- Классификация и механизм действия гормонов

Содержание
- 2. 4 основные системы регуляции метаболизма: Центральная нервная система (за счет передачи сигналов посредством нервных импульсов и
- 3. Эндокринная система регулирует обмен веществ посредством гормонов. Гормоны (др.-греч. ὁρμάω — возбуждаю, побуждаю) - биологически активные
- 4. Классическим гормонам присущ ряд признаков: Дистантность действия – синтез в железах внутренней секреции, а регуляция отдаленных
- 5. Организация нервно-гормональной регуляции Существует строгая иерархия или соподчиненность гормонов. Поддержание уровня гормонов в организме в большинстве
- 8. Регуляция уровня гормонов в организме Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет
- 9. Классификация гормонов по биологическим функциям; по механизму действия; по химическому строению; различают 4 группы: 1. Белково-пептидные
- 10. Классификация гормонов по биологическим функциям.
- 11. Классификация гормонов по химическому строению
- 12. 1. Белково - пептидные гормоны Гормоны гипоталамуса; гормоны гипофиза; гормоны поджелудочной железы - инсулин, глюкагон; гормоны
- 13. Главный анаболический гормон – инсулин, главный катаболический гормон - глюкагон
- 14. Некоторые представители белково - пептидных гормонов: тиролиберина (пироглу-гис-про-NН2), инсулина и соматостатина.
- 15. 2. Гормоны - производные аминокислот Являются производными аминокислоты - тирозина. К ним относятся гормоны щитовидной железы
- 16. Гормоны щитовидной железы
- 17. Схема синтеза трийодтиронинов
- 18. 3. Гормоны стероидной природы Синтезируются из холестерина (на рис.) Гормоны коркового вещества надпочечников – кортикостероиды (кортизол,
- 19. Синтез основных кортикостероидов
- 20. Эйкозаноиды Предшественником всех эйкозаноидов является арахидоновая кислота. Они делятся на 3 группы – простагландины, лейкотриены, тромбоксаны.
- 21. Характеристика разных групп эйказоноидов Простагландины (Pg) — синтезируются практически во всех клетках, кроме эритроцитов и лимфоцитов.
- 22. Тромбоксаны и лейкотриены Тромбоксаны (Tx) образуются в тромбоцитах, стимулируют их агрегацию и вызывают сужение мелких сосудов.
- 23. Взаимодействие гормонов с рецепторами клеток-мишеней Для проявления биологической активности связывание гормонов с рецепторами должно приводить к
- 25. Общая характеристика рецепторов Рецепторы могут находится: - на поверхности клеточной мембраны - внутри клетки – в
- 26. Основные этапы передачи гормонального сигнала: через мембранные (гидрофобные) и внутриклеточные (гидрофильные) рецепторы. Это быстрый и медленный
- 27. Гормональный сигнал меняет скорость метаболических процессов ответ путем: - изменение активности ферментов - изменение количества ферментов.
- 28. Передача гормонального сигнала через внутриклеточные рецепторы для стероидных гормонов (гормоны коры надпочечников и половые гормоны), тиреодных
- 29. Передача гормонального сигнала через ядерный рецептор.
- 30. Передача гормонального сигнала через мембранные рецепторы Передача информации от первичного посредника гормона осуществляется через рецептор. Этот
- 31. Сигнал от гормона трансформируется в изменении концентрации вторичных посредников – цАМФ, цГТФ, ИФ3, ДАГ, СА2+, NO.
- 32. Самая распространенная система передача гормонального сигнала через мембранные рецепторы – аденилатциклазная система. Комплекс гормон-рецептор связан с
- 33. Адреналин и глюкагон через аденилатциклазную систему передачи гормонального сигнала активируют гормонзависимую ТАГ-липазу адипоцитов. Происходит при напряжении
- 34. Передача сигнала с адренорецепторов. АС – аденилатциклаза, PkA – протеинкиназа А, PkC – протеинкиназа С, ФлС
- 35. Адренорецепторы всех типов реализуют свое действие через Gs-белки. α- субъединицы этого белка активируют аденилатциклазу, которая обеспечивает
- 36. Американские ученые Роберт Лефковиц и Брайан Кобилка удостоились Нобелевской премии в 2012 г. за постижение механизмов
- 37. G-белки (англ. G proteins) — это семейство белков, относящихся к ГТФазам и функционирующих в качестве посредников
- 38. G-белки делятся на две основных группы — гетеротримерные («большие») и «малые». Гетеротримерные G-белки — это белки
- 39. Циклический аденозинмонофосфат (циклический AMФ, цAMФ, cAMP) — производное АТФ, выполняющее в организме роль вторичного посредника, использующегося
- 40. Каждой из систем передачи гормонального сигнала соответствует определенный класс протеинкиназ Активность протеинкиназ типа А регулируется цАМФ,
- 41. Действие инсулина на клетки-мишени начинается после его связывания с мембранными рецепторами, при этом внутриклеточный домен рецептора
- 42. Другая система – гуанилатциклазная мессенджерская система. Цитоплазматический домен рецептора обладает активностью гуанилатциклазы (гемсодержащий фермент). Молекулы цГТФ
- 43. Передача гормонального сигнала через NO
- 44. Инозитолфосфатная система. Связывание гормона с рецептором, вызывает изменение конформациии рецептора. Происходит диссоциация G-белка и ГДФ заменяется
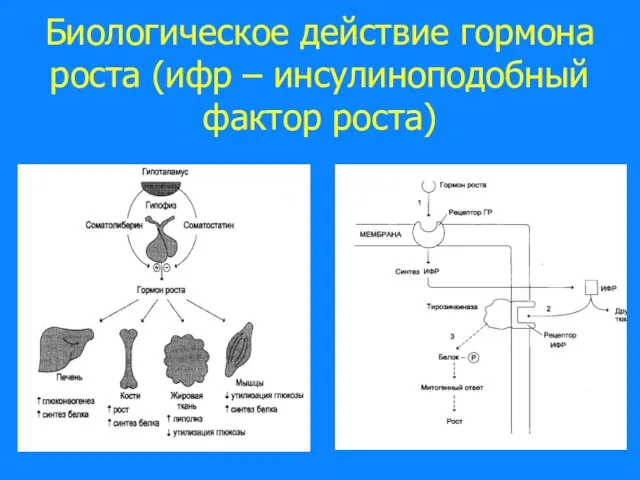
- 45. Биологическое действие гормона роста (ифр – инсулиноподобный фактор роста)
- 47. Скачать презентацию
Слайд 24 основные системы регуляции метаболизма:
Центральная нервная система (за счет передачи сигналов посредством
4 основные системы регуляции метаболизма:
Центральная нервная система (за счет передачи сигналов посредством

