- Липиды

Содержание
- 2. КЛАССИФИКАЦИЯ ЛИПИДОВ В зависимости от состава, строения и роли в организме сложилась следующая классификация липидов. 1.
- 3. 2. Сложные липиды. К ним относятся фосфолипиды и гликолипиды. Они состоят из остатков ВЖК, глицерина (или
- 4. 3. Отдельно следует выделить стериды - сложные эфиры ВЖК и полициклических спиртов - стеролов. Кроме того
- 5. РОЛЬ ЛИПИДОВ В ОРГАНИЗМЕ 1) Являются компонентами мембран. В мембране обладают не только структурной, но и
- 6. . 4) Могут быть предшественниками других важных соединений (простагландинов, простациклинов, тромбоксана), которые являются регуляторами многих важных
- 7. 7) Липиды влияют на иммунные реакции организма, усиливают реакции иммунитета. 8) С помощью липидов в организм
- 8. ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЛИПИДОВ В организм липиды поступают с жирной растительной, и особенно, с животной пищей.
- 9. НЖ при гидролизе распадается на глицерин и ВЖК под влиянием липазы поджелудочной железы. Для работы этого
- 10. В желчи человека преобладают холевая, и хенодезоксихолевая кислоты. Причем они находятся в виде парных или конъюгированных
- 11. Соли Жк, будучи поверхностно активными веществами 1) эмульгируют частицы жира и поэтому облегчают действие липолитических ферментов.
- 12. СН2-0-COR CH2OH CH-O-COR + 3H2O СHOH + 3RCOOH CH2-O-COR CH2OH Происходит гидролиз эфирных связей с присоединением
- 13. Жирные кислоты не растворимы в воде, поэтому для их всасывания необходимо образование мицелл. В состав мицелл
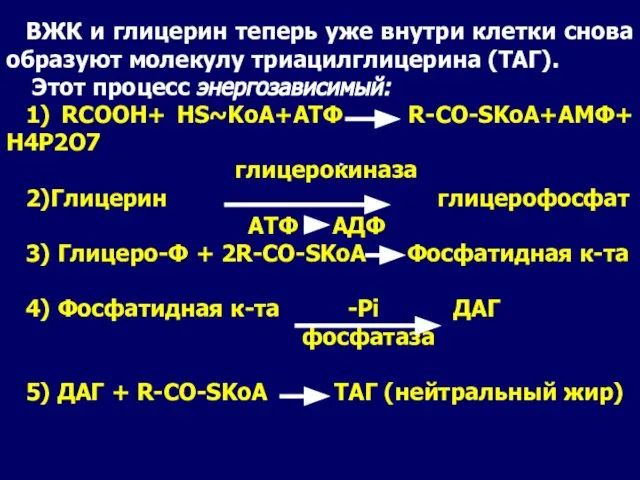
- 14. ВЖК и глицерин теперь уже внутри клетки снова образуют молекулу триацилглицерина (ТАГ). Этот процесс энергозависимый: 1)
- 15. Жиры в водной среде крови не могут находиться в свободном состоянии, т.к. они не растворимы в
- 16. КЛАССИФИКАЦИЯ ЛИПОПРОТЕИДОВ КРОВИ Липопротеиды называются по плотности или по положению, которое занимают на электрофореграмме: ХМ (Хиломикроны)
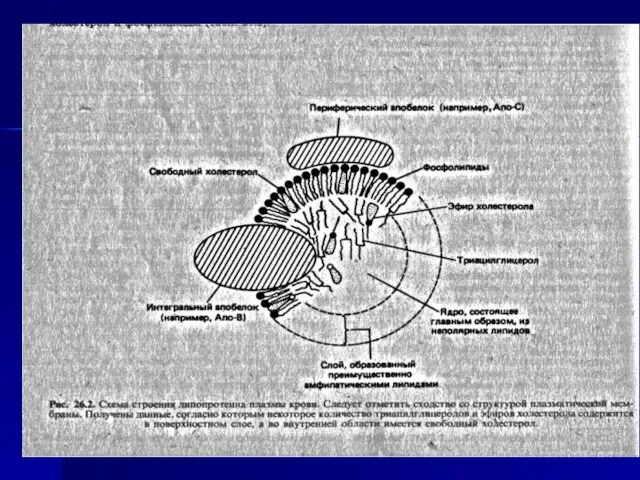
- 17. В состав липопротеидов входят белки (апопротеины), фосфолипиды, нейтральные жиры, эфиры холестерина и свободный холестерин. Снаружи располагаются
- 19. ХИЛОМИКРОНЫ В энтероцитах образуются хиломикроны, которые на 80% состоят из НЖ. ХМ попадают не в кровь,
- 20. На поверхности эндотелиальных клеток, выстилающих капилляры, имеется фермент липопротеид-липаза. ЛП-липаза неактивная. Активация происходит под влиянием гепарина,
- 21. После отделения триацилглицеридов ХМ превращаются в хилоремнанты ("ремнанты" - остатки), которые поступают в печень. В печени
- 22. ЛИПОПРОТЕИДЫ ОЧЕНЬ НИЗКОЙ ПЛОТНОСТИ ЛПОНП образуются в печени. Белка 10%, липидов - 90% (55% - НЖ.
- 23. ЛПОНП после их образования в печеночных клетках выносятся в кровеносное русло и разносятся с током крови
- 24. ЛИПОПРОТЕИДЫ НИЗКОЙ ПЛОТНОСТИ ЛПНП - образуются из ЛПОНП частично в печени, частично непосредственно в плазме крови.
- 25. Около 50% ЛПНП распадается в печени, другая половина - во внепеченочных тканях. Разрушение ЛП в клетках
- 26. ЛИПОПРОТЕИДЫ ВЫСОКОЙ ПЛОТНОСТИ ЛПВП – синтезируются в печени, но окончательно формируются в крови. В них белка
- 27. На поверхности ЛПВП расположены ФЛ - лецитины. Фермент ЛХАТ (лецитин-холестерин-ацил-трансфераза) от лецитина отнимает жирную кислоту и
- 28. П Е Р Е Р Ы В
- 29. Концентрация ЛП в крови меняется в зависимости от: 1) приема пищи, 2) от активности ЛП-липаз 3)
- 30. Нарушения липопротеидного обмена бывают наследственные, но нередки и при некоторых других заболеваниях, как сахарный диабет, алкоголизм,
- 31. Типы гиперлипопротеидемий: I тип: повышена концентрация ХМ. Наблюдается при наследственной недостаточности ЛП-липазы. В отличие от здоровых
- 32. II тип: повышено содержание ЛПНП или суммарной фракции ЛПОНП + ЛПНП. Причина заключается в недостатке или
- 33. II тип: повышено содержание ЛПНП или суммарной фракции ЛПОНП + ЛПНП. Причина заключается в недостатке или
- 34. II тип: повышено содержание ЛПНП или суммарной фракции ЛПОНП + ЛПНП. Причина заключается в недостатке или
- 35. III тип: нарушается превращение ЛПОНП в ЛПНП. В крови аномальные пре-бета-ЛП и ХМ. Проявляется это заболевание
- 36. IV тип. В крови повышено содержание ЛПОНП, Х, ТАГ. Это обычно бывает у больных сахарным диабетом
- 37. V тип: повышено количество ХМ и ЛПОНП. Концентрация ЛПНП и ЛПВП обычно снижена. Это редкая форма.
- 38. ОБМЕН НЕЙТРАЛЬНОГО ЖИРА НЖ состоит из смеси ТАГ (95%), ДАГ (3%) и МАГ (2%). ВЖК могут
- 39. СИНТЕЗ НЕЙТРАЛЬНОГО ЖИРА RCOOH + HS~KoA + ATФ R-CO-SKoA + AMФ + Н4Р2О7 2)Глицерин глицерокиназа глицеро-Ф
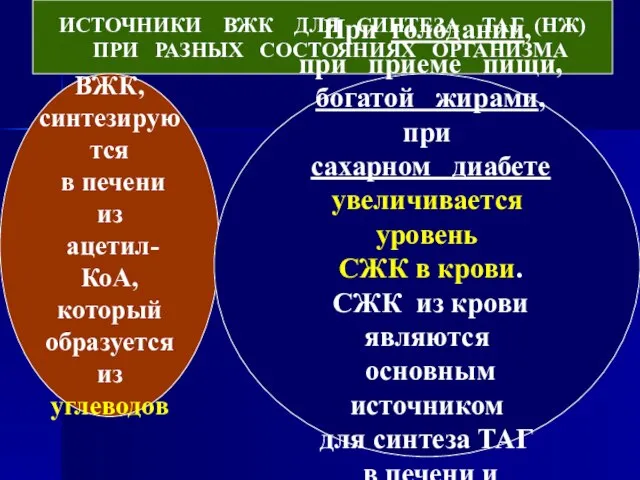
- 40. ИСТОЧНИКИ ВЖК ДЛЯ СИНТЕЗА ТАГ (НЖ) ПРИ РАЗНЫХ СОСТОЯНИЯХ ОРГАНИЗМА ВЖК, синтезируются в печени из ацетил-КоА,
- 41. В норме ТАГ не накапливаются в печени. Сразу же после образования они соединяются с ЛПОНП и
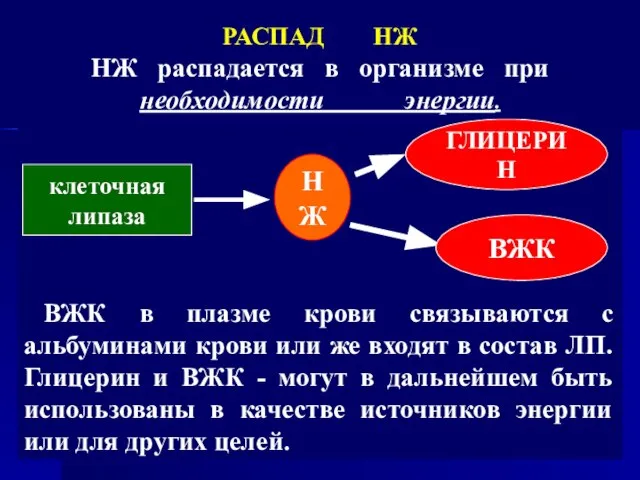
