- Методи досліджень та ідентифікаціi вірусів

Содержание
- 2. Список основної літератури: Вирусология. Методы. Под ред. Б. Мейхи. -М., Мир, 1988, -344 стр. И.П. Западнюк
- 3. Візуальна діагностика (?) Пряме вивчення (Direct Examination) Непряме вивчення (Virus Isolation) Серологія (?)
- 4. Опосередковані дослідження вірусу Методи виділеня, накопичення вірусів на різних модельних системах Виділення та дослідження нуклеїнової кислоти
- 5. Direct Examination 1. Детекція антигенів іммунофлуоресценція, ELISA etc. 2. Електронна мікроскопія морфологія віріонів імунна електронна мікроскпія
- 6. Indirect Examination 1. Культура клітин цитопатичний ефект (CPE) гемадсорбція іммунофлуоресценція 2. Курячі ембріони хоріоналантоїсна оболонка гемаглютинація
- 7. Серологічні методи досліджень
- 8. Візуальна діагностика За симтомами вірусних інфекцій людина тварини рослини бактерії ??
- 9. Симптоми вірусних інфекцій у людини Вірус віспи Вірус папіломи Вірус герпесу Вірус паротиту
- 10. Симптоми вірусних інфекцій у тварин а в с а, в – зовнішні сиптоми ураження вірусом ящуру
- 11. Симпотми ураження папіломавірусами у людини і тварин
- 12. From Medical Microbiology, 5th ed., Murray, Rosenthal & Pfaller, Mosby Inc., 2005, Table 51-1. Діагностика вірусних
- 13. Симптоми вірусної інфекції у рослин Вірус тютюнової мозаїки на томатах ВЖКЯ на озимій пшениці Вірус мозаїки
- 14. Утворення бляшок на газоні E.coli на чашці Петрі. бляшки малого діаметру – “дикий” фаг Т4 бляшки
- 15. Приклади модельних систем Модельні системи — системи, що використовуються як моделі для вивчення властивостей, процесів та
- 16. бактерія E. coli – бактеріофаг Т4
- 17. Arabidopsis thaliana 157 000 000 пнп, 5 хромосом, 25,498 генів.
- 18. Caenorhabditis elegans 100 000 000 пнп 20,000 генів
- 19. Використання тварин, рослин та бактерій для ідентифікації вірусів
- 20. Лабораторні тварини для безпосереднього виділення вірусів з оточуючого середовища; для виявлення (індикації) вірусу в патологічному матеріалі,
- 21. Використання курячих ембріонів Davis, Duylbecco, Eisen, Ginsberg “Microbiology” 4th ed, J.B. Lippincott 1990, Fig. 48-1
- 22. Первинна культура клітин
- 23. Субкультура фермент
- 24. Цитопатичний ефект Cytopathic effect of enterovirus 71 and HSV in cell culture: note the ballooning of
- 25. Використання рослин для ідентифікації вірусів Визначення: Рослини-індикатори – це рослини, які дають чітку специфічну реакцію на
- 26. Використання бактерій Негативні колонії (форма, розмір, ореол)
- 27. Світлова мікроскопія Заокруглення клітин у випадку зараження культури клітин вірусом простого герпесу
- 28. Електронна мікроскопія
- 29. Імуносорбентна електронна мікроскопія
- 30. Атомно-силова мікроскопія АСМ зображення фагу Т4 АСМ зображення ВТМ
- 31. Рентгено-структурний аналіз вірусів The phase problem preliminary model refined model The solving of the structure Ωκ={r:
- 32. Серологічні методи досліджень в вірусології Всі серологічні реакції базуються на специфічній взаємодії антигену з антитілом. З
- 33. Основні компоненти серологічних реакцій Антигени Антитіла (поліклональні та моноклональні)
- 34. Імунофлюоресцентні методи
- 36. Імунофлуоресцентний аналіз Виявлення АГ вірусу грипу типу А
- 37. Імуно-голд (Immunogold) електронна мікроскопія Антитіла до ВЖМЦ (вірусу жовтої мозаїки цукіні) кон’юговані колоїдним золотом