Слайд 3Эндокринная система регулирует обмен веществ посредством гормонов.
Гормоны (др.-греч. ὁρμάω — возбуждаю, побуждаю)
Эндокринная система регулирует обмен веществ посредством гормонов. Гормоны (др.-греч. ὁρμάω — возбуждаю, побуждаю)

Слайд 4Классическим гормонам присущ ряд признаков:
Дистантность действия – синтез в железах внутренней секреции,
Классическим гормонам присущ ряд признаков:
Дистантность действия – синтез в железах внутренней секреции,

Слайд 5Организация нервно-гормональной регуляции
Существует строгая иерархия или соподчиненность гормонов.
Поддержание уровня гормонов в организме
Организация нервно-гормональной регуляции
Существует строгая иерархия или соподчиненность гормонов.
Поддержание уровня гормонов в организме

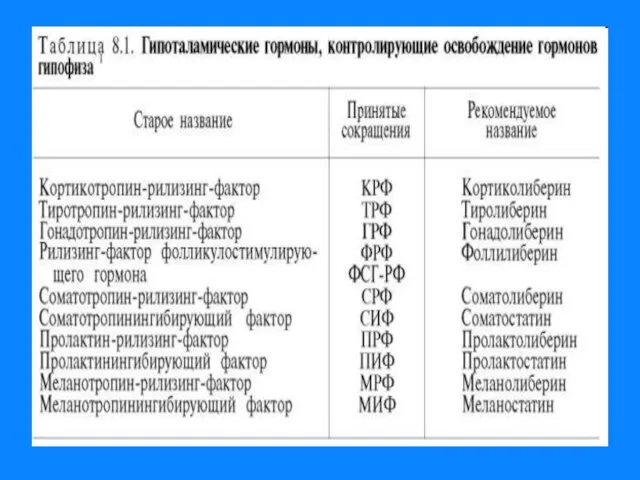
Слайд 8Регуляция уровня гормонов в организме
Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной
Регуляция уровня гормонов в организме
Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной

Слайд 9Классификация гормонов
по биологическим функциям;
по механизму действия;
по химическому строению;
различают 4 группы:
1.
Классификация гормонов
по биологическим функциям;
по механизму действия;
по химическому строению;
различают 4 группы:
1.

Слайд 10Классификация гормонов по биологическим функциям.
Классификация гормонов по биологическим функциям.

Слайд 11Классификация гормонов по химическому строению
Классификация гормонов по химическому строению

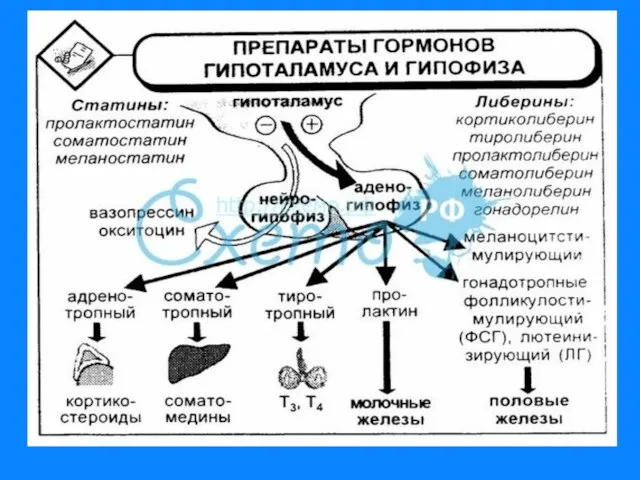
Слайд 121. Белково - пептидные
гормоны
Гормоны гипоталамуса; гормоны гипофиза; гормоны поджелудочной железы - инсулин,
1. Белково - пептидные
гормоны
Гормоны гипоталамуса; гормоны гипофиза; гормоны поджелудочной железы - инсулин,