- 42. ВЖК в плазме крови связываются с альбуминами крови или же входят в состав ЛП. Глицерин и
- 43. БЕТА-ОКИСЛЕНИЕ ВЖК Бета-окисление ВЖК осуществляется для получения энергии. ВЖК освобождаются при липолизе нейтрального жира ВЖК окисляются
- 44. Фермент ацил-КоА-синтетаза (тиокиназа) в присутствии АТФ и КоА превращает СЖК в активную ЖК или ацил-КоА: ВЖК+АТФ+КоА
- 45. На наружной стороне Мх-мембраны происходит следующая реакция: Ацил-КоА+Карнитин Ацилкарнитин + HS~КоА (карнитинацилтрансфераза I) Ацилкарнитин проникает в
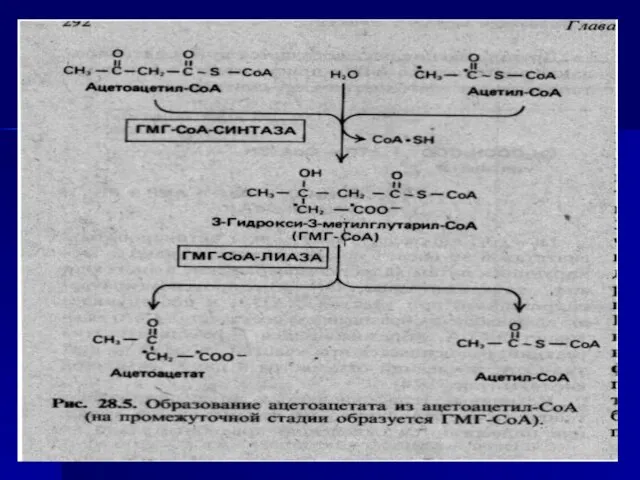
- 46. R-CH2-CH2-CH2-CO~SKoA Ацил-КоА-ДГ ФАД ФАДН2 R-CH2-CH=CH-CO~SKoA +HOH еноил-КоА-гидратаза R-CH2-CH-CH2-CO~SKoA OH гидроксил-КоА-ДГ НАД НАДН2 R-CH2-CO-CH2-CO~SKoA +HSKoA Тиолаза R-CH2-CO~SKoA
- 47. Таким образом, за один цикл (4 реакции) образуется: 1 молекула ацетил-КоА, 1 - ФАДН2 и 1
- 48. При окислении ВЖК с четным числом атомов (например, пальмитоил-КоА (16 углеродных атомов) таких циклов будет 7:
- 49. СИНТЕЗ ВЖК Главным органом, где происходит синтез большей части ВЖК, является печень. Для биосинтеза ВЖК необходимо
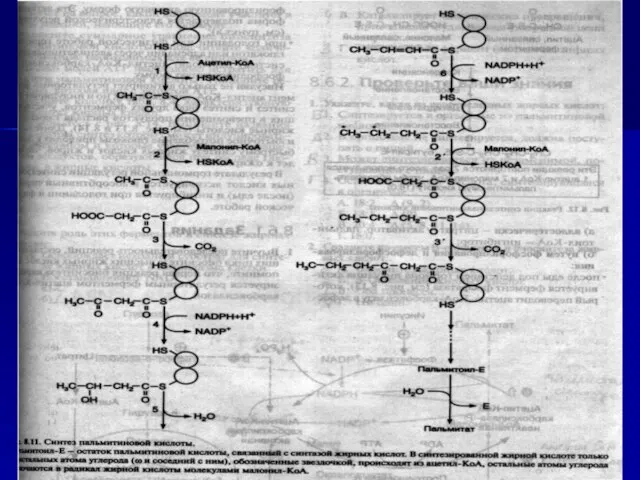
- 50. 1)СН3СО-КоА+ СО2-Ф-биотин + АТФ ацетил-КоА-карбоксилаза СООН-СН2-СО-КоА + АДФ + Рi + Ф-биот (малонил-КоА) 2) ацетил –КоА
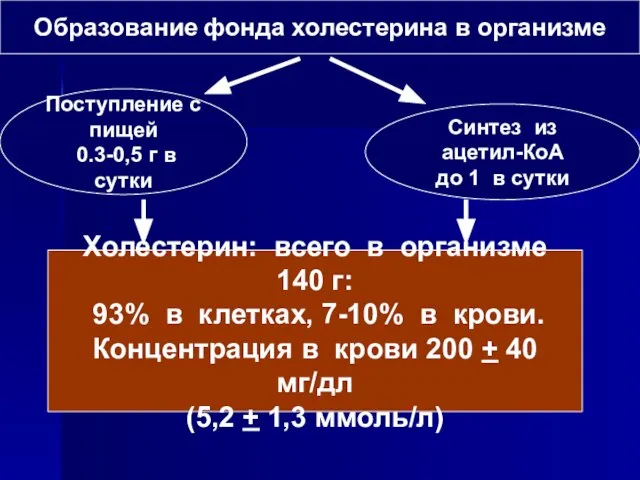
- 52. ОБМЕН ХОЛЕСТЕРИНА Холестерин содержится в организме либо в виде свободного холестерина либо в виде эфиров с
- 53. 1 Из ХОЛЕСТЕРИНА синтезируются очень важные стероиды, а именно, кортикостероидные гормоны, половые стероидные гормоны, желчные кислоты,
- 54. От микровязкости мембран, в свою очередь зависит следующее: а) функционирование мембранных белков; б) свойства рецепторных белков,
- 55. г)сопротивляемость мембран действию электрического тока. При снижении этой сопротивляемочти происходит так называемый "электрический пробой". Ток проходит
- 56. Приблизительно половина Х, образуется в самом организме путем биосинтеза, а другая половина поступает с пищей. ПУТЬ
- 57. Образование фонда холестерина в организме Поступление с пищей 0.3-0,5 г в сутки Синтез из ацетил-КоА до
- 58. I стадия: СН3СО-КоА + СН3СО-КоА ---тиолаза СН3СОСН2СО-КоА + HS-КоА 2. СН3СОСН2СОКоА + СН3СО-КоА OH СООН-СН2-С-СН2-СО-КоА СН3
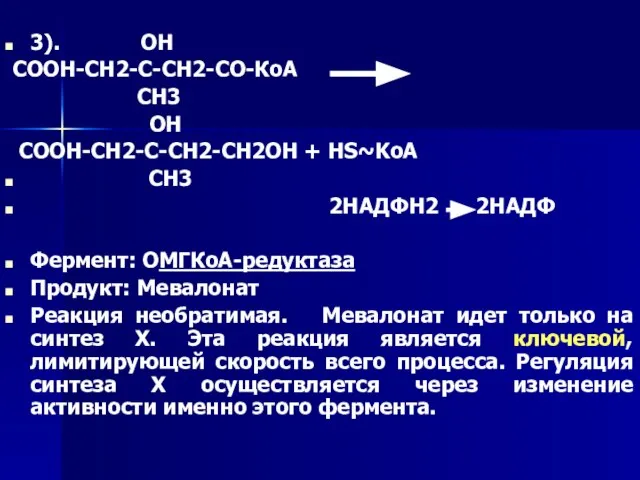
- 59. 3). ОН СООН-СН2-С-СН2-СО-КоА СН3 ОН СООН-СН2-С-СН2-СН2ОН + HS~KoA СН3 2НАДФН2 2НАДФ Фермент: ОМГКоА-редуктаза Продукт: Мевалонат Реакция
- 60. II стадия. На второй стадии мевалонат (6 С) происходит ряд реакций фосфорилирования, декарбоксилирования и конденсации. В
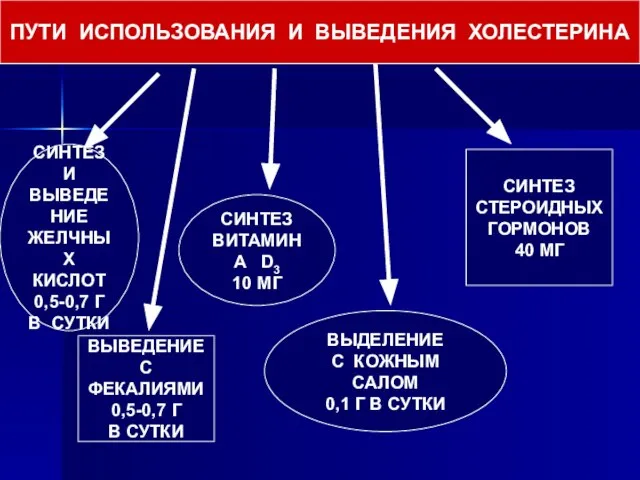
- 61. ПУТИ ИСПОЛЬЗОВАНИЯ И ВЫВЕДЕНИЯ ХОЛЕСТЕРИНА СИНТЕЗ И ВЫВЕДЕНИЕ ЖЕЛЧНЫХ КИСЛОТ 0,5-0,7 Г В СУТКИ ВЫВЕДЕНИЕ С
- 62. ФОСФОЛИПИДЫ ФЛ относятся к сложным липидам, в их молекуле присутствует фосфорная кислота. ФЛ по химическому строению
- 63. Молекула ФАГ состоит из глицерина, 2-х ВЖК, остатка фосфорной кислоты, в третьем положении глицерина (фосфатидная кислота)
- 64. Сфингомиелины представляют из себя ФЛ, в которых вместо глицерина содержится другой сложный аминоспирт - сфингозин, к
- 65. Структурная особенность ФЛ такова, что в них имеется две различных части: неполярный гидрофобный "хвост", состоящий из
- 66. Функции ФЛ. 1) В клеточных мембранах вместе с холестерином ФЛ определяют микровязкость клеточных мембран: Холестерин придает
- 67. 2) Полиненасыщенные ВЖК в составе фосфолипидов могут быть источниками гормонов или гормоноподобных веществ, таких как простагландины
- 68. 4) ФЛ входят в составе ЛП плазмы крови и вместе с апоЛП образуют гидрофильную оболочку, благодаря
- 69. 6) ФЛ входят в состав мицелл в желчи, они помогают удерживать от выпадения в осадок холестерина
- 70. Расщепление ФЛ происходит с помощью ферментов фосфолипаз. В зависимости от того, в каком положении ФЛ-аза отщепляет
- 71. 3) ФЛ-аза А2 (во втором положении), в этом случае образуется лизо-ФЛ, соединение увеличивающее проницаемость клеточных мембран.
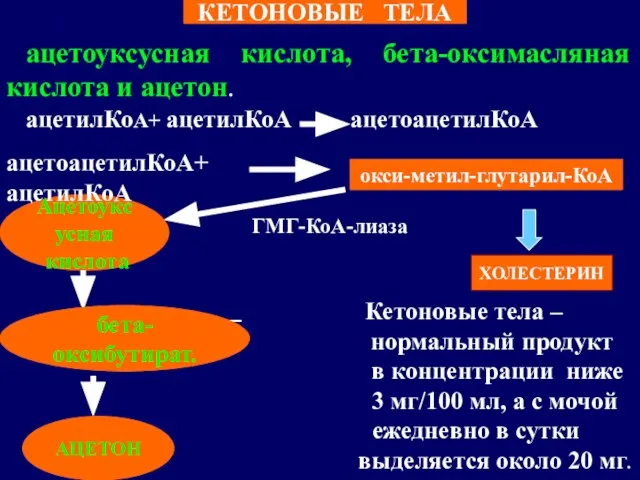
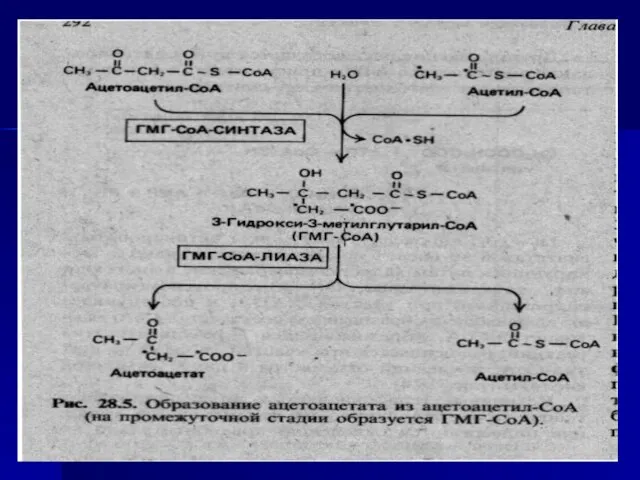
- 72. ацетоуксусная кислота, бета-оксимасляная кислота и ацетон. ацетилКоА+ ацетилКоА ацетоацетилКоА ГМГ-КоА-лиаза Кетоновые тела – нормальный продукт в
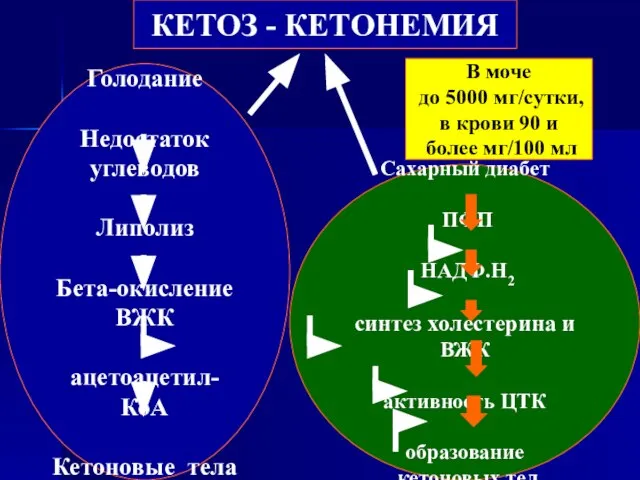
- 74. КЕТОЗ - КЕТОНЕМИЯ Голодание Недостаток углеводов Липолиз Бета-окисление ВЖК ацетоацетил-КоА Кетоновые тела Сахарный диабет ПФП НАДФ.Н2
- 75. Наступает выраженное обезвоживание. Обезвоживание и ацидоз, обусловленные кетозом, сопровождаются наличием запаха ацетона при дыхании и присутствием
- 76. Липиды выполняют многие структурные и метаболические функции, но основная их роль в обмене веществ и поддержании
- 77. В организме должны быть механизмы регуляции, которые бы направляли энергетические потоки в нужную сторону. Человек, так
- 78. В процессе липогенеза происходит превращение глюкозы и промежуточных продуктов ее метаболизма (ПВК, лактата и ацетил-КоА) в