- 38. Імуноферментний аналіз Початком використання імуноферментних методик у вірусологічних дослідженнях вважають появу перших повідомлень про можливість приєднання
- 39. Компоненти ІФА Антиген Специфічні антитіла Антитіла специфічні мічені ферментом або антивидові антитіла, мічені ферментом Субстрат
- 40. Основні етапи ІФА (непрямого)
- 41. Прямий імуноферментний аналіз АГ ПХ
- 42. Непрямий імуноферментний аналіз АГ ПХ
- 43. Сендвіч - імуноферментний аналіз (сендвіч-метод) АГ ПХ
- 46. Імуноелектроблотинг (вестерн –блотинг)
- 47. Імуноелектроблотинг (вестерн –блотинг) 1 етап – електрофорез білків в денатуруючих умовах (за Леммлі) 2 етап –
- 48. Імуноелектроблотинг (вестерн –блотинг)
- 49. Молекулярно-генетичні методи детекції вірусів
- 50. Генетичні методи детекції Гібридизація нуклеїнових кислот (Саузерн,- нозерн–блотинг, гібридизація in situ) Рестрикційний аналіз Полімеразна ланцюгова реакція
- 51. Використання ПЛР ПЛР використовується не тільки для детекції аномальних генів та вірусів, але і для фундаментальних
- 52. Процес ампліфікації Процес ампліфікації складається з циклів, що повторюються: температурної денатурації ДНК, відпалу праймерів з комплементарними
- 53. 1й крок
- 54. 2й крок
- 55. 3й крок
- 56. Експоненційне збільшення ділянок ДНК, які утворюються за допомогою ПЛР. А= М x(2n-n-1) ≈ 2 n ,
- 57. Обладнання для ПЛР Реакції зазвичай проводять у 0,2 чи 0,5 мл мікропробірках Епендорф. Обладнання для нагрівання
- 58. Компоненти реакції Для проведення ПЛР необхідні такі складові частини: послідовність ДНК, що досліджується; буфер; дезоксирибонуклеозидтрифосфати (dNTP);
- 59. Послідовність ДНК, що досліджується, має бути попередньо підготована для аналізу (повинно бути проведено виділення нуклеїнової кислоти
- 60. Параметри температурних циклів ПЛР передбачає інкубацію зразків при трьох температурах, які відповідають трьом етапам циклу ампліфікації
- 61. Аналіз ПЛР-ампліфікованої ДНК Для аналізу продукту, отриманого в ПЛР, використовують різні методи, такі як: гель-електрофорез, дот-блот-гібридизацію
- 62. Діагностика вірусу сказу
- 63. Праймери, специфічні до послідовності гену капсидного білка ВШМЯ left - 5’GTGAGGAGGTGATGGGTAAT3’; right - 5’TTCCAGTCTTTCAGAATCTCTC3’ продукт ампліфікації
- 64. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
- 66. «nested» PCR Для збільшення чутливості та специфічності методу ПЛР була розроболена так звана «гніздова» полімеразна ланцюгова
- 67. «nested» PCR ДНК-мішень Перша пара праймерів Друга пара праймерів Амплікони Внутрішньої послідовності
- 68. 10 причин, з яких ПЛР може «не йти» Неякісний дизайн праймерів Невірна концентрація праймерів Занадто багато
- 69. Real-time PCR ПЛР з детекцією накопичення продуктів в режимі реального часу Основний принцип: Накопичення продуктів ПЛР
- 70. Детекція флуоресціюючого сигналу Ампліфікатор Флюориметр Випромінювач Детектор
- 71. Оптична система iQ5™ Збудження: вольфрам-галогенова лампа Детектування: 12-bit CCD камера
- 72. Real-time PCR Інтеркалюючі фарбники SYBR green Гібридизаційні зонди Taqman Molecular beacons FRET probes
- 73. Real-time PCR Інтеркалюючі фарбники SYBR green Гібридизаційні зонди Taqman Molecular beacons FRET probes
- 74. SYBR green Цей спосіб детекції заснований на тому факті, що флуоресценція бромистого етидію та SYBR Green