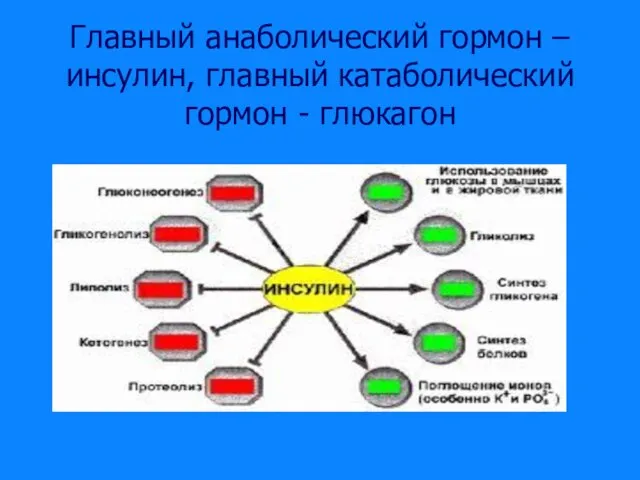
Слайд 13Главный анаболический гормон – инсулин, главный катаболический гормон - глюкагон
Главный анаболический гормон – инсулин, главный катаболический гормон - глюкагон
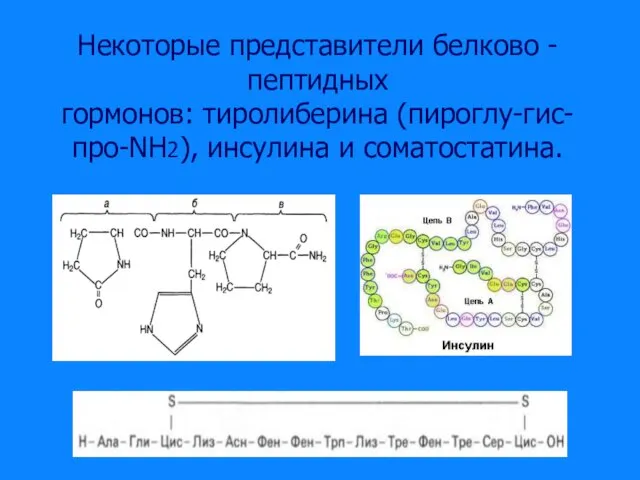
Слайд 14Некоторые представители белково - пептидных
гормонов: тиролиберина (пироглу-гис-про-NН2), инсулина и соматостатина.
Некоторые представители белково - пептидных
гормонов: тиролиберина (пироглу-гис-про-NН2), инсулина и соматостатина.
Слайд 152. Гормоны - производные аминокислот
Являются производными аминокислоты -
тирозина.
К ним относятся
2. Гормоны - производные аминокислот
Являются производными аминокислоты -
тирозина.
К ним относятся
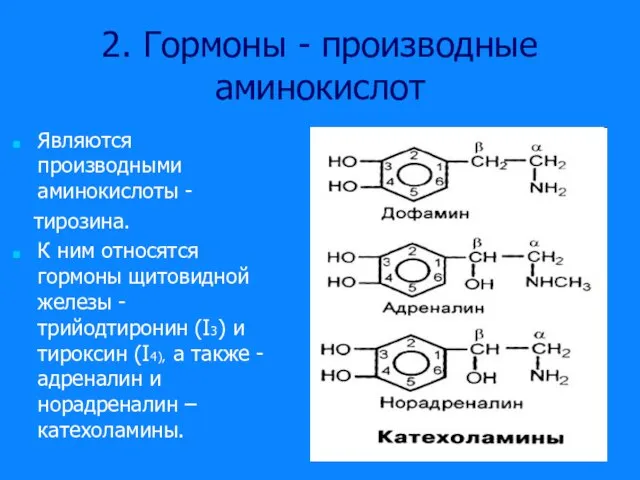
Слайд 16Гормоны щитовидной железы
Гормоны щитовидной железы
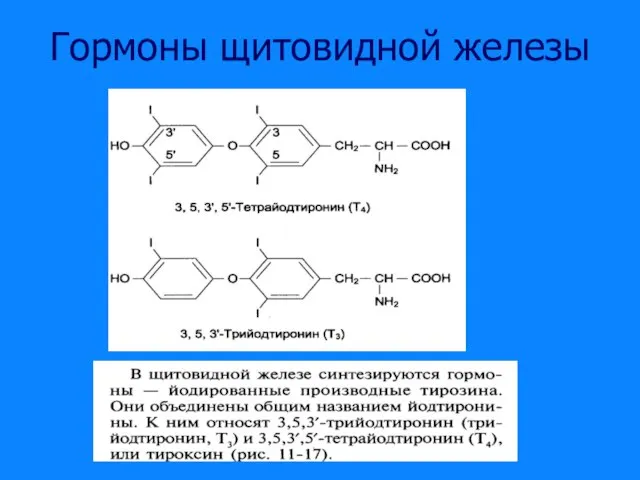
Слайд 17Схема синтеза трийодтиронинов
Схема синтеза трийодтиронинов