- 79. Высокая скорость липогенеза наблюдается при приеме с пищей большого количества легко усвояемых углеводов. В жировой ткани
- 80. Из продуктов гликолиза некоторые являются прямыми предшествен-никами липидов. Это глицерофосфат, который участвует в синтезе ацилглицеролов (ТАГ
- 81. Образование ТАГ и ЛПОНП в печени увеличивается в следующих условиях: 1) При употреблении богатой углеводами пищи
- 82. Этанол - высококалорийный продукт. Но он поставляет "голодные" калории, т.к. лишен минеральных компонентов, витаминов, белков. Длительное
- 83. Этанол повышает в плазме крови содержание ТГ, усиливает периферическую мобилизацию жиров. Таким образом увеличивается доступность субстрата
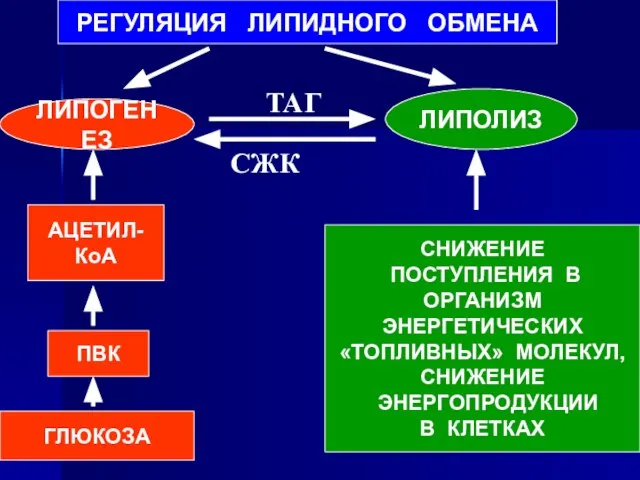
- 84. РЕГУЛЯЦИЯ ЛИПИДНОГО ОБМЕНА ЛИПОГЕНЕЗ ЛИПОЛИЗ ГЛЮКОЗА ПВК АЦЕТИЛ-КоА СНИЖЕНИЕ ПОСТУПЛЕНИЯ В ОРГАНИЗМ ЭНЕРГЕТИЧЕСКИХ «ТОПЛИВНЫХ» МОЛЕКУЛ, СНИЖЕНИЕ
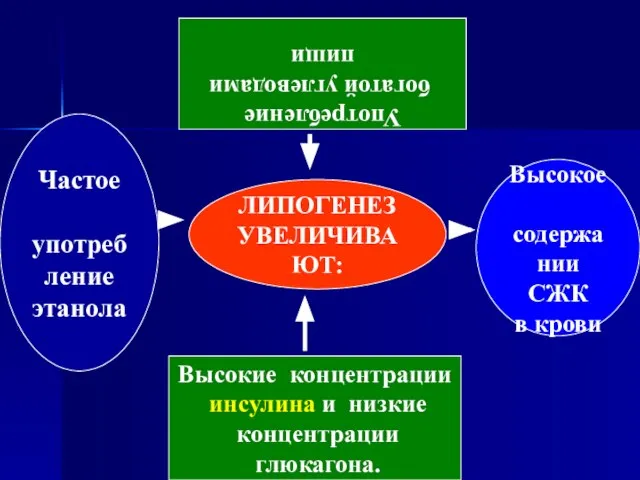
- 85. ЛИПОГЕНЕЗ УВЕЛИЧИВАЮТ: Высокое содержании СЖК в крови Частое употребление этанола Употребление богатой углеводами пищи Высокие концентрации
- 86. ПОСЛЕДОВАТЕЛЬНОСТЬ ЛИПОЛИЗА Основное депо НЖ - это жировые клетки. Сначала ТАГ расщепляется внутри жировой ткани на
- 87. Под влиянием гормонов адреналина, тироксина АКТИВИРУЕТСЯ аденилатциклаза, повышается количество цАМФ в клетке. цАМФ действует на протеинкиназу,
- 88. 2) На втором этапе свободные ЖК выходят в кровь, связываются с сывороточным альбумином и переносятся в
- 89. 3) Часть ЖК может окисляться в процессе бета-окисления. Однако, если имеется избыток глюкозы, которая покрывает энергетические
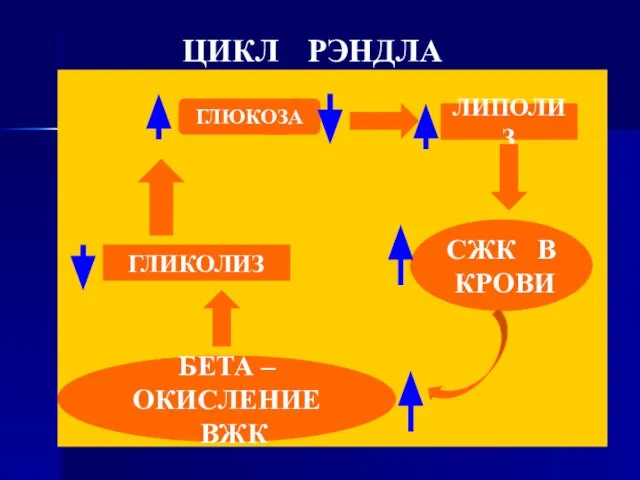
- 90. ЦИКЛ РЭНДЛА ГЛЮКОЗА ЛИПОЛИЗ СЖК В КРОВИ БЕТА – ОКИСЛЕНИЕ ВЖК ГЛИКОЛИЗ
- 91. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ЛИПОЛИЗА Скорость липогенеза и липолиза регулируется рядом гормонов, главными из которых являются ИНСУЛИН, ГЛЮКОКОРТИКОИДЫ,
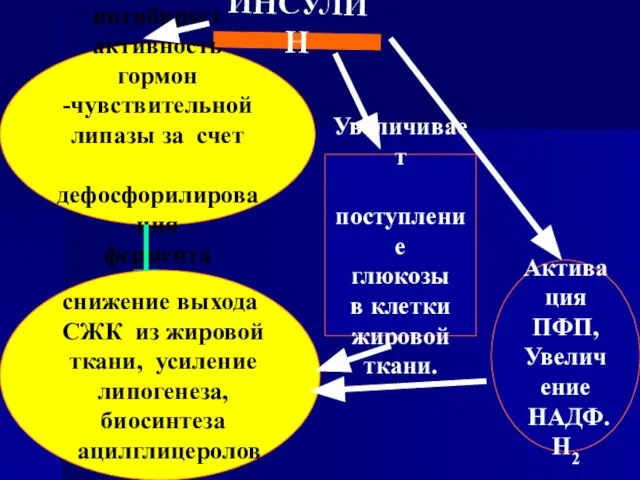
- 92. ИНСУЛИН ингибирует активность гормон -чувствительной липазы за счет дефосфорилирования фермента снижение выхода СЖК из жировой ткани,
- 93. Инсулин действует как антогонист – контргормон всех других гормонов, которые усиливают липолиз. К таким гормонам относятся
- 94. Большинство из этих гормонов действуют через цАМФ. цАМФ активирует протеинкиназу (ПК). ПК действует на неактивную липазу,
- 95. ОЖИРЕНИЕ
- 96. I Алиментарное ожирение, т.е. пищевое, возникает от переедания. А) Переедание – абсолютное, если в пище слишком
- 97. Если цикл Кребса блокируется, то Ацетил-КоА не сгорает до СО2, а идет на синтез ВЖК. Окисление
- 99. II. Гормональное ожирение. Причин гормонального ожирения может быть много. Например: А) снижение выработки гормонов глюкагона, Б)
- 100. «БОРЬБА» СО СТРЕССОМ
- 101. ЖИРОВОЕ ПЕРЕРОЖДЕНИЕ ПЕЧЕНИ Причин несколько: 1) В норме СЖК быстро удаляются из крови: часть идет на
- 102. В результате в клетках печени наблюдается жировая дистрофия или жировое перерождение. Такое ожирение печени наблюдается при
- 103. 3-я причина --- нарушение синтеза ФЛ. ТАГ и ФЛ синтезируются по сходному пути: Сначала к глицерину
- 104. При синтезе ФЛ должна образоваться эфирная связь фосфатидной кислоты с холином для образования фосфолипида - фосфатидилхолина,
- 105. 4) Четвертая причина жирового перерождения печени - алкоголизм. При длительном употреблении алкоголя в печени накапливаются СЖК,
- 106. В результате происходит усиление эстерификации ЖК с образованием ТАГ, который и накапливается в печени. Жировое перерождение
- 107. 5-я причина - действие некоторых гепатотропных ядов: СCl4 - в клинической практике часто встречается отравление этим
- 108. При некоторых хронических инфекционных заболеваниях также нарушается обмен липидов в печени, в результате чего происходит ее
- 109. : . ПОХУДАНИЕ Резкая потеря жира из подкожного и сальникового депо Длительное снижение калорийности пищи или
- 110. ГОРМОНАЛЬНЫЕ МЕХАНИЗМЫ ПОХУДАНИЯ Избыток адреналина, глюкагона резко усиливает липолиз, происходит потеря массы жировой ткани. Избыток гормонов
- 111. ацетоуксусная кислота, бета-оксимасляная кислота и ацетон. ацетилКоА+ ацетилКоА ацетоацетилКоА ГМГ-КоА-лиаза Кетоновые тела – нормальный продукт в
- 114. КЕТОЗ - КЕТОНЕМИЯ Голодание Недостаток углеводов Липолиз Бета-окисление ВЖК ацетоацетил-КоА Кетоновые тела Сахарный диабет ПФП НАДФ.Н2
- 115. ОСЛОЖНЕННИЯ КЕТОЗА Ацетоацетат и бета-оксибутират относятся к умеренно сильным кислотам, существуют в основном в виде анионов.
- 116. Одновременно, с мочой теряется большое количество жидкости. У таких больных есть тошнота и рвота, так что
- 118. Скачать презентацию
Слайд 2КЛАССИФИКАЦИЯ ЛИПИДОВ
В зависимости от состава, строения и роли в организме сложилась следующая
КЛАССИФИКАЦИЯ ЛИПИДОВ
В зависимости от состава, строения и роли в организме сложилась следующая