- 75. SYBR green Ампліфікація Інтеркаляція SYBR green олДНК длДНК
- 76. SYBR green
- 77. Van der Velden, Leukemia 2003 (www)
- 78. Real-time PCR Інтеркалюючі фарбники SYBR green Гібридизаційні зонди Taqman Molecular beacons FRET probes
- 79. PCR Real - Time На відміну від класичного ПЛР методу в реакції ПЛР в реальному часі
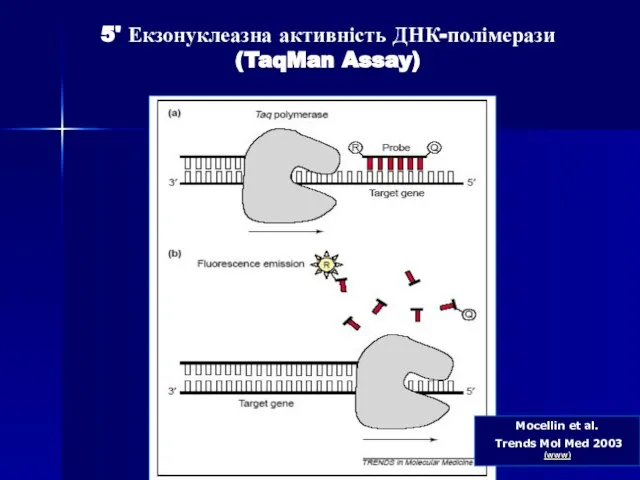
- 80. Mocellin et al. Trends Mol Med 2003 (www) 5' Екзонуклеазна активність ДНК-полімерази (TaqMan Assay)
- 81. Дана методика заснована на використанні 5'-екзонуклеазної активності полімерази. В реакційну суміш додають ДНК-зонди, в склад яких
- 82. В ході ПЛР під час стадії відпалу праймерів відбувається приєднання ДНК-зонду до комплементарного ланцюга ДНК, причому
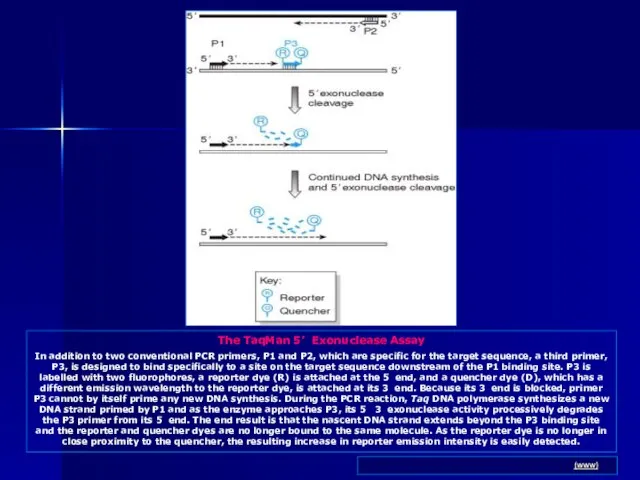
- 83. The TaqMan 5’ Exonuclease Assay In addition to two conventional PCR primers, P1 and P2, which
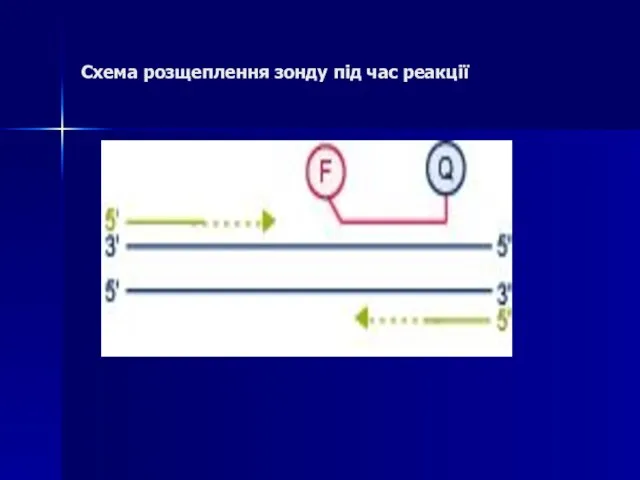
- 84. Схема розщеплення зонду під час реакції
- 85. Схема розщеплення зонду під час реакції
- 86. Схема розщеплення зонду під час реакції
- 87. Molecular beacons Дана методика відрізняється від попередньої (TaqMan Assay) тим, що кінцеві послідовності зонду являють собою
- 88. Mocellin et al. Trends Mol Med 2003 (www) Molecular beacons
- 89. Застосування 2-х зондів з резонансним переносом энергії (LightCycler assay) Даний спосіб детекції накопичення продуктів ампліфікації відрізняється
- 90. Застосування 2-х зондів з резонансним переносом энергії (LightCycler assay)
- 91. Застосування 2-х зондів з резонансним переносом энергії (LightCycler assay)
- 92. FRET = Förster/fluorescence resonance energy transfer ABI: Real-Time PCR vs Traditional PCR (www)
- 93. Nigel Walker, NIEHS (www)
- 94. ДНК-чіпи (DNA-array)
- 95. Технологія ДНК-чіпів
- 97. ДНК-чіп
- 98. + АСМ
- 99. Microarrays
- 101. Визначення філогенетичних зв’язків українських ізолятів вірусів рослин за допомогою порівняльного та філогенетичного аналізу ділянок гену білку
- 102. побудова філогенетичних дерев The time will come, I believe, though I shall not live to see
- 103. Реальні події : Дані: Побудоване дерево еволюція в природі чи в наприклад, дерево, лабораторії, н.к. послідо-
- 104. Поширення PPV в світі
- 105. ВШС (PPV, Potyvirus, Potyviridae) Гнучні палички без оболонки, близько 760 нм довжиною і 20 нм в
- 106. Геном ВШС Для діагностики вірусу шарки сливи використовували праймери специфічні до послідовності капсидного білка left- 5’
- 107. Електрофореграма продуктів RT-PCR в агарозному гелі по виявленню вірусу шарки сливи 1 – маркери (HyperLadder 1);
- 108. TGCATGTTGCGATTAACGTCACCAGCGGTGTGTCTCTCTGTGTCCTCTTCTTGTGTTCCGACGTTTCCATCCAAGCCAAATAAACGATTTTGAACATTTCTCAATGCTGCTGCCTTCATCTGGATATGAGCTTCACGTGCCCGTACGGGTGTCGTTGAAGTCATTTCGTAAAAATCAAAGGCATATCTGGCGAGGCTGTAGTCT Нуклеотидна послідовність амплікону, отриманого з інфікованих зразків сливи Сиквенс кДНК
- 109. Вигляд таблиці результатів вирівнювання нуклеотидних послідовностей гену капсидного білка з послідовностями генів капсидного білка, представлених в
- 110. Філогенетичне дерево нуклеотидних послідовностей капсидного білка українського ізоляту та відомих ізолятів ВШС, побудоване за методом NJ
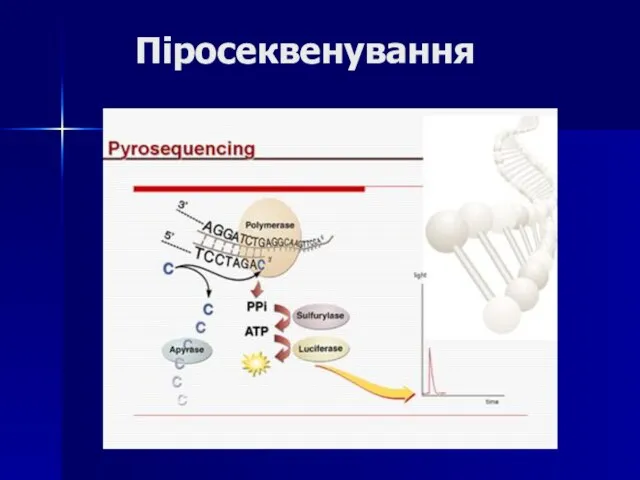
- 111. Піросеквенування
- 113. Скачать презентацию

