- Макроэволюция

Содержание
- 2. Макроэволюция: разметка участка Крайние точки, оконтуривающие круг макроэволюционных процессов: Араморфоз - идиоадаптация Дивергенция – параллелизм -
- 3. Макроэволюция - разметка участка Изучение палеонтологических находок и современных форм позволяет в настоящее время считать достаточно
- 4. Макроэволюция - разметка участка Эволюционные преобразования, которые значительно увеличивают адаптивную возможность данной группы, (позволяя расширить старую
- 5. Макроэволюция - разметка участка А. Н.Северцов назвал эволюционные преобразования организации, ведущие к морфофизиологическому прогрессу, ароморфозами. Согласно
- 6. Макроэволюция - разметка участка Верхняя и нижняя челюсти и зубы различных змей 1 – питон, 2,7
- 7. Макроэволюция - разметка участка Миксоспоридии – пример катагенеза Генетический анализ показал, что миксоспоридии, всегда считавшиеся «хорошим»
- 8. Макроэволюция - разметка участка Дивергенция (от средневекового лат. divergo — отклоняюсь) Росомаха питается преимущес-твенно падалью. Широкие
- 9. Макроэволюция - разметка участка Конвергенция не есть полная противо-положность дивергенции. Дивергируют таксоны, а конвергенция – это
- 10. Макроэволюция - разметка участка Параллелизм – конвергенция гомологичных органов Поза «стояния столбиком» и характерное для грызунов
- 11. Макроэволюция - разметка участка Термин параллелизм употребляется также не для характеристи-ки изменения органов, а для указания
- 12. Макроэволюция - разметка участка Полифилия – объединение в один таксон разных по происхождению групп. Поли- много,
- 13. Макроэволюция – современные сложности Нейтралистская теория эволюции – нейтральные признаки только дивергируют. Парафилия и полифилия на
- 14. Филогенетические связи между 8 группами позвоночных животных (верхняя схема) и различия по числу аминокислотных замен (Каа)
- 15. Макроэволюция – современные сложности К настоящему времени возможности морфологического, эмбриологического и палеонтологического методов анализа филогении практически
- 16. Макроэволюция – современные сложности Эволюционное дерево эукариот. Точки ответвления от основного ствола выше эвгленозоев проставлены произвольно.
- 17. Горизонтальный перенос генов – к чему это приводит Прокариоты представлены двумя царствами – архебактерии и эубактерии
- 18. Биологический прогресс: противоречие между сложностью и успехом А.Н. Северцов снимает его, разделяя биологический и морфо-физиологический прогресс
- 19. Основная философская проблема макроэволюции - направленность Ранние эволюционисты объясняли развитие природы через конечные причины, стремлением к
- 21. Скачать презентацию
Слайд 2Макроэволюция: разметка участка
Крайние точки, оконтуривающие круг макроэволюционных процессов:
Араморфоз - идиоадаптация
Дивергенция – параллелизм
Макроэволюция: разметка участка
Крайние точки, оконтуривающие круг макроэволюционных процессов:
Араморфоз - идиоадаптация
Дивергенция – параллелизм

Слайд 3Макроэволюция - разметка участка
Изучение палеонтологических находок и современных форм позволяет в настоящее
Макроэволюция - разметка участка
Изучение палеонтологических находок и современных форм позволяет в настоящее