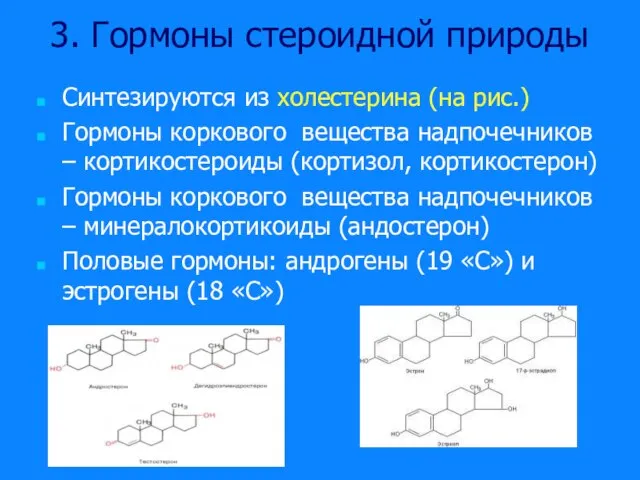
Слайд 183. Гормоны стероидной природы
Синтезируются из холестерина (на рис.)
Гормоны коркового вещества надпочечников –
3. Гормоны стероидной природы
Синтезируются из холестерина (на рис.)
Гормоны коркового вещества надпочечников –
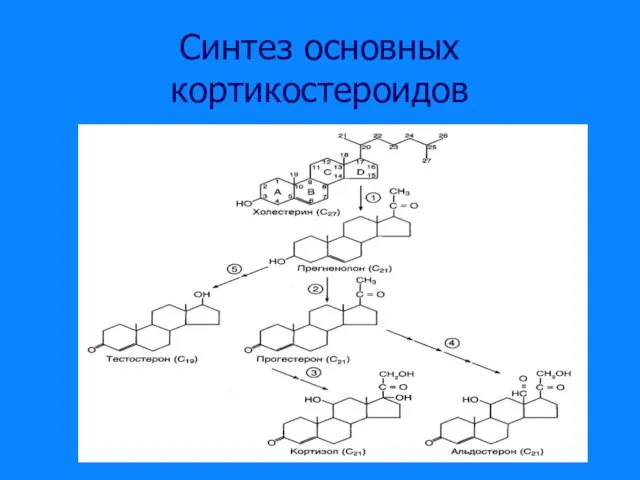
Слайд 19Синтез основных кортикостероидов
Синтез основных кортикостероидов
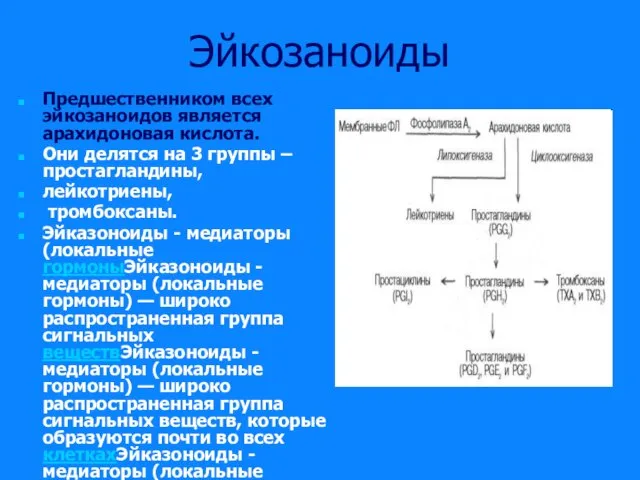
Слайд 20Эйкозаноиды
Предшественником всех эйкозаноидов является арахидоновая кислота.
Они делятся на 3 группы –
Эйкозаноиды
Предшественником всех эйкозаноидов является арахидоновая кислота.
Они делятся на 3 группы –
Слайд 21Характеристика разных групп эйказоноидов
Простагландины (Pg) — синтезируются практически во всех клетках, кроме эритроцитов
Характеристика разных групп эйказоноидов
Простагландины (Pg) — синтезируются практически во всех клетках, кроме эритроцитов

Слайд 22Тромбоксаны и лейкотриены
Тромбоксаны (Tx) образуются в тромбоцитах, стимулируют их агрегацию и вызывают
Тромбоксаны и лейкотриены
Тромбоксаны (Tx) образуются в тромбоцитах, стимулируют их агрегацию и вызывают

Слайд 23Взаимодействие гормонов с рецепторами клеток-мишеней
Для проявления биологической активности связывание гормонов с рецепторами
Взаимодействие гормонов с рецепторами клеток-мишеней
Для проявления биологической активности связывание гормонов с рецепторами