Слайд 3
Візуальна діагностика (?)
Пряме вивчення (Direct Examination)
Непряме вивчення (Virus Isolation)
Серологія (?)
Візуальна діагностика (?)
Пряме вивчення (Direct Examination)
Непряме вивчення (Virus Isolation)
Серологія (?)

Слайд 4Опосередковані дослідження вірусу
Методи виділеня, накопичення вірусів на різних модельних системах
Виділення
Опосередковані дослідження вірусу
Методи виділеня, накопичення вірусів на різних модельних системах
Виділення

Слайд 5Direct Examination
1. Детекція антигенів іммунофлуоресценція, ELISA etc.
2. Електронна мікроскопія морфологія віріонів
Direct Examination
1. Детекція антигенів іммунофлуоресценція, ELISA etc.
2. Електронна мікроскопія морфологія віріонів

Слайд 6Indirect Examination
1. Культура клітин цитопатичний ефект (CPE)
гемадсорбція
іммунофлуоресценція
2. Курячі ембріони хоріоналантоїсна
Indirect Examination
1. Культура клітин цитопатичний ефект (CPE)
гемадсорбція
іммунофлуоресценція
2. Курячі ембріони хоріоналантоїсна

Слайд 7Серологічні методи досліджень
Серологічні методи досліджень

Слайд 8Візуальна діагностика
За симтомами вірусних інфекцій
людина
тварини
рослини
бактерії ??
Візуальна діагностика
За симтомами вірусних інфекцій
людина
тварини
рослини
бактерії ??

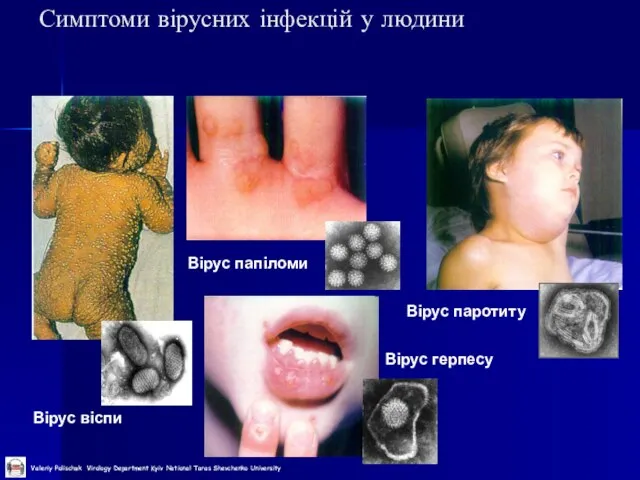
Слайд 9Симптоми вірусних інфекцій у людини
Вірус віспи
Вірус папіломи
Вірус герпесу
Вірус паротиту
Симптоми вірусних інфекцій у людини
Вірус віспи
Вірус папіломи
Вірус герпесу
Вірус паротиту
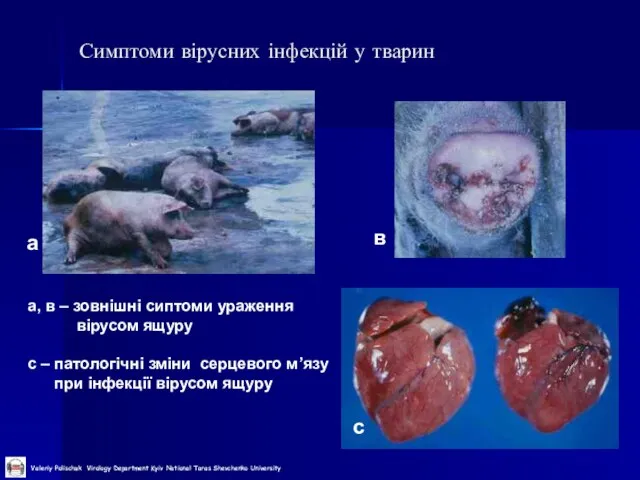
Слайд 10Симптоми вірусних інфекцій у тварин
а
в
с
а, в – зовнішні сиптоми ураження
вірусом ящуру
с
Симптоми вірусних інфекцій у тварин
а
в
с
а, в – зовнішні сиптоми ураження
вірусом ящуру
с
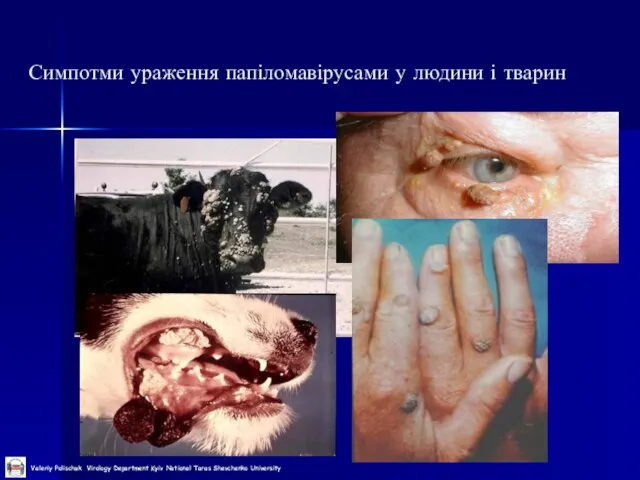
Слайд 11Симпотми ураження папіломавірусами у людини і тварин
Симпотми ураження папіломавірусами у людини і тварин
Слайд 12From Medical Microbiology, 5th ed., Murray, Rosenthal & Pfaller, Mosby Inc., 2005,
From Medical Microbiology, 5th ed., Murray, Rosenthal & Pfaller, Mosby Inc., 2005,