Слайд 4Макроэволюция - разметка участка
Эволюционные преобразования, которые значительно увеличивают адаптивную возможность данной группы,
Макроэволюция - разметка участка
Эволюционные преобразования, которые значительно увеличивают адаптивную возможность данной группы,
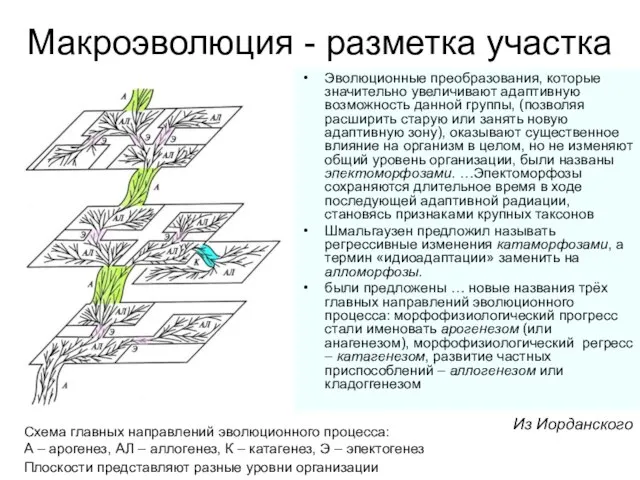
Слайд 5Макроэволюция - разметка участка
А. Н.Северцов назвал эволюционные преобразования организации, ведущие к морфофизиологическому
Макроэволюция - разметка участка
А. Н.Северцов назвал эволюционные преобразования организации, ведущие к морфофизиологическому

Слайд 6Макроэволюция - разметка участка
Верхняя и нижняя челюсти и зубы различных змей
1 –
Макроэволюция - разметка участка
Верхняя и нижняя челюсти и зубы различных змей
1 –

Слайд 7Макроэволюция - разметка участка
Миксоспоридии – пример катагенеза
Генетический анализ показал, что миксоспоридии, всегда
Макроэволюция - разметка участка
Миксоспоридии – пример катагенеза
Генетический анализ показал, что миксоспоридии, всегда
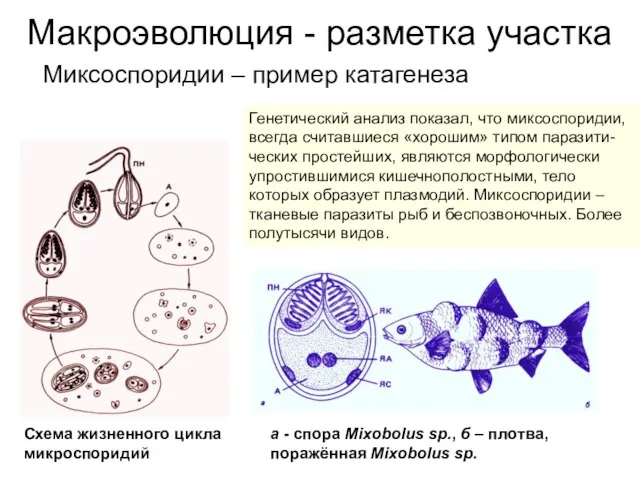
Слайд 8Макроэволюция - разметка участка
Дивергенция (от средневекового лат. divergo — отклоняюсь)
Росомаха питается преимущес-твенно
Макроэволюция - разметка участка
Дивергенция (от средневекового лат. divergo — отклоняюсь)
Росомаха питается преимущес-твенно

Слайд 9Макроэволюция - разметка участка
Конвергенция не есть полная противо-положность дивергенции. Дивергируют таксоны, а
Макроэволюция - разметка участка
Конвергенция не есть полная противо-положность дивергенции. Дивергируют таксоны, а