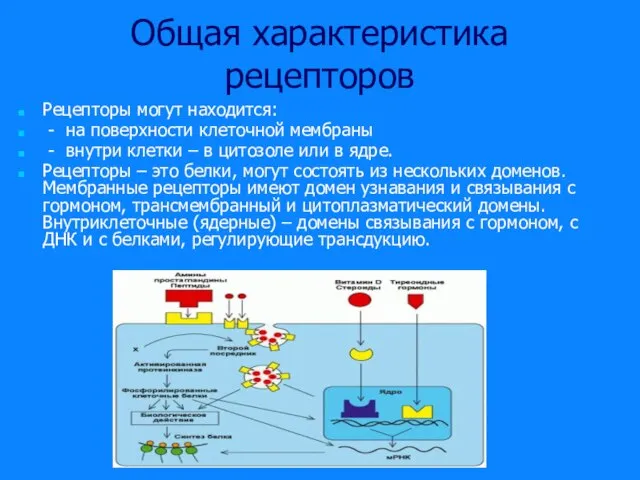
Слайд 25Общая характеристика рецепторов
Рецепторы могут находится:
- на поверхности клеточной мембраны
- внутри
Общая характеристика рецепторов
Рецепторы могут находится:
- на поверхности клеточной мембраны
- внутри
Слайд 26Основные этапы передачи гормонального сигнала:
через мембранные (гидрофобные) и внутриклеточные (гидрофильные) рецепторы. Это
Основные этапы передачи гормонального сигнала: через мембранные (гидрофобные) и внутриклеточные (гидрофильные) рецепторы. Это

Слайд 27Гормональный сигнал меняет скорость метаболических процессов ответ путем:
- изменение активности ферментов
Гормональный сигнал меняет скорость метаболических процессов ответ путем: - изменение активности ферментов

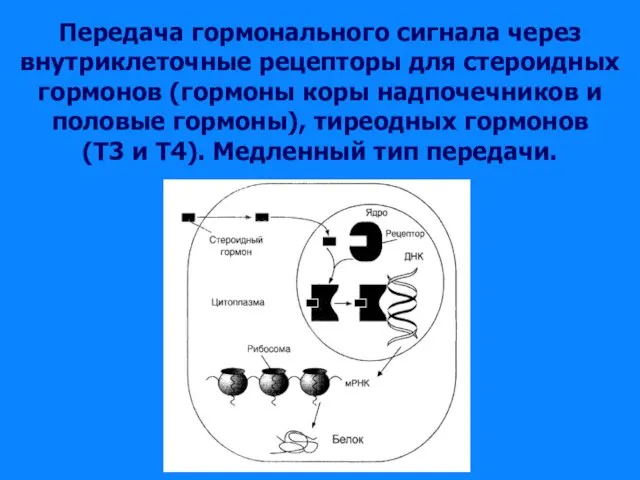
Слайд 28Передача гормонального сигнала через
внутриклеточные рецепторы для стероидных гормонов (гормоны коры надпочечников
Передача гормонального сигнала через внутриклеточные рецепторы для стероидных гормонов (гормоны коры надпочечников
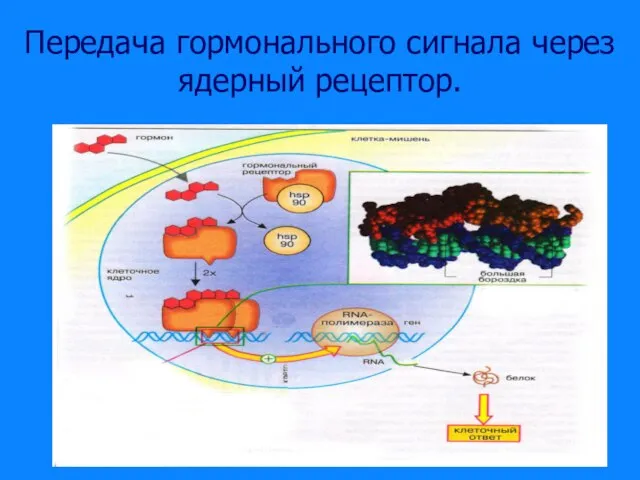
Слайд 29Передача гормонального сигнала через ядерный рецептор.
Передача гормонального сигнала через ядерный рецептор.
Слайд 30Передача гормонального сигнала через мембранные рецепторы
Передача информации от первичного посредника гормона осуществляется
Передача гормонального сигнала через мембранные рецепторы
Передача информации от первичного посредника гормона осуществляется