Слайд 13Симптоми вірусної інфекції у рослин
Вірус тютюнової мозаїки на томатах
ВЖКЯ на озимій
Симптоми вірусної інфекції у рослин
Вірус тютюнової мозаїки на томатах
ВЖКЯ на озимій

Слайд 14
Утворення бляшок на газоні E.coli на чашці Петрі.
бляшки малого
Утворення бляшок на газоні E.coli на чашці Петрі.
бляшки малого
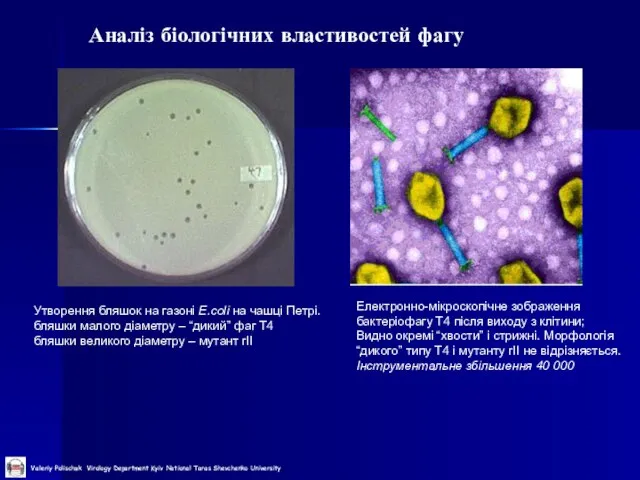
Слайд 15Приклади модельних систем
Модельні системи — системи, що використовуються як моделі для вивчення
Приклади модельних систем
Модельні системи — системи, що використовуються як моделі для вивчення

Слайд 16
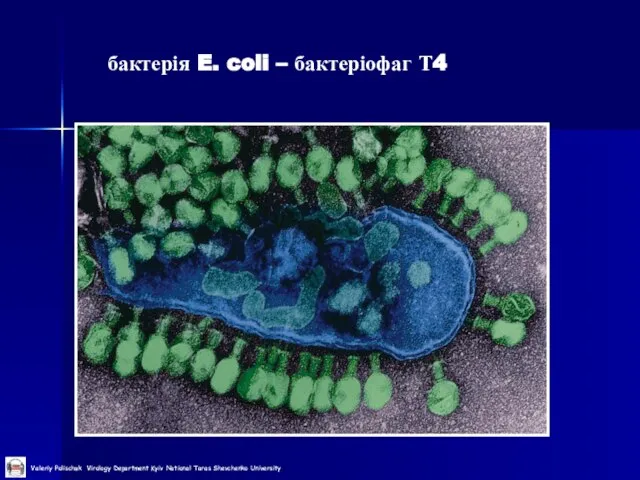
бактерія E. coli – бактеріофаг Т4
бактерія E. coli – бактеріофаг Т4
Слайд 17
Arabidopsis thaliana
157 000 000 пнп, 5 хромосом, 25,498 генів.
Arabidopsis thaliana
157 000 000 пнп, 5 хромосом, 25,498 генів.

Слайд 18
Caenorhabditis elegans
100 000 000 пнп 20,000 генів
Caenorhabditis elegans
100 000 000 пнп 20,000 генів

Слайд 19
Використання тварин, рослин та бактерій для ідентифікації вірусів
Використання тварин, рослин та бактерій для ідентифікації вірусів

Слайд 20Лабораторні тварини
для безпосереднього виділення вірусів з оточуючого середовища;
для виявлення (індикації) вірусу в
Лабораторні тварини
для безпосереднього виділення вірусів з оточуючого середовища;
для виявлення (індикації) вірусу в

