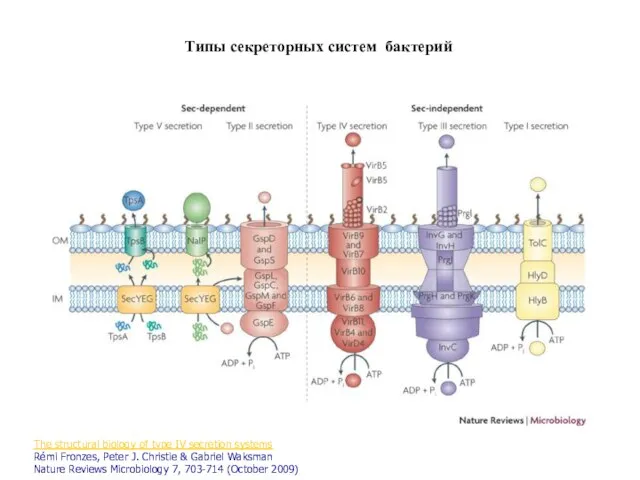
Слайд 74Типы секреторных систем бактерий
The structural biology of type IV secretion systems
Rémi Fronzes,
Типы секреторных систем бактерий
The structural biology of type IV secretion systems
Rémi Fronzes,
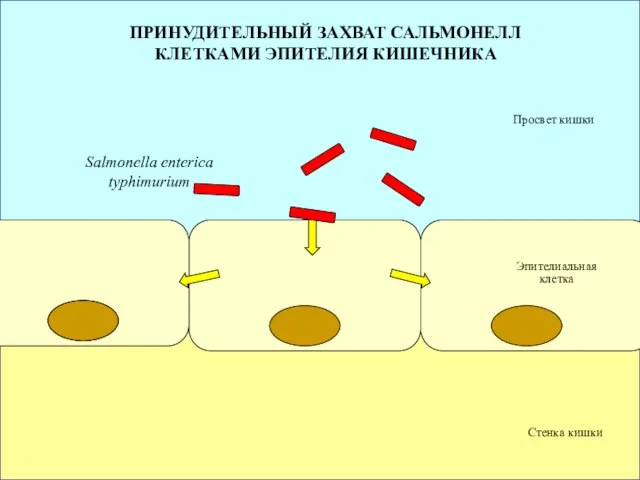
Слайд 75Просвет кишки
Стенка кишки
Salmonella enterica
typhimurium
Эпителиальная
клетка
ПРИНУДИТЕЛЬНЫЙ ЗАХВАТ САЛЬМОНЕЛЛ
КЛЕТКАМИ ЭПИТЕЛИЯ КИШЕЧНИКА
Просвет кишки
Стенка кишки
Salmonella enterica
typhimurium
Эпителиальная
клетка
ПРИНУДИТЕЛЬНЫЙ ЗАХВАТ САЛЬМОНЕЛЛ
КЛЕТКАМИ ЭПИТЕЛИЯ КИШЕЧНИКА
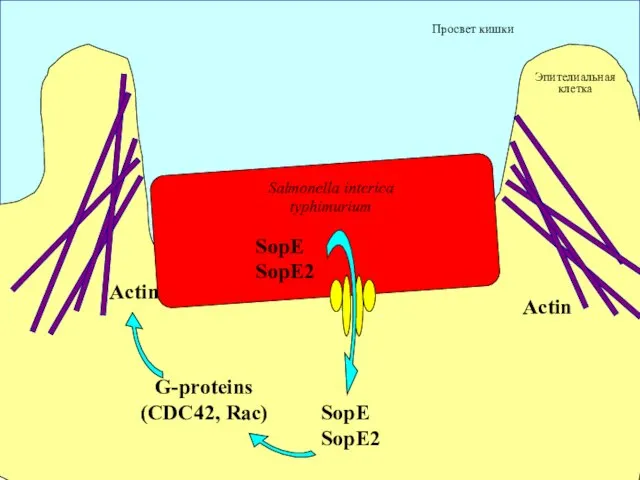
Слайд 76Эпителиальная
клетка
Просвет кишки
SopE
SopE2
Actin
Actin
Salmonella interica
typhimurium
G-proteins
(CDC42, Rac)
SopE
SopE2
Эпителиальная
клетка
Просвет кишки
SopE
SopE2
Actin
Actin
Salmonella interica
typhimurium
G-proteins
(CDC42, Rac)
SopE
SopE2
Слайд 77Макрофаг
Salmonella enterica typhimurium
Shigella flexneri
IpaB
Caspase-1
Бактерия индуцирует апоптоз макрофага
Type III secretory
system
Макрофаг
Salmonella enterica typhimurium
Shigella flexneri
IpaB
Caspase-1
Бактерия индуцирует апоптоз макрофага
Type III secretory
system
Слайд 78Динамическое противостояние двух живых систем
на примере взаимоотношений
вирусной инфекции и иммунитета
Динамическое противостояние двух живых систем
на примере взаимоотношений
вирусной инфекции и иммунитета

Слайд 79Стратегия вируса – спрятаться в наших клетках и стать недоступным
для средств иммунной
Стратегия вируса – спрятаться в наших клетках и стать недоступным
для средств иммунной
Слайд 80MHC-I
Антиген
Т-киллер узнает вирусные
антигены в комплексе
с МНС-I на поверхности
инфицированной клетки
Клетка,
MHC-I
Антиген
Т-киллер узнает вирусные
антигены в комплексе
с МНС-I на поверхности
инфицированной клетки
Клетка,
Слайд 81Клетка, инфицированная
вирусом Эпштейна-Барр
в латентном состоянии
(нет репликации вируса)
Некоторые вирусы нарушают выход вирусных
Клетка, инфицированная
вирусом Эпштейна-Барр
в латентном состоянии
(нет репликации вируса)
Некоторые вирусы нарушают выход вирусных
Слайд 82Вновь синтезированная
молекула МНС-I
Вновь синтезированный
белок вируса