Слайд 31Сигнал от гормона трансформируется в изменении концентрации вторичных посредников – цАМФ, цГТФ,
Сигнал от гормона трансформируется в изменении концентрации вторичных посредников – цАМФ, цГТФ,
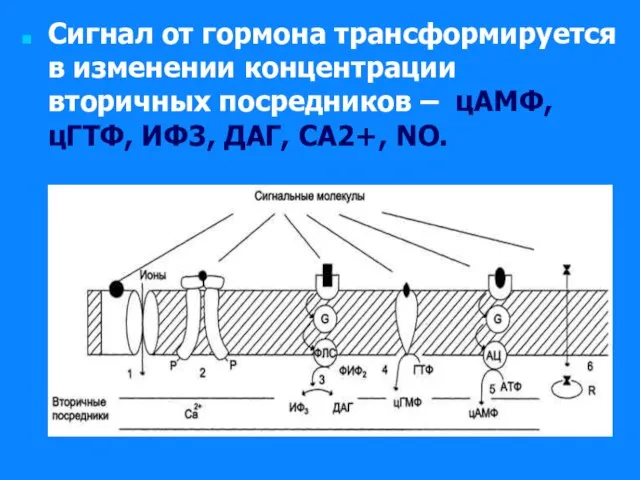
Слайд 32Самая распространенная система передача гормонального сигнала через мембранные рецепторы – аденилатциклазная система.
Комплекс
Самая распространенная система передача гормонального сигнала через мембранные рецепторы – аденилатциклазная система.
Комплекс

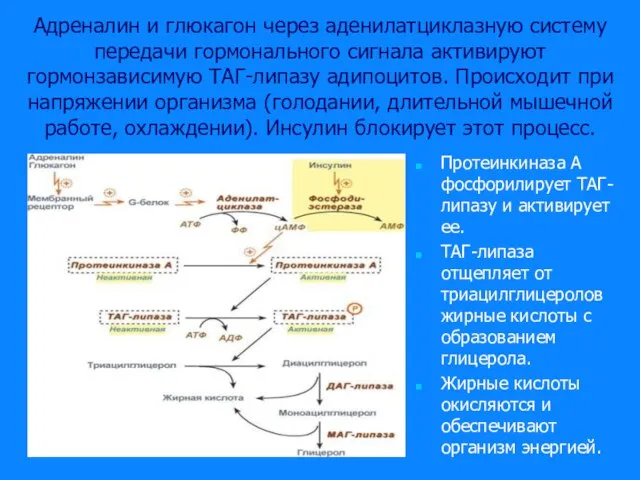
Слайд 33Адреналин и глюкагон через аденилатциклазную систему передачи гормонального сигнала активируют гормонзависимую ТАГ-липазу
Адреналин и глюкагон через аденилатциклазную систему передачи гормонального сигнала активируют гормонзависимую ТАГ-липазу
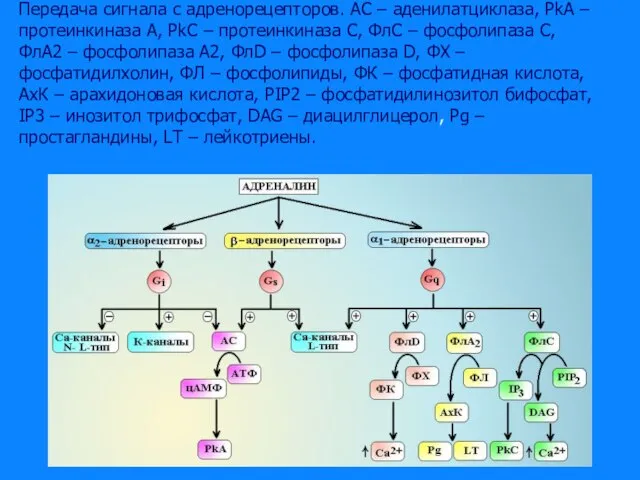
Слайд 34Передача сигнала с адренорецепторов. АС – аденилатциклаза, PkA – протеинкиназа А, PkC
Передача сигнала с адренорецепторов. АС – аденилатциклаза, PkA – протеинкиназа А, PkC
Слайд 35Адренорецепторы всех типов реализуют свое действие через Gs-белки. α- субъединицы этого белка
Адренорецепторы всех типов реализуют свое действие через Gs-белки. α- субъединицы этого белка

Слайд 36Американские ученые Роберт Лефковиц и Брайан Кобилка удостоились Нобелевской премии в 2012
Американские ученые Роберт Лефковиц и Брайан Кобилка удостоились Нобелевской премии в 2012

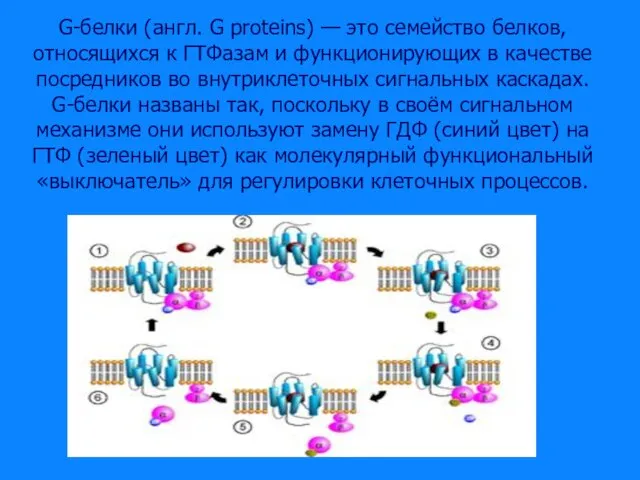
Слайд 37G-белки (англ. G proteins) — это семейство белков, относящихся к ГТФазам и
G-белки (англ. G proteins) — это семейство белков, относящихся к ГТФазам и
Слайд 38G-белки делятся на две основных группы — гетеротримерные («большие») и «малые». Гетеротримерные
G-белки делятся на две основных группы — гетеротримерные («большие») и «малые». Гетеротримерные