Слайд 32. Сложные липиды. К ним относятся фосфолипиды и гликолипиды. Они состоят из
2. Сложные липиды. К ним относятся фосфолипиды и гликолипиды. Они состоят из

Слайд 43. Отдельно следует выделить стериды - сложные эфиры ВЖК и полициклических спиртов
3. Отдельно следует выделить стериды - сложные эфиры ВЖК и полициклических спиртов

Слайд 5РОЛЬ ЛИПИДОВ В ОРГАНИЗМЕ
1) Являются компонентами мембран. В мембране обладают не только
РОЛЬ ЛИПИДОВ В ОРГАНИЗМЕ
1) Являются компонентами мембран. В мембране обладают не только

Слайд 6.
4) Могут быть предшественниками других важных соединений (простагландинов, простациклинов, тромбоксана),
.
4) Могут быть предшественниками других важных соединений (простагландинов, простациклинов, тромбоксана),

Слайд 77) Липиды влияют на иммунные реакции организма, усиливают реакции иммунитета.
8)
7) Липиды влияют на иммунные реакции организма, усиливают реакции иммунитета.
8)

Слайд 8ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЛИПИДОВ
В организм липиды поступают с жирной растительной, и особенно,
ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЛИПИДОВ
В организм липиды поступают с жирной растительной, и особенно,

Слайд 9НЖ при гидролизе распадается на глицерин и ВЖК под влиянием липазы поджелудочной
НЖ при гидролизе распадается на глицерин и ВЖК под влиянием липазы поджелудочной

Слайд 10В желчи человека преобладают холевая, и хенодезоксихолевая кислоты. Причем они находятся в
В желчи человека преобладают холевая, и хенодезоксихолевая кислоты. Причем они находятся в

Слайд 11Соли Жк, будучи поверхностно активными веществами
1) эмульгируют частицы жира и поэтому
Соли Жк, будучи поверхностно активными веществами
1) эмульгируют частицы жира и поэтому

Слайд 12СН2-0-COR CH2OH
CH-O-COR + 3H2O СHOH + 3RCOOH
CH2-O-COR CH2OH
Происходит гидролиз эфирных связей с
СН2-0-COR CH2OH
CH-O-COR + 3H2O СHOH + 3RCOOH
CH2-O-COR CH2OH
Происходит гидролиз эфирных связей с

Слайд 13Жирные кислоты не растворимы в воде, поэтому для их всасывания необходимо образование
Жирные кислоты не растворимы в воде, поэтому для их всасывания необходимо образование

Слайд 14ВЖК и глицерин теперь уже внутри клетки снова образуют молекулу триацилглицерина (ТАГ).
ВЖК и глицерин теперь уже внутри клетки снова образуют молекулу триацилглицерина (ТАГ).
Слайд 15Жиры в водной среде крови не могут находиться в свободном состоянии, т.к.
Жиры в водной среде крови не могут находиться в свободном состоянии, т.к.