Фрагменты
антигена
Транспортная
везикула
TAP
Клеточная
мембрана
Представление вирусного
Вновь синтезированная
молекула МНС-I
Вновь синтезированный
белок вируса
Фрагменты
антигена
Транспортная
везикула
TAP
Клеточная
мембрана
Представление вирусного
Слайд 83Вновь синтезированная
молекула МНС-I
Вновь синтезированный
белок вируса
Транспортная
везикула
TAP
Клеточная
мембрана
Белок EBNA-1 вируса
Вновь синтезированная
молекула МНС-I
Вновь синтезированный
белок вируса
Транспортная
везикула
TAP
Клеточная
мембрана
Белок EBNA-1 вируса
Слайд 84Клетка, инфицированная
вирусом
Нет МНС-I
на клеточной мембране
Многие вирусы нарушают экспрессию молекул МНС-I
на
Клетка, инфицированная
вирусом
Нет МНС-I
на клеточной мембране
Многие вирусы нарушают экспрессию молекул МНС-I
на
Слайд 85Клетка, инфицированная
вирусом
Нет МНС-I
на клеточной мембране
Если на клетке нет МНС-I,
то
Клетка, инфицированная
вирусом
Нет МНС-I
на клеточной мембране
Если на клетке нет МНС-I,
то
Слайд 86Для блокирования NK-клеток
вирусы производят белки, похожие на МНС-I
(HCMV белок UL18, MCMV
Для блокирования NK-клеток
вирусы производят белки, похожие на МНС-I
(HCMV белок UL18, MCMV
Слайд 87T-киллер
Белок nef вируса иммунодефицита человека
подавляет экспрессию молекул HLA-A и HLA-B,
не затрагивая
T-киллер
Белок nef вируса иммунодефицита человека
подавляет экспрессию молекул HLA-A и HLA-B,
не затрагивая
Слайд 88«ШАХМАТНАЯ ПАРТИЯ»
«ШАХМАТНАЯ ПАРТИЯ»
Слайд 89УРОКИ:
ИММУНИТЕТ И ИНФЕКЦИЯ - ДИНАМИЧНЫЕ И РАЦИОНАЛЬНЫЕ ЖИВЫЕ СИСТЕМЫ, НЕПРЕРЫВНО ИЗМЕНЯЮЩИЕСЯ АДЕКВАТНО
УРОКИ:
ИММУНИТЕТ И ИНФЕКЦИЯ - ДИНАМИЧНЫЕ И РАЦИОНАЛЬНЫЕ ЖИВЫЕ СИСТЕМЫ, НЕПРЕРЫВНО ИЗМЕНЯЮЩИЕСЯ АДЕКВАТНО

Слайд 90ИНФЕКЦИЯ
ИММУНИТЕТ
ШАНСЫ ИММУНИТЕТА И ИНФЕКЦИИ
ПРИМЕРНО РАВНЫ
В ИХ БЕСКОНЕЧНОМ ПРОТИВОСТОЯНИИ
ИНФЕКЦИЯ
ИММУНИТЕТ
ШАНСЫ ИММУНИТЕТА И ИНФЕКЦИИ
ПРИМЕРНО РАВНЫ
В ИХ БЕСКОНЕЧНОМ ПРОТИВОСТОЯНИИ
Слайд 91ИНФЕКЦИЯ
ИММУНИТЕТ
ЭТИОТРОПНОЕ
ЛЕЧЕНИЕ
ДЕЗИНТОКСИКАЦИЯ
ОСЛАБЛЕНИЕ
ИММУНИТЕТА
ВАКЦИНЫ
ИНФЕКЦИЯ
ИММУНИТЕТ
ЭТИОТРОПНОЕ
ЛЕЧЕНИЕ
ДЕЗИНТОКСИКАЦИЯ
ОСЛАБЛЕНИЕ
ИММУНИТЕТА
ВАКЦИНЫ

 Стыд, вина и извинение
Стыд, вина и извинение Фотографические ресурсы Фотокамера изнутри. Необходимый набор техники