Слайд 10Макроэволюция - разметка участка
Параллелизм – конвергенция гомологичных органов
Поза «стояния столбиком» и характерное
Макроэволюция - разметка участка
Параллелизм – конвергенция гомологичных органов
Поза «стояния столбиком» и характерное
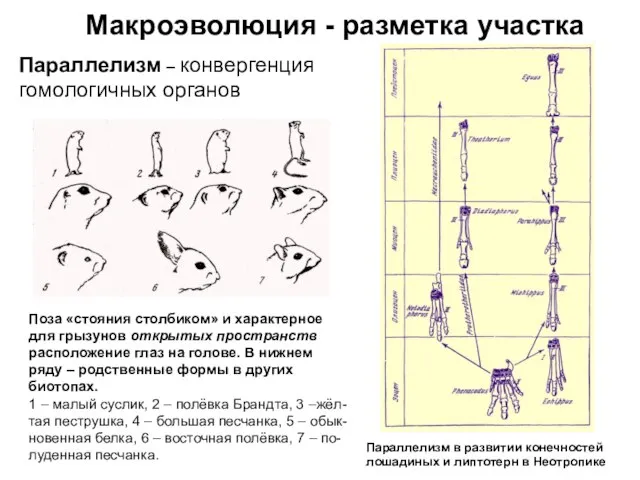
Слайд 11Макроэволюция - разметка участка
Термин параллелизм употребляется также не для характеристи-ки изменения органов,
Макроэволюция - разметка участка
Термин параллелизм употребляется также не для характеристи-ки изменения органов,

Слайд 12Макроэволюция - разметка участка
Полифилия – объединение в один таксон разных по происхождению
Макроэволюция - разметка участка
Полифилия – объединение в один таксон разных по происхождению

Слайд 13Макроэволюция – современные сложности
Нейтралистская теория эволюции – нейтральные признаки только дивергируют. Парафилия
Макроэволюция – современные сложности
Нейтралистская теория эволюции – нейтральные признаки только дивергируют. Парафилия

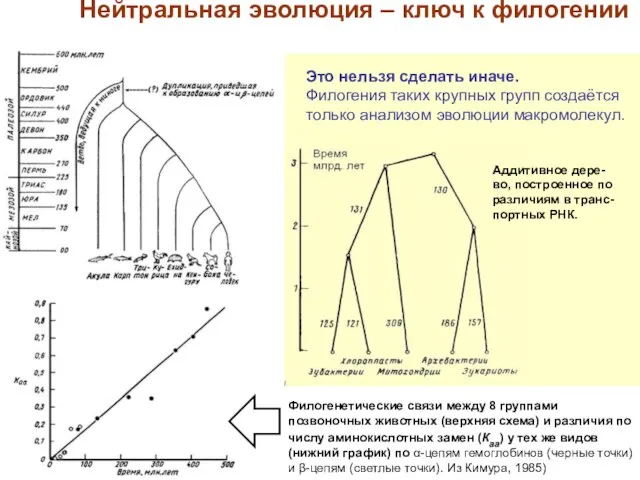
Слайд 14Филогенетические связи между 8 группами позвоночных животных (верхняя схема) и различия по
Филогенетические связи между 8 группами позвоночных животных (верхняя схема) и различия по
Слайд 15Макроэволюция – современные сложности
К настоящему времени возможности морфологического, эмбриологического и палеонтологического методов
Макроэволюция – современные сложности
К настоящему времени возможности морфологического, эмбриологического и палеонтологического методов

Слайд 16Макроэволюция – современные сложности
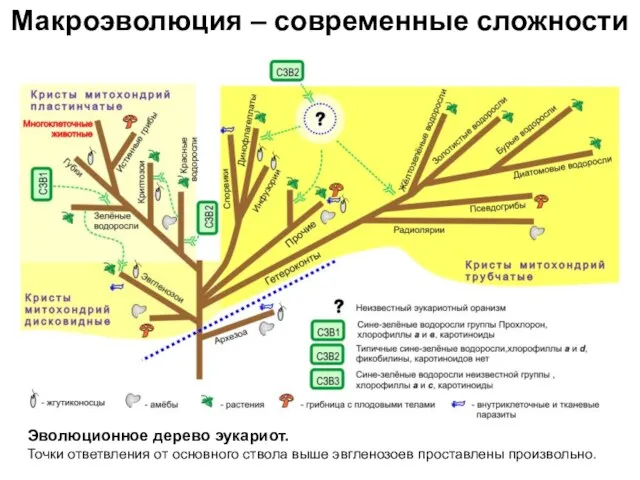
Эволюционное дерево эукариот.
Точки ответвления от основного ствола выше
Макроэволюция – современные сложности
Эволюционное дерево эукариот. Точки ответвления от основного ствола выше
Слайд 17Горизонтальный перенос генов – к чему это приводит
Прокариоты представлены двумя царствами –
Горизонтальный перенос генов – к чему это приводит
Прокариоты представлены двумя царствами –