Слайд 16КЛАССИФИКАЦИЯ ЛИПОПРОТЕИДОВ КРОВИ
Липопротеиды называются по плотности или по положению, которое занимают на
КЛАССИФИКАЦИЯ ЛИПОПРОТЕИДОВ КРОВИ
Липопротеиды называются по плотности или по положению, которое занимают на

Слайд 17В состав липопротеидов входят белки (апопротеины), фосфолипиды, нейтральные жиры, эфиры холестерина и
В состав липопротеидов входят белки (апопротеины), фосфолипиды, нейтральные жиры, эфиры холестерина и

Слайд 19ХИЛОМИКРОНЫ
В энтероцитах образуются хиломикроны, которые на 80% состоят из НЖ. ХМ попадают
ХИЛОМИКРОНЫ
В энтероцитах образуются хиломикроны, которые на 80% состоят из НЖ. ХМ попадают

Слайд 20 На поверхности эндотелиальных клеток, выстилающих капилляры, имеется фермент липопротеид-липаза. ЛП-липаза неактивная.
На поверхности эндотелиальных клеток, выстилающих капилляры, имеется фермент липопротеид-липаза. ЛП-липаза неактивная.

Слайд 21После отделения триацилглицеридов ХМ превращаются в хилоремнанты ("ремнанты" - остатки), которые поступают
После отделения триацилглицеридов ХМ превращаются в хилоремнанты ("ремнанты" - остатки), которые поступают

Слайд 22ЛИПОПРОТЕИДЫ ОЧЕНЬ НИЗКОЙ ПЛОТНОСТИ
ЛПОНП образуются в печени. Белка 10%, липидов - 90%
ЛИПОПРОТЕИДЫ ОЧЕНЬ НИЗКОЙ ПЛОТНОСТИ
ЛПОНП образуются в печени. Белка 10%, липидов - 90%

Слайд 23ЛПОНП после их образования в печеночных клетках выносятся в кровеносное русло и
ЛПОНП после их образования в печеночных клетках выносятся в кровеносное русло и

Слайд 24ЛИПОПРОТЕИДЫ НИЗКОЙ ПЛОТНОСТИ
ЛПНП - образуются из ЛПОНП частично в печени, частично непосредственно
ЛИПОПРОТЕИДЫ НИЗКОЙ ПЛОТНОСТИ
ЛПНП - образуются из ЛПОНП частично в печени, частично непосредственно

Слайд 25Около 50% ЛПНП распадается в печени, другая половина - во внепеченочных тканях.
Около 50% ЛПНП распадается в печени, другая половина - во внепеченочных тканях.

Слайд 26ЛИПОПРОТЕИДЫ ВЫСОКОЙ ПЛОТНОСТИ
ЛПВП – синтезируются в печени, но окончательно формируются в крови.
ЛИПОПРОТЕИДЫ ВЫСОКОЙ ПЛОТНОСТИ
ЛПВП – синтезируются в печени, но окончательно формируются в крови.

Слайд 27На поверхности ЛПВП расположены ФЛ - лецитины. Фермент ЛХАТ (лецитин-холестерин-ацил-трансфераза) от лецитина
На поверхности ЛПВП расположены ФЛ - лецитины. Фермент ЛХАТ (лецитин-холестерин-ацил-трансфераза) от лецитина

Слайд 28П Е Р Е Р Ы В

Слайд 29Концентрация ЛП в крови меняется в зависимости от:
1) приема пищи, 2)
Концентрация ЛП в крови меняется в зависимости от:
1) приема пищи, 2)

Слайд 30Нарушения липопротеидного обмена бывают наследственные, но нередки и при некоторых других заболеваниях,
Нарушения липопротеидного обмена бывают наследственные, но нередки и при некоторых других заболеваниях,

Слайд 31 Типы гиперлипопротеидемий:
I тип: повышена концентрация ХМ.
Наблюдается при наследственной недостаточности ЛП-липазы.
Типы гиперлипопротеидемий:
I тип: повышена концентрация ХМ.
Наблюдается при наследственной недостаточности ЛП-липазы.

Слайд 32II тип: повышено содержание ЛПНП или суммарной фракции ЛПОНП + ЛПНП.
Причина
II тип: повышено содержание ЛПНП или суммарной фракции ЛПОНП + ЛПНП.
Причина

Слайд 33II тип: повышено содержание ЛПНП или суммарной фракции ЛПОНП + ЛПНП.
Причина
II тип: повышено содержание ЛПНП или суммарной фракции ЛПОНП + ЛПНП.
Причина

Слайд 34II тип: повышено содержание ЛПНП или суммарной фракции ЛПОНП + ЛПНП.
Причина
II тип: повышено содержание ЛПНП или суммарной фракции ЛПОНП + ЛПНП.
Причина

Слайд 35III тип: нарушается превращение ЛПОНП в ЛПНП. В крови аномальные пре-бета-ЛП и
III тип: нарушается превращение ЛПОНП в ЛПНП. В крови аномальные пре-бета-ЛП и

Слайд 36IV тип. В крови повышено содержание ЛПОНП, Х, ТАГ. Это обычно бывает
IV тип. В крови повышено содержание ЛПОНП, Х, ТАГ. Это обычно бывает

Слайд 37V тип: повышено количество ХМ и ЛПОНП. Концентрация ЛПНП и ЛПВП обычно
V тип: повышено количество ХМ и ЛПОНП. Концентрация ЛПНП и ЛПВП обычно

Слайд 38ОБМЕН НЕЙТРАЛЬНОГО ЖИРА
НЖ состоит из смеси ТАГ (95%), ДАГ (3%) и МАГ
ОБМЕН НЕЙТРАЛЬНОГО ЖИРА
НЖ состоит из смеси ТАГ (95%), ДАГ (3%) и МАГ

Слайд 39СИНТЕЗ НЕЙТРАЛЬНОГО ЖИРА
RCOOH + HS~KoA + ATФ R-CO-SKoA + AMФ + Н4Р2О7
2)Глицерин
СИНТЕЗ НЕЙТРАЛЬНОГО ЖИРА
RCOOH + HS~KoA + ATФ R-CO-SKoA + AMФ + Н4Р2О7
2)Глицерин

Слайд 40ИСТОЧНИКИ ВЖК ДЛЯ СИНТЕЗА ТАГ (НЖ)
ПРИ РАЗНЫХ СОСТОЯНИЯХ ОРГАНИЗМА
ВЖК,
синтезируются
ИСТОЧНИКИ ВЖК ДЛЯ СИНТЕЗА ТАГ (НЖ)
ПРИ РАЗНЫХ СОСТОЯНИЯХ ОРГАНИЗМА
ВЖК,
синтезируются
Слайд 41В норме ТАГ не накапливаются в печени. Сразу же после образования они
В норме ТАГ не накапливаются в печени. Сразу же после образования они

Слайд 42
ВЖК в плазме крови связываются с альбуминами крови или же входят в
ВЖК в плазме крови связываются с альбуминами крови или же входят в
Слайд 43 БЕТА-ОКИСЛЕНИЕ ВЖК
Бета-окисление ВЖК осуществляется для получения энергии.
ВЖК освобождаются при липолизе нейтрального
БЕТА-ОКИСЛЕНИЕ ВЖК
Бета-окисление ВЖК осуществляется для получения энергии.
ВЖК освобождаются при липолизе нейтрального

Слайд 44Фермент ацил-КоА-синтетаза (тиокиназа) в присутствии АТФ и КоА превращает СЖК в активную
Фермент ацил-КоА-синтетаза (тиокиназа) в присутствии АТФ и КоА превращает СЖК в активную

Слайд 45На наружной стороне Мх-мембраны происходит следующая реакция:
Ацил-КоА+Карнитин Ацилкарнитин + HS~КоА
(карнитинацилтрансфераза I)
Ацилкарнитин
На наружной стороне Мх-мембраны происходит следующая реакция:
Ацил-КоА+Карнитин Ацилкарнитин + HS~КоА
(карнитинацилтрансфераза I)
Ацилкарнитин

Слайд 46R-CH2-CH2-CH2-CO~SKoA
Ацил-КоА-ДГ
ФАД ФАДН2
R-CH2-CH=CH-CO~SKoA
+HOH
еноил-КоА-гидратаза
R-CH2-CH-CH2-CO~SKoA
OH гидроксил-КоА-ДГ
НАД НАДН2
R-CH2-CO-CH2-CO~SKoA
Ацил-КоА-ДГ
ФАД ФАДН2
R-CH2-CH=CH-CO~SKoA
+HOH
еноил-КоА-гидратаза
R-CH2-CH-CH2-CO~SKoA
OH гидроксил-КоА-ДГ
НАД НАДН2
R-CH2-CO-CH2-CO~SKoA
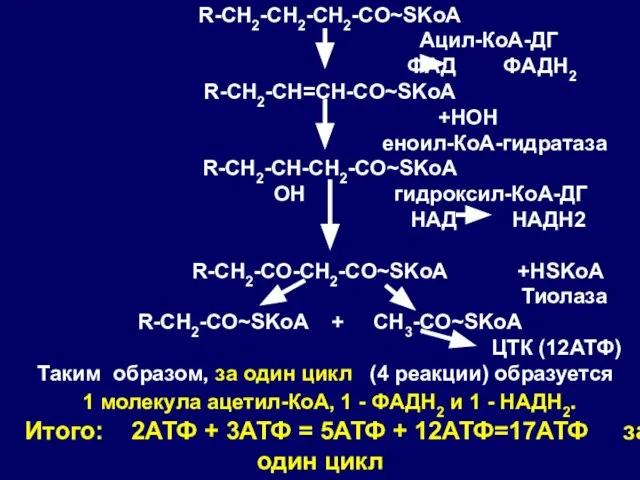
Слайд 47Таким образом, за один цикл (4 реакции) образуется:
1 молекула ацетил-КоА, 1 -
Таким образом, за один цикл (4 реакции) образуется:
1 молекула ацетил-КоА, 1 -