Фотографические ресурсы Фотокамера изнутри. Необходимый набор техники Структура и содержание экспертного заключения. Принципы и основания написания экспертного заключения
Структура и содержание экспертного заключения. Принципы и основания написания экспертного заключения Начало реализма. История искусства. Часть 13
Начало реализма. История искусства. Часть 13 Волейбол
Волейбол Власть. Государство. Формы государства. Политическая система
Власть. Государство. Формы государства. Политическая система Счастье наше и счастье наших детей
Счастье наше и счастье наших детей Китайская Народная Республика.
Китайская Народная Республика. Презентация без названия (3)
Презентация без названия (3) Новосибирскте болашақ электростанциясы құрастырылды
Новосибирскте болашақ электростанциясы құрастырылды русская народная сказка
русская народная сказка Innovation incandescent lamp
Innovation incandescent lamp Конкурс «Посылка»!
Конкурс «Посылка»! Участие граждан в политической жизни
Участие граждан в политической жизни №1_Види конструкційних матеріалів. Деревина
№1_Види конструкційних матеріалів. Деревина Aragon. Электрооборудование
Aragon. Электрооборудование ПЛАНИРОВАНИЕ И ПРОГНОЗИРОВАНИЕ В СЕРВИСЕ
ПЛАНИРОВАНИЕ И ПРОГНОЗИРОВАНИЕ В СЕРВИСЕ Кількісний склад розчину. Масова частка розчиненої речовини
Кількісний склад розчину. Масова частка розчиненої речовини Жизнь и быт южноуральцев в послевоенное время.(1946-середина 1960-х гг).
Жизнь и быт южноуральцев в послевоенное время.(1946-середина 1960-х гг). Задание А17
Задание А17 Международное патриотическое движение за здоровый образ жизни. Русские пробежки
Международное патриотическое движение за здоровый образ жизни. Русские пробежки Творческий проект по технологии
Творческий проект по технологии Папирус
Папирус День КОНСТИТУЦИИ
День КОНСТИТУЦИИ Обобщающий урок
Обобщающий урок Безопасность при чрезвычайных ситуациях природного характера
Безопасность при чрезвычайных ситуациях природного характера Достижения Н.И.Вавилова в селекции растений
Достижения Н.И.Вавилова в селекции растений Психология творческой деятельности. Лекция 3. Творческий процесс
Психология творческой деятельности. Лекция 3. Творческий процесс