Слайд 18Биологический прогресс: противоречие между сложностью и успехом
А.Н. Северцов снимает его, разделяя биологический
Биологический прогресс: противоречие между сложностью и успехом
А.Н. Северцов снимает его, разделяя биологический

Слайд 19Основная философская проблема макроэволюции - направленность
Ранние эволюционисты объясняли развитие природы через конечные
Основная философская проблема макроэволюции - направленность
Ранние эволюционисты объясняли развитие природы через конечные

 Правовое обеспечение деятельности государственных таможенных органов
Правовое обеспечение деятельности государственных таможенных органов Технологический процесс работы пассажирской технической станции Пятилетка
Технологический процесс работы пассажирской технической станции Пятилетка Тема: «Формирование эмоционально-ценностного отношения к миру у учащихся через экологический модуль на уроках химии»
Тема: «Формирование эмоционально-ценностного отношения к миру у учащихся через экологический модуль на уроках химии» Evolution3 система «открытого типа» с неограниченной возможностью наращивания функций. Сетевая архитектура построения комплекса спос
Evolution3 система «открытого типа» с неограниченной возможностью наращивания функций. Сетевая архитектура построения комплекса спос Астраханский Кремль
Астраханский Кремль О принципах формирования «Национальной комплексной программы профилактики суицидального поведения несовершеннолетних в Казах
О принципах формирования «Национальной комплексной программы профилактики суицидального поведения несовершеннолетних в Казах Презентация на тему Бесполое и половое размножение организмов
Презентация на тему Бесполое и половое размножение организмов  Составление сетчатых орнаментов на основе законов симметрии
Составление сетчатых орнаментов на основе законов симметрии Презентация на тему Равнины суши
Презентация на тему Равнины суши Откуда берутся снег и лед?«Окружающий мир» 1 класс
Откуда берутся снег и лед?«Окружающий мир» 1 класс Система экспресс-обработки НТМИ
Система экспресс-обработки НТМИ О правилах приема в ОУ
О правилах приема в ОУ Если б Гарри Поттером был я…
Если б Гарри Поттером был я… Отрасли права
Отрасли права Мотивация. Виды мотивации
Мотивация. Виды мотивации Что такое система?
Что такое система? Презентация на тему Мера ответственности родителей за воспитание детей
Презентация на тему Мера ответственности родителей за воспитание детей Проблемные мотивации на уроках русского языка и литературы и пути их решения
Проблемные мотивации на уроках русского языка и литературы и пути их решения Глаза - один из ценнейших органов чувств человека. Благодаря глазам мы получаем почти всю информацию об окружающем мире.
Глаза - один из ценнейших органов чувств человека. Благодаря глазам мы получаем почти всю информацию об окружающем мире. 7.1В Влияние деятельности человека на окружающую среду_презентация
7.1В Влияние деятельности человека на окружающую среду_презентация Регенерация
Регенерация Уральские горы (4 класс)
Уральские горы (4 класс) Занятие по обществознанию на тему: «Политические партии» Для студентов 1 курса СПО
Занятие по обществознанию на тему: «Политические партии» Для студентов 1 курса СПО Москва-река
Москва-река Украинский национальный костюм
Украинский национальный костюм Презентация%20по%20теме%20правописание%20приставок%20и%20предлогов
Презентация%20по%20теме%20правописание%20приставок%20и%20предлогов Олимпийские символы
Олимпийские символы Структура и содержание уголовно-исполнительного законодательства РФ
Структура и содержание уголовно-исполнительного законодательства РФ