Слайд 48При окислении ВЖК с четным числом атомов (например, пальмитоил-КоА (16 углеродных атомов)
При окислении ВЖК с четным числом атомов (например, пальмитоил-КоА (16 углеродных атомов)

Слайд 49СИНТЕЗ ВЖК
Главным органом, где происходит синтез большей части ВЖК, является печень.
Для биосинтеза
СИНТЕЗ ВЖК
Главным органом, где происходит синтез большей части ВЖК, является печень.
Для биосинтеза

Слайд 501)СН3СО-КоА+ СО2-Ф-биотин + АТФ
ацетил-КоА-карбоксилаза
СООН-СН2-СО-КоА + АДФ + Рi + Ф-биот
(малонил-КоА)
2)
1)СН3СО-КоА+ СО2-Ф-биотин + АТФ
ацетил-КоА-карбоксилаза
СООН-СН2-СО-КоА + АДФ + Рi + Ф-биот
(малонил-КоА)
2)


Слайд 52ОБМЕН ХОЛЕСТЕРИНА
Холестерин содержится в организме либо в виде свободного холестерина либо в
ОБМЕН ХОЛЕСТЕРИНА
Холестерин содержится в организме либо в виде свободного холестерина либо в

Слайд 531 Из ХОЛЕСТЕРИНА синтезируются очень важные стероиды, а именно, кортикостероидные гормоны, половые
1 Из ХОЛЕСТЕРИНА синтезируются очень важные стероиды, а именно, кортикостероидные гормоны, половые

Слайд 54От микровязкости мембран, в свою очередь зависит следующее:
а) функционирование мембранных белков;
От микровязкости мембран, в свою очередь зависит следующее:
а) функционирование мембранных белков;

Слайд 55г)сопротивляемость мембран действию электрического тока. При снижении этой сопротивляемочти происходит так называемый
г)сопротивляемость мембран действию электрического тока. При снижении этой сопротивляемочти происходит так называемый

Слайд 56Приблизительно половина Х, образуется в самом организме путем биосинтеза, а другая половина
Приблизительно половина Х, образуется в самом организме путем биосинтеза, а другая половина

Слайд 57Образование фонда холестерина в организме
Поступление с пищей
0.3-0,5 г в сутки
Синтез из
Образование фонда холестерина в организме
Поступление с пищей
0.3-0,5 г в сутки
Синтез из
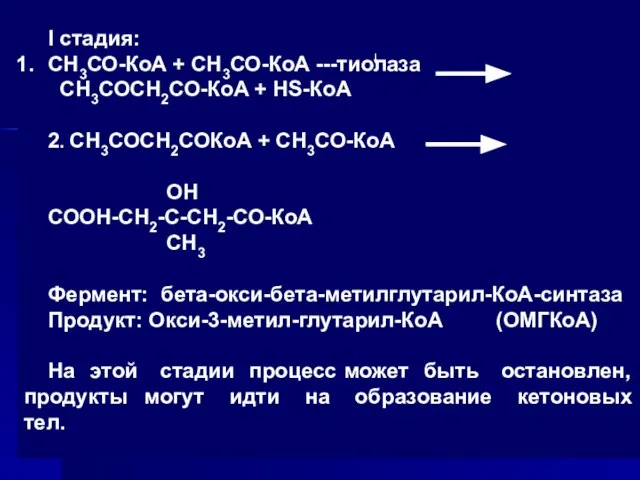
Слайд 58I стадия:
СН3СО-КоА + СН3СО-КоА ---тиолаза
СН3СОСН2СО-КоА + HS-КоА
2. СН3СОСН2СОКоА + СН3СО-КоА
I стадия:
СН3СО-КоА + СН3СО-КоА ---тиолаза
СН3СОСН2СО-КоА + HS-КоА
2. СН3СОСН2СОКоА + СН3СО-КоА
Слайд 593). ОН
СООН-СН2-С-СН2-СО-КоА
СН3
ОН
СООН-СН2-С-СН2-СН2ОН + HS~KoA
СН3
3). ОН
СООН-СН2-С-СН2-СО-КоА
СН3
ОН
СООН-СН2-С-СН2-СН2ОН + HS~KoA
СН3
Слайд 60II стадия.
На второй стадии мевалонат (6 С) происходит ряд реакций фосфорилирования, декарбоксилирования
II стадия.
На второй стадии мевалонат (6 С) происходит ряд реакций фосфорилирования, декарбоксилирования

Слайд 61ПУТИ ИСПОЛЬЗОВАНИЯ И ВЫВЕДЕНИЯ ХОЛЕСТЕРИНА
СИНТЕЗ И
ВЫВЕДЕНИЕ
ЖЕЛЧНЫХ
КИСЛОТ
0,5-0,7 Г
В СУТКИ
ВЫВЕДЕНИЕ
С ФЕКАЛИЯМИ
0,5-0,7 Г
В СУТКИ
СИНТЕЗ
ВИТАМИНА
ПУТИ ИСПОЛЬЗОВАНИЯ И ВЫВЕДЕНИЯ ХОЛЕСТЕРИНА
СИНТЕЗ И
ВЫВЕДЕНИЕ
ЖЕЛЧНЫХ
КИСЛОТ
0,5-0,7 Г
В СУТКИ
ВЫВЕДЕНИЕ
С ФЕКАЛИЯМИ
0,5-0,7 Г
В СУТКИ
СИНТЕЗ
ВИТАМИНА
Слайд 62ФОСФОЛИПИДЫ
ФЛ относятся к сложным липидам, в их молекуле присутствует фосфорная кислота.
ФЛ по
ФОСФОЛИПИДЫ
ФЛ относятся к сложным липидам, в их молекуле присутствует фосфорная кислота.
ФЛ по

Слайд 63Молекула ФАГ состоит из глицерина, 2-х ВЖК, остатка фосфорной кислоты, в третьем
Молекула ФАГ состоит из глицерина, 2-х ВЖК, остатка фосфорной кислоты, в третьем

Слайд 64Сфингомиелины представляют из себя ФЛ, в которых вместо глицерина содержится другой сложный
Сфингомиелины представляют из себя ФЛ, в которых вместо глицерина содержится другой сложный

Слайд 65Структурная особенность ФЛ такова, что в них имеется две различных части: неполярный
Структурная особенность ФЛ такова, что в них имеется две различных части: неполярный

Слайд 66Функции ФЛ.
1) В клеточных мембранах вместе с холестерином ФЛ определяют микровязкость клеточных
Функции ФЛ.
1) В клеточных мембранах вместе с холестерином ФЛ определяют микровязкость клеточных

Слайд 672) Полиненасыщенные ВЖК в составе фосфолипидов могут быть источниками гормонов или гормоноподобных
2) Полиненасыщенные ВЖК в составе фосфолипидов могут быть источниками гормонов или гормоноподобных

Слайд 684) ФЛ входят в составе ЛП плазмы крови и вместе с апоЛП
4) ФЛ входят в составе ЛП плазмы крови и вместе с апоЛП

Слайд 696) ФЛ входят в состав мицелл в желчи, они помогают удерживать от
6) ФЛ входят в состав мицелл в желчи, они помогают удерживать от

Слайд 70Расщепление ФЛ происходит с помощью ферментов фосфолипаз. В зависимости от того, в
Расщепление ФЛ происходит с помощью ферментов фосфолипаз. В зависимости от того, в

Слайд 713) ФЛ-аза А2 (во втором положении), в этом случае образуется лизо-ФЛ, соединение
3) ФЛ-аза А2 (во втором положении), в этом случае образуется лизо-ФЛ, соединение

Слайд 72ацетоуксусная кислота, бета-оксимасляная кислота и ацетон.
ацетилКоА+ ацетилКоА ацетоацетилКоА
ГМГ-КоА-лиаза
Кетоновые тела –
ацетоуксусная кислота, бета-оксимасляная кислота и ацетон.
ацетилКоА+ ацетилКоА ацетоацетилКоА
ГМГ-КоА-лиаза
Кетоновые тела –
Слайд 74КЕТОЗ - КЕТОНЕМИЯ
Голодание
Недостаток углеводов
Липолиз
Бета-окисление ВЖК
ацетоацетил-КоА
Кетоновые тела
Сахарный диабет
ПФП
НАДФ.Н2
синтез холестерина и ВЖК
активность ЦТК
образование
КЕТОЗ - КЕТОНЕМИЯ
Голодание
Недостаток углеводов
Липолиз
Бета-окисление ВЖК
ацетоацетил-КоА
Кетоновые тела
Сахарный диабет
ПФП
НАДФ.Н2
синтез холестерина и ВЖК
активность ЦТК
образование
Слайд 75Наступает выраженное обезвоживание. Обезвоживание и ацидоз, обусловленные кетозом, сопровождаются наличием запаха ацетона
Наступает выраженное обезвоживание. Обезвоживание и ацидоз, обусловленные кетозом, сопровождаются наличием запаха ацетона