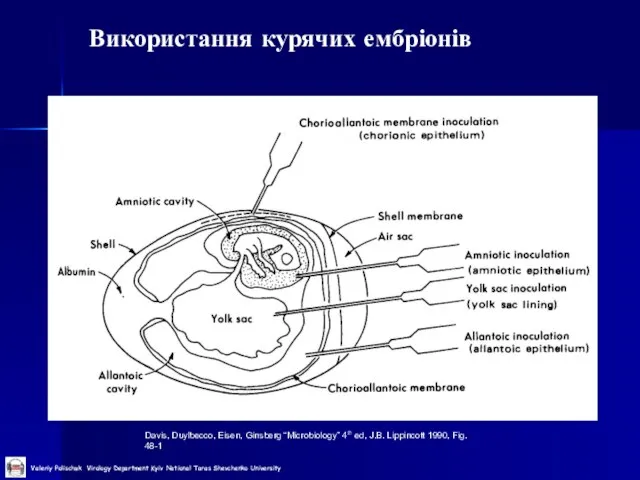
Слайд 21Використання курячих ембріонів
Davis, Duylbecco, Eisen, Ginsberg “Microbiology” 4th ed, J.B. Lippincott 1990,
Використання курячих ембріонів
Davis, Duylbecco, Eisen, Ginsberg “Microbiology” 4th ed, J.B. Lippincott 1990,
Слайд 22Первинна культура клітин
Первинна культура клітин
Слайд 23Субкультура
фермент
Субкультура
фермент
Слайд 24Цитопатичний ефект
Cytopathic effect of enterovirus 71 and HSV in cell culture: note
Цитопатичний ефект
Cytopathic effect of enterovirus 71 and HSV in cell culture: note
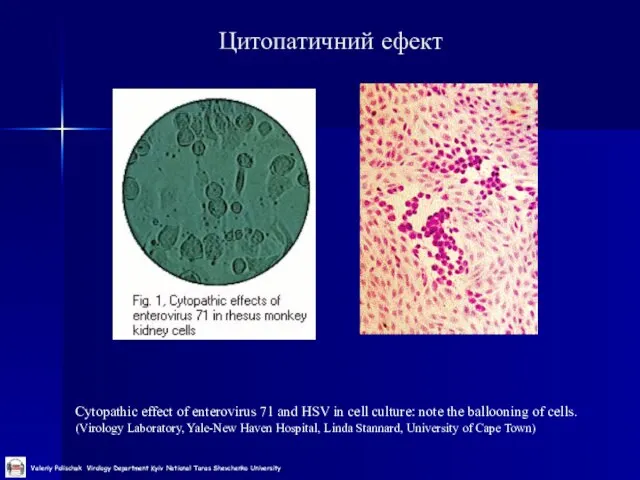
Слайд 25Використання рослин
для ідентифікації вірусів
Визначення:
Рослини-індикатори – це рослини, які дають чітку
Використання рослин
для ідентифікації вірусів
Визначення:
Рослини-індикатори – це рослини, які дають чітку

Слайд 26Використання бактерій
Негативні колонії (форма, розмір, ореол)
Використання бактерій
Негативні колонії (форма, розмір, ореол)
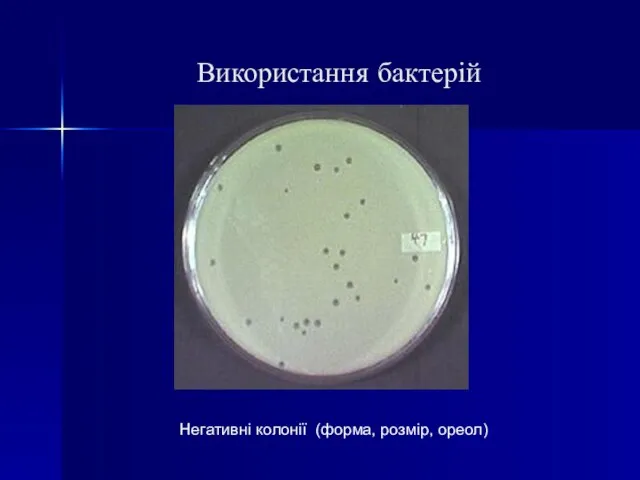
Слайд 27Світлова мікроскопія
Заокруглення клітин у випадку зараження культури клітин вірусом простого герпесу
Світлова мікроскопія
Заокруглення клітин у випадку зараження культури клітин вірусом простого герпесу
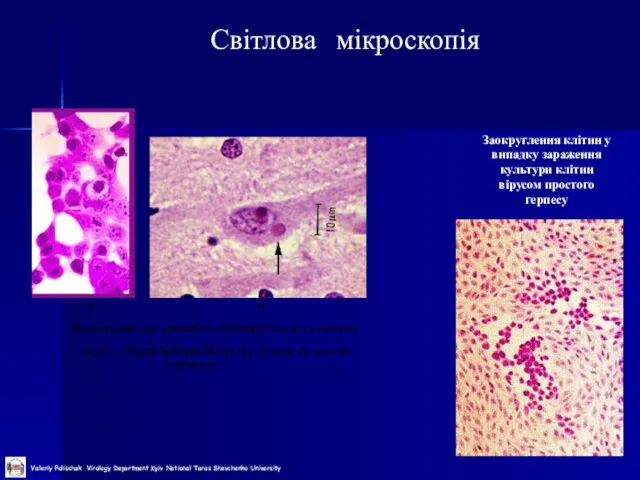
Слайд 28Електронна мікроскопія
Електронна мікроскопія
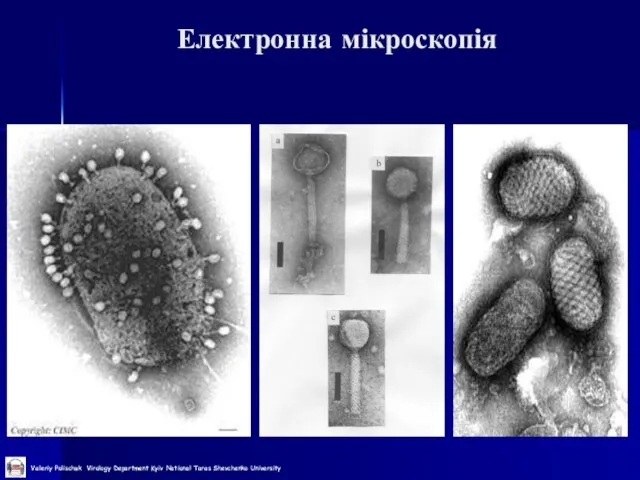
Слайд 29
Імуносорбентна електронна мікроскопія
Імуносорбентна електронна мікроскопія
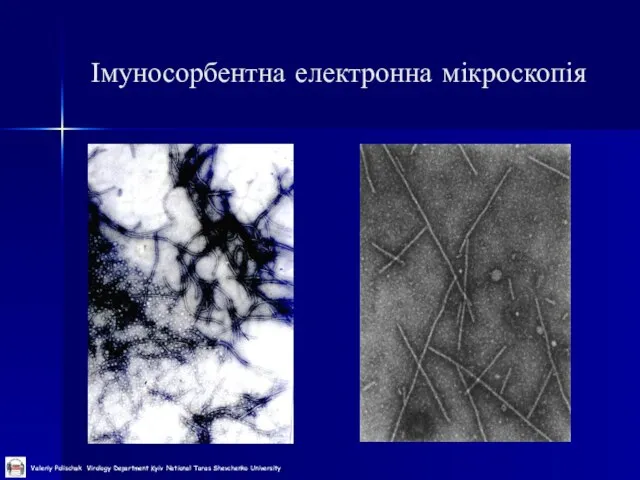
Слайд 30Атомно-силова мікроскопія
АСМ зображення фагу Т4
АСМ зображення ВТМ
Атомно-силова мікроскопія
АСМ зображення фагу Т4
АСМ зображення ВТМ