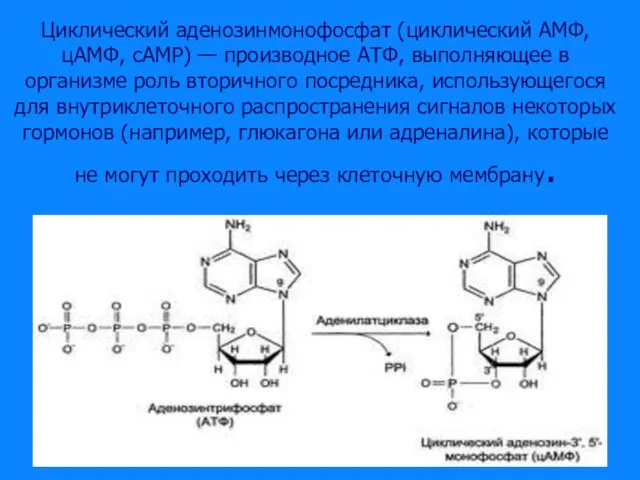
Слайд 39Циклический аденозинмонофосфат (циклический AMФ, цAMФ, cAMP) — производное АТФ, выполняющее в организме
Циклический аденозинмонофосфат (циклический AMФ, цAMФ, cAMP) — производное АТФ, выполняющее в организме
Слайд 40Каждой из систем передачи гормонального сигнала соответствует определенный класс протеинкиназ
Активность протеинкиназ типа
Каждой из систем передачи гормонального сигнала соответствует определенный класс протеинкиназ
Активность протеинкиназ типа

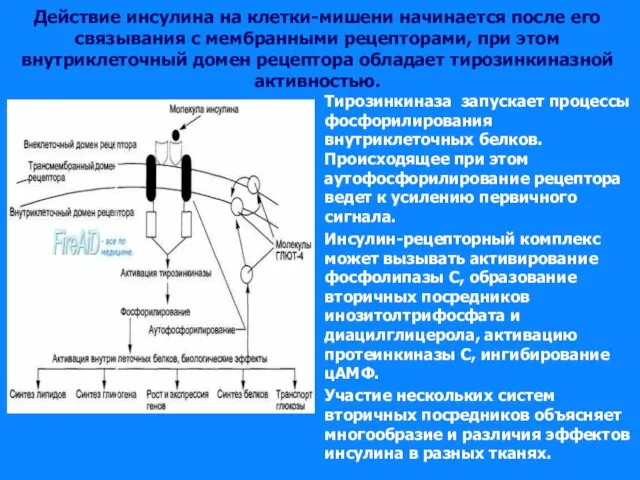
Слайд 41Действие инсулина на клетки-мишени начинается после его связывания с мембранными рецепторами, при
Действие инсулина на клетки-мишени начинается после его связывания с мембранными рецепторами, при
Слайд 42Другая система – гуанилатциклазная мессенджерская система.
Цитоплазматический домен рецептора обладает активностью гуанилатциклазы (гемсодержащий
Другая система – гуанилатциклазная мессенджерская система.
Цитоплазматический домен рецептора обладает активностью гуанилатциклазы (гемсодержащий