Слайд 76Липиды выполняют многие структурные и метаболические функции, но основная их роль в
Липиды выполняют многие структурные и метаболические функции, но основная их роль в

Слайд 77В организме должны быть механизмы регуляции, которые бы направляли энергетические потоки в
В организме должны быть механизмы регуляции, которые бы направляли энергетические потоки в

Слайд 78В процессе липогенеза происходит превращение глюкозы и промежуточных продуктов ее метаболизма (ПВК,
В процессе липогенеза происходит превращение глюкозы и промежуточных продуктов ее метаболизма (ПВК,

Слайд 79Высокая скорость липогенеза наблюдается при приеме с пищей большого количества легко усвояемых
Высокая скорость липогенеза наблюдается при приеме с пищей большого количества легко усвояемых

Слайд 80Из продуктов гликолиза некоторые являются прямыми предшествен-никами липидов. Это глицерофосфат, который участвует
Из продуктов гликолиза некоторые являются прямыми предшествен-никами липидов. Это глицерофосфат, который участвует

Слайд 81Образование ТАГ и ЛПОНП в печени увеличивается в следующих условиях:
1) При употреблении
Образование ТАГ и ЛПОНП в печени увеличивается в следующих условиях:
1) При употреблении

Слайд 82Этанол - высококалорийный продукт. Но он поставляет "голодные" калории, т.к. лишен минеральных
Этанол - высококалорийный продукт. Но он поставляет "голодные" калории, т.к. лишен минеральных

Слайд 83Этанол повышает в плазме крови содержание ТГ, усиливает периферическую мобилизацию жиров. Таким
Этанол повышает в плазме крови содержание ТГ, усиливает периферическую мобилизацию жиров. Таким

Слайд 84РЕГУЛЯЦИЯ ЛИПИДНОГО ОБМЕНА
ЛИПОГЕНЕЗ
ЛИПОЛИЗ
ГЛЮКОЗА
ПВК
АЦЕТИЛ-КоА
СНИЖЕНИЕ
ПОСТУПЛЕНИЯ В
ОРГАНИЗМ
ЭНЕРГЕТИЧЕСКИХ
«ТОПЛИВНЫХ» МОЛЕКУЛ,
СНИЖЕНИЕ
ЭНЕРГОПРОДУКЦИИ
В КЛЕТКАХ
СЖК
ТАГ
РЕГУЛЯЦИЯ ЛИПИДНОГО ОБМЕНА
ЛИПОГЕНЕЗ
ЛИПОЛИЗ
ГЛЮКОЗА
ПВК
АЦЕТИЛ-КоА
СНИЖЕНИЕ
ПОСТУПЛЕНИЯ В
ОРГАНИЗМ
ЭНЕРГЕТИЧЕСКИХ
«ТОПЛИВНЫХ» МОЛЕКУЛ,
СНИЖЕНИЕ
ЭНЕРГОПРОДУКЦИИ
В КЛЕТКАХ
СЖК
ТАГ
Слайд 85ЛИПОГЕНЕЗ
УВЕЛИЧИВАЮТ:
Высокое
содержании
СЖК
в крови
Частое
употребление
этанола
Употребление
богатой углеводами
пищи
Высокие
ЛИПОГЕНЕЗ
УВЕЛИЧИВАЮТ:
Высокое
содержании
СЖК
в крови
Частое
употребление
этанола
Употребление
богатой углеводами
пищи
Высокие
Слайд 86ПОСЛЕДОВАТЕЛЬНОСТЬ ЛИПОЛИЗА
Основное депо НЖ - это жировые клетки.
Сначала ТАГ расщепляется внутри жировой
ПОСЛЕДОВАТЕЛЬНОСТЬ ЛИПОЛИЗА
Основное депо НЖ - это жировые клетки.
Сначала ТАГ расщепляется внутри жировой

Слайд 87Под влиянием гормонов адреналина, тироксина АКТИВИРУЕТСЯ аденилатциклаза, повышается количество цАМФ в клетке.
Под влиянием гормонов адреналина, тироксина АКТИВИРУЕТСЯ аденилатциклаза, повышается количество цАМФ в клетке.

Слайд 882) На втором этапе свободные ЖК выходят в кровь, связываются с сывороточным
2) На втором этапе свободные ЖК выходят в кровь, связываются с сывороточным

Слайд 893) Часть ЖК может окисляться в процессе бета-окисления.
Однако, если имеется избыток глюкозы,
3) Часть ЖК может окисляться в процессе бета-окисления.
Однако, если имеется избыток глюкозы,

Слайд 90ЦИКЛ РЭНДЛА
ГЛЮКОЗА
ЛИПОЛИЗ
СЖК В
КРОВИ
БЕТА – ОКИСЛЕНИЕ
ВЖК
ГЛИКОЛИЗ
ЦИКЛ РЭНДЛА
ГЛЮКОЗА
ЛИПОЛИЗ
СЖК В
КРОВИ
БЕТА – ОКИСЛЕНИЕ
ВЖК
ГЛИКОЛИЗ
Слайд 91ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ЛИПОЛИЗА
Скорость липогенеза и липолиза регулируется рядом гормонов, главными из которых
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ЛИПОЛИЗА
Скорость липогенеза и липолиза регулируется рядом гормонов, главными из которых

Слайд 92ИНСУЛИН
ингибирует
активность гормон
-чувствительной
липазы за счет
дефосфорилирования
фермента
снижение выхода
СЖК из жировой
ткани,
ИНСУЛИН
ингибирует
активность гормон
-чувствительной
липазы за счет
дефосфорилирования
фермента
снижение выхода
СЖК из жировой
ткани,
Слайд 93Инсулин действует как антогонист – контргормон всех других гормонов, которые усиливают липолиз.
Инсулин действует как антогонист – контргормон всех других гормонов, которые усиливают липолиз.

Слайд 94Большинство из этих гормонов действуют через цАМФ. цАМФ активирует протеинкиназу (ПК).
ПК
Большинство из этих гормонов действуют через цАМФ. цАМФ активирует протеинкиназу (ПК).
ПК

Слайд 95ОЖИРЕНИЕ
ОЖИРЕНИЕ

Слайд 96I Алиментарное ожирение, т.е. пищевое, возникает от переедания.
А) Переедание – абсолютное,
I Алиментарное ожирение, т.е. пищевое, возникает от переедания.
А) Переедание – абсолютное,

Слайд 97Если цикл Кребса блокируется, то Ацетил-КоА не сгорает до СО2, а идет
Если цикл Кребса блокируется, то Ацетил-КоА не сгорает до СО2, а идет


Слайд 99II. Гормональное ожирение.
Причин гормонального ожирения может быть много. Например:
А) снижение выработки
II. Гормональное ожирение.
Причин гормонального ожирения может быть много. Например:
А) снижение выработки

Слайд 100«БОРЬБА» СО СТРЕССОМ
«БОРЬБА» СО СТРЕССОМ

Слайд 101ЖИРОВОЕ ПЕРЕРОЖДЕНИЕ ПЕЧЕНИ
Причин несколько:
1) В норме СЖК быстро удаляются из крови: часть
ЖИРОВОЕ ПЕРЕРОЖДЕНИЕ ПЕЧЕНИ
Причин несколько:
1) В норме СЖК быстро удаляются из крови: часть

Слайд 102В результате в клетках печени наблюдается жировая дистрофия или жировое перерождение.
Такое
В результате в клетках печени наблюдается жировая дистрофия или жировое перерождение.
Такое

Слайд 1033-я причина --- нарушение синтеза ФЛ.
ТАГ и ФЛ синтезируются по сходному
3-я причина --- нарушение синтеза ФЛ.
ТАГ и ФЛ синтезируются по сходному

Слайд 104При синтезе ФЛ должна образоваться эфирная связь фосфатидной кислоты с холином для
При синтезе ФЛ должна образоваться эфирная связь фосфатидной кислоты с холином для

Слайд 1054) Четвертая причина жирового перерождения печени - алкоголизм.
При длительном употреблении алкоголя
4) Четвертая причина жирового перерождения печени - алкоголизм.
При длительном употреблении алкоголя

Слайд 106В результате происходит усиление эстерификации ЖК с образованием ТАГ, который и накапливается
В результате происходит усиление эстерификации ЖК с образованием ТАГ, который и накапливается

Слайд 1075-я причина - действие некоторых гепатотропных ядов:
СCl4 - в клинической практике
5-я причина - действие некоторых гепатотропных ядов:
СCl4 - в клинической практике

Слайд 108При некоторых хронических инфекционных заболеваниях также нарушается обмен липидов в печени, в
При некоторых хронических инфекционных заболеваниях также нарушается обмен липидов в печени, в