Слайд 31Рентгено-структурний аналіз вірусів
The phase problem
preliminary model
refined model
The solving of
the structure
Ωκ={r: ρS(r)
Рентгено-структурний аналіз вірусів
The phase problem
preliminary model
refined model
The solving of
the structure
Ωκ={r: ρS(r)
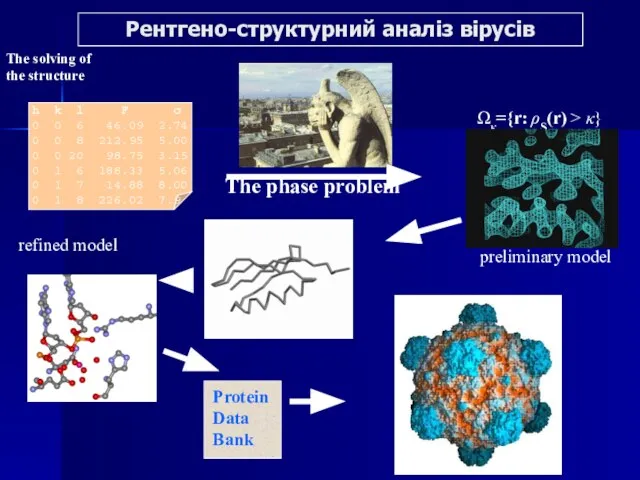
Слайд 32Серологічні методи
досліджень
в вірусології
Всі серологічні реакції базуються на специфічній взаємодії антигену
Серологічні методи
досліджень
в вірусології
Всі серологічні реакції базуються на специфічній взаємодії антигену

Слайд 33Основні компоненти серологічних реакцій
Антигени
Антитіла (поліклональні та моноклональні)
Основні компоненти серологічних реакцій
Антигени
Антитіла (поліклональні та моноклональні)

Слайд 34Імунофлюоресцентні методи
Імунофлюоресцентні методи
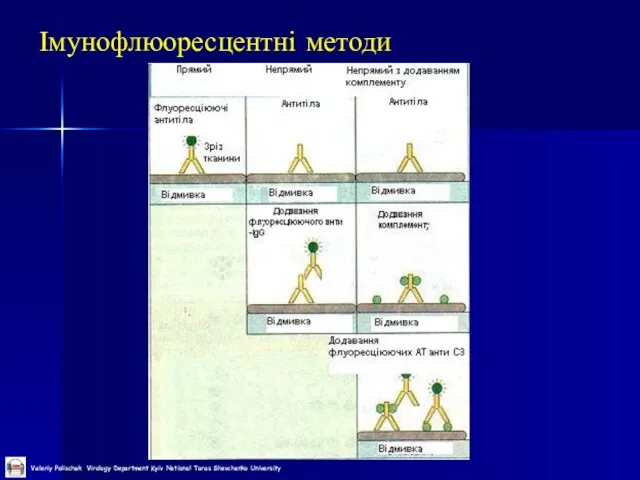
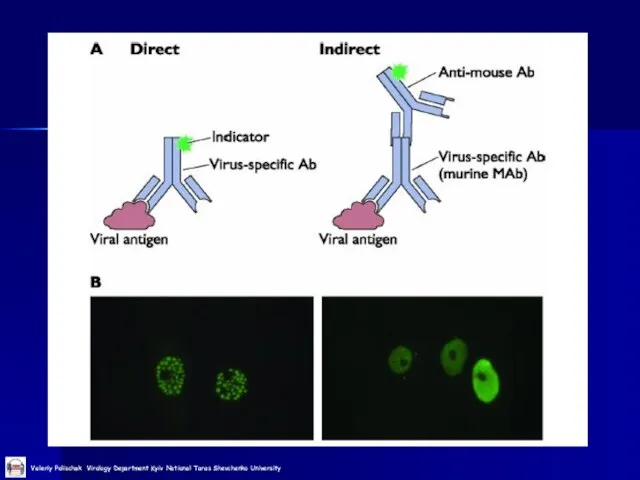
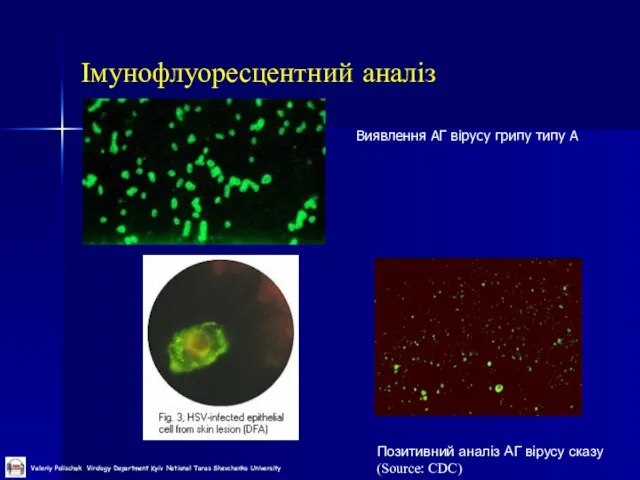
Слайд 36Імунофлуоресцентний аналіз
Виявлення АГ вірусу грипу типу А
Імунофлуоресцентний аналіз
Виявлення АГ вірусу грипу типу А
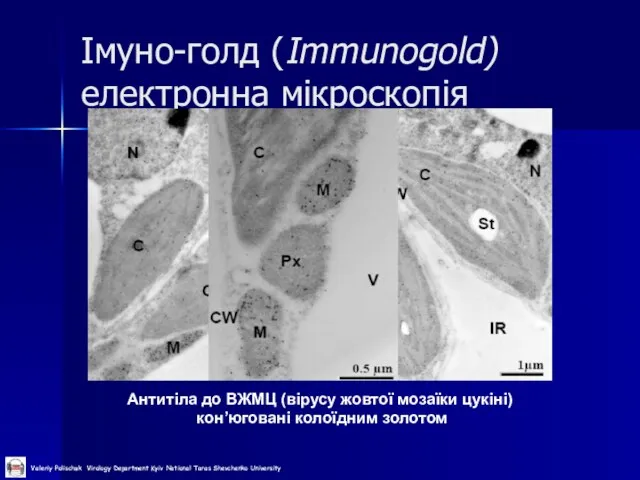
Слайд 37Імуно-голд (Immunogold) електронна мікроскопія
Антитіла до ВЖМЦ (вірусу жовтої мозаїки цукіні)
кон’юговані
Імуно-голд (Immunogold) електронна мікроскопія
Антитіла до ВЖМЦ (вірусу жовтої мозаїки цукіні)
кон’юговані
Слайд 38Імуноферментний аналіз
Початком використання імуноферментних методик у вірусологічних дослідженнях вважають появу перших повідомлень
Імуноферментний аналіз
Початком використання імуноферментних методик у вірусологічних дослідженнях вважають появу перших повідомлень

Слайд 39Компоненти ІФА
Антиген
Специфічні антитіла
Антитіла специфічні мічені ферментом або антивидові антитіла, мічені ферментом
Субстрат
Компоненти ІФА
Антиген
Специфічні антитіла
Антитіла специфічні мічені ферментом або антивидові антитіла, мічені ферментом
Субстрат

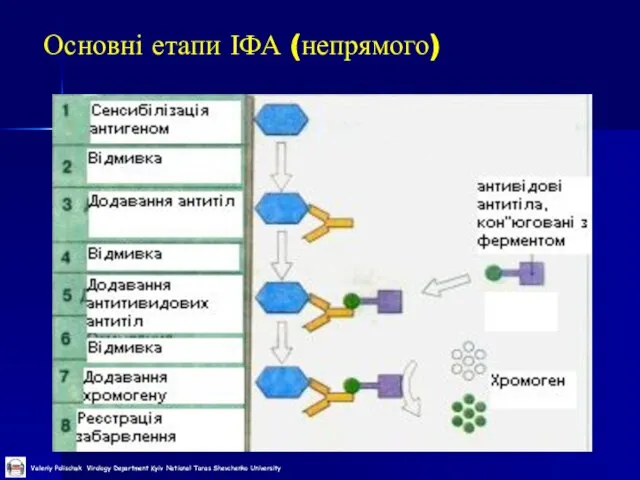
Слайд 40Основні етапи ІФА (непрямого)
Основні етапи ІФА (непрямого)
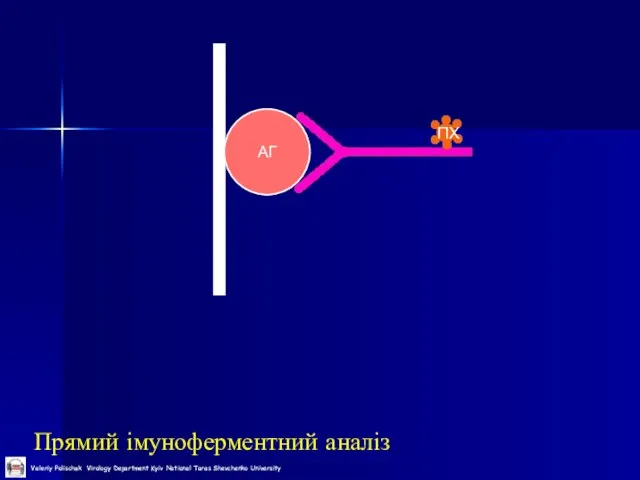
Слайд 41Прямий імуноферментний аналіз
АГ
ПХ
Прямий імуноферментний аналіз
АГ
ПХ