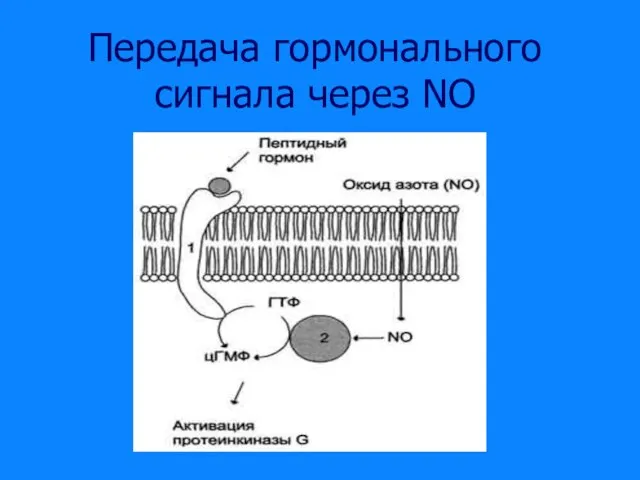
Слайд 43Передача гормонального сигнала через NO
Передача гормонального сигнала через NO
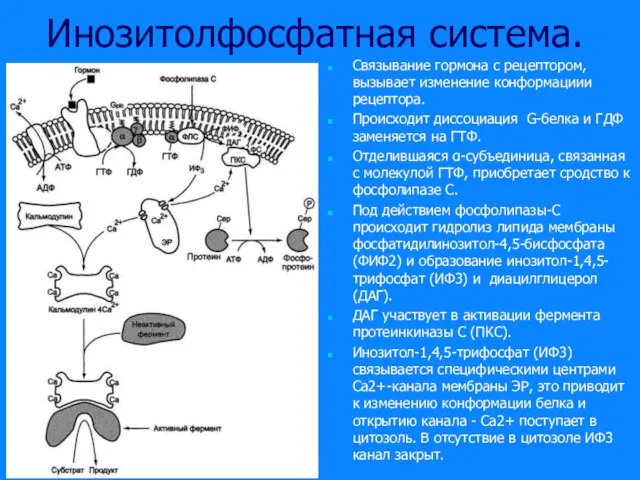
Слайд 44Инозитолфосфатная система.
Связывание гормона с рецептором, вызывает изменение конформациии рецептора.
Происходит диссоциация G-белка и
Инозитолфосфатная система.
Связывание гормона с рецептором, вызывает изменение конформациии рецептора.
Происходит диссоциация G-белка и
Слайд 45Биологическое действие гормона роста (ифр – инсулиноподобный фактор роста)
Биологическое действие гормона роста (ифр – инсулиноподобный фактор роста)
 Тема Доклада: "Об исполнении консолидированного бюджета Волгоградской области за первое полугодие 2009 года (итоги, проблемы, пути р
Тема Доклада: "Об исполнении консолидированного бюджета Волгоградской области за первое полугодие 2009 года (итоги, проблемы, пути р Какая почва в республике Мордовия? (4класс)
Какая почва в республике Мордовия? (4класс) Избирательное право
Избирательное право Исторический футбол «Мы – славяне» (Игра для учащихся 6х классов)
Исторический футбол «Мы – славяне» (Игра для учащихся 6х классов) Год науки и технологий
Год науки и технологий Лови момент, или, как не попасть в ловушку
Лови момент, или, как не попасть в ловушку Методические положения технико-экономической оценки мероприятий по реконструкции электрических сетей
Методические положения технико-экономической оценки мероприятий по реконструкции электрических сетей Действие ионизирующих излучений на организм человека. Профилактика радиационных поражений.
Действие ионизирующих излучений на организм человека. Профилактика радиационных поражений. Налоги и налогообложение
Налоги и налогообложение Автономный робот-манипулятор
Автономный робот-манипулятор Обобщение изученного за курс 7 класса
Обобщение изученного за курс 7 класса Презентация Збройні Сили всеосвіта
Презентация Збройні Сили всеосвіта Славьте Бога Господи, как странно, что я жил, Тебя не зная, жил без милости Твоей святой и без Твоей любви, но ныне я с Тобой, меня в
Славьте Бога Господи, как странно, что я жил, Тебя не зная, жил без милости Твоей святой и без Твоей любви, но ныне я с Тобой, меня в  Налоговое законодательство Латвии
Налоговое законодательство Латвии Ведение успешного аптечного бизнеса
Ведение успешного аптечного бизнеса ПРОБЛЕМЫ ПРИМЕНЕНИЯ МЕЖДУНАРОДНЫХ СТАНДАРТОВ В РАСПРЕДЕЛИТЕЛЬНОМ ЭЛЕКТРОСЕТЕВОМ КОМПЛЕКСЕ
ПРОБЛЕМЫ ПРИМЕНЕНИЯ МЕЖДУНАРОДНЫХ СТАНДАРТОВ В РАСПРЕДЕЛИТЕЛЬНОМ ЭЛЕКТРОСЕТЕВОМ КОМПЛЕКСЕ Строение насекомого
Строение насекомого Класифікація хімічних реакцій за кількістю і складом реагентів та продуктів реакцій
Класифікація хімічних реакцій за кількістю і складом реагентів та продуктів реакцій Долг и совесть
Долг и совесть Источники информации для журналистов деловых СМИ
Источники информации для журналистов деловых СМИ Определение перевода и виды перевода
Определение перевода и виды перевода Обручев Владимир Афанасьевич (1863-1956 гг.)
Обручев Владимир Афанасьевич (1863-1956 гг.) Мастерская Умелец
Мастерская Умелец ПРЕЗЕНТАЦИЯ ПЕДАГОГИЧЕСКОГО ОПЫТАучителя английского языкаПершотравенской общеобразовательной школы І – ІІІ ступеней №1КОЛЕ
ПРЕЗЕНТАЦИЯ ПЕДАГОГИЧЕСКОГО ОПЫТАучителя английского языкаПершотравенской общеобразовательной школы І – ІІІ ступеней №1КОЛЕ Маркетинг – это... М А Р К Е Т И Н Г Олег Фельдман, Генеральный директор КОМКОН-Фарма
Маркетинг – это... М А Р К Е Т И Н Г Олег Фельдман, Генеральный директор КОМКОН-Фарма Лихтенштейн
Лихтенштейн Мир моих кумиров
Мир моих кумиров Офис в облаках
Офис в облаках