Слайд 109:
.
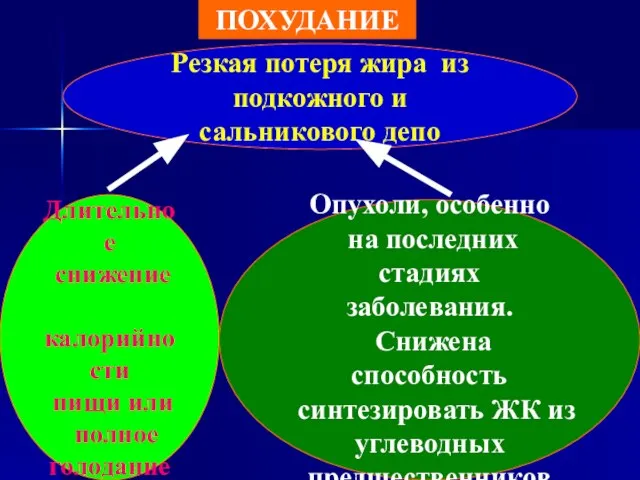
ПОХУДАНИЕ
Резкая потеря жира из
подкожного и сальникового депо
Длительное
снижение
калорийности
.
ПОХУДАНИЕ
Резкая потеря жира из
подкожного и сальникового депо
Длительное
снижение
калорийности
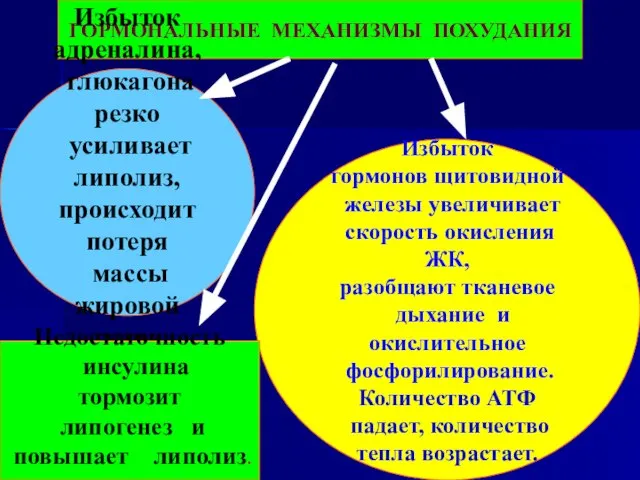
Слайд 110ГОРМОНАЛЬНЫЕ МЕХАНИЗМЫ ПОХУДАНИЯ
Избыток
адреналина,
глюкагона резко
усиливает липолиз,
происходит потеря
массы жировой
ГОРМОНАЛЬНЫЕ МЕХАНИЗМЫ ПОХУДАНИЯ
Избыток
адреналина,
глюкагона резко
усиливает липолиз,
происходит потеря
массы жировой
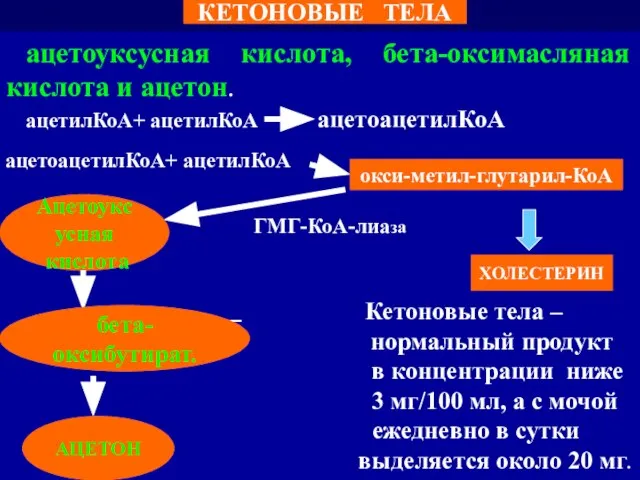
Слайд 111ацетоуксусная кислота, бета-оксимасляная кислота и ацетон.
ацетилКоА+ ацетилКоА ацетоацетилКоА
ГМГ-КоА-лиаза
Кетоновые тела –
ацетоуксусная кислота, бета-оксимасляная кислота и ацетон.
ацетилКоА+ ацетилКоА ацетоацетилКоА
ГМГ-КоА-лиаза
Кетоновые тела –
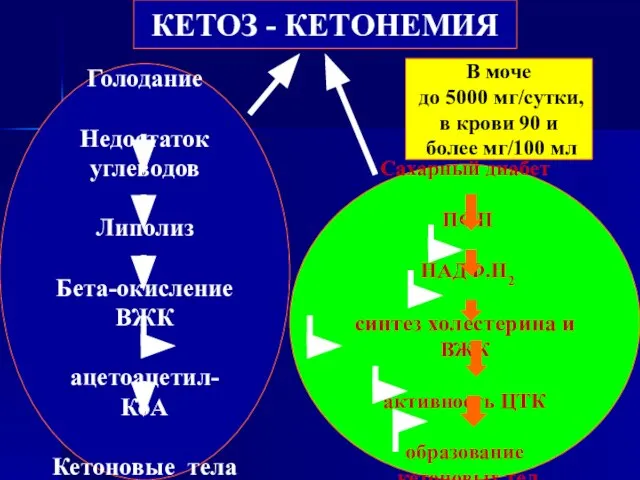
Слайд 114КЕТОЗ - КЕТОНЕМИЯ
Голодание
Недостаток углеводов
Липолиз
Бета-окисление ВЖК
ацетоацетил-КоА
Кетоновые тела
Сахарный диабет
ПФП
НАДФ.Н2
синтез холестерина и ВЖК
активность ЦТК
образование
КЕТОЗ - КЕТОНЕМИЯ
Голодание
Недостаток углеводов
Липолиз
Бета-окисление ВЖК
ацетоацетил-КоА
Кетоновые тела
Сахарный диабет
ПФП
НАДФ.Н2
синтез холестерина и ВЖК
активность ЦТК
образование
Слайд 115ОСЛОЖНЕННИЯ КЕТОЗА
Ацетоацетат и бета-оксибутират относятся к умеренно сильным кислотам, существуют в основном
ОСЛОЖНЕННИЯ КЕТОЗА
Ацетоацетат и бета-оксибутират относятся к умеренно сильным кислотам, существуют в основном

Слайд 116Одновременно, с мочой теряется большое количество жидкости. У таких больных есть тошнота
Одновременно, с мочой теряется большое количество жидкости. У таких больных есть тошнота

 КОЛЛЕКЦИЯ АРОМАТОВ ДЛЯ ГЕЛЕЙ ДЛЯ ДУША И ПЕНЫ ДЛЯ ВАНН Изысканные,тонизирующие и опьяняющие ароматы с запахом наших любимых напитк
КОЛЛЕКЦИЯ АРОМАТОВ ДЛЯ ГЕЛЕЙ ДЛЯ ДУША И ПЕНЫ ДЛЯ ВАНН Изысканные,тонизирующие и опьяняющие ароматы с запахом наших любимых напитк Медуза Горгона: несчастная жертва или жестокое чудовище?
Медуза Горгона: несчастная жертва или жестокое чудовище? Процесс принятия и реализации управленческих решений
Процесс принятия и реализации управленческих решений Сетевые структуры в современной мировой политике
Сетевые структуры в современной мировой политике Конструктивный рисунок с натуры
Конструктивный рисунок с натуры Тематическое и поурочное планирование курса «ОРКиСЭ»
Тематическое и поурочное планирование курса «ОРКиСЭ» Доклад на тему Белл-ланкастерская система обучения
Доклад на тему Белл-ланкастерская система обучения ? Всем привет! ?меня зовут Надеждав 17:00(по Москве) начнём!В11: Стереометрия
? Всем привет! ?меня зовут Надеждав 17:00(по Москве) начнём!В11: Стереометрия Факультет Международного туризма
Факультет Международного туризма 1 Тестирование производительности веб–приложений: Как перестать беспокоиться и начать делать ЭТО Тимур Хайруллин Организатор.
1 Тестирование производительности веб–приложений: Как перестать беспокоиться и начать делать ЭТО Тимур Хайруллин Организатор. ДОПОЛНИТЕЛЬНОЕ ОБРАЗОВАНИЕ В ШКОЛЕ
ДОПОЛНИТЕЛЬНОЕ ОБРАЗОВАНИЕ В ШКОЛЕ САМОУПРАВЛЕНИЕ ОБУЧАЮЩИХСЯ
САМОУПРАВЛЕНИЕ ОБУЧАЮЩИХСЯ Встраиваемая бытовая техника
Встраиваемая бытовая техника Ёкаи в Японии
Ёкаи в Японии Антропогенное воздействие на литосферу
Антропогенное воздействие на литосферу Основные сценарии использования мобильных платформ для работы с системами электронного документооборота: проблемы и поиск пут
Основные сценарии использования мобильных платформ для работы с системами электронного документооборота: проблемы и поиск пут Презентация по теме: Химические реакции
Презентация по теме: Химические реакции Смешанные колориты
Смешанные колориты Презентация к уроку английского языка в 10 классе
Презентация к уроку английского языка в 10 классе 20161215_21.12.16.prezent_k_seminaru_geografii_0
20161215_21.12.16.prezent_k_seminaru_geografii_0 Юбилею М.В.Ломоносова посвящается …
Юбилею М.В.Ломоносова посвящается … Гиперактивный ребёнок в школе
Гиперактивный ребёнок в школе Открытый урок математики в 5 классе
Открытый урок математики в 5 классе Презентация на тему Общее знакомство с растениями Мир растений
Презентация на тему Общее знакомство с растениями Мир растений Процессуальные решения и документы, сроки и судебные издержки. Гражданский иск в уголовном процессе
Процессуальные решения и документы, сроки и судебные издержки. Гражданский иск в уголовном процессе Анализ основ рекламной деятельности бренда “BMW M5”
Анализ основ рекламной деятельности бренда “BMW M5” es-Persona индивидуальные заказы
es-Persona индивидуальные заказы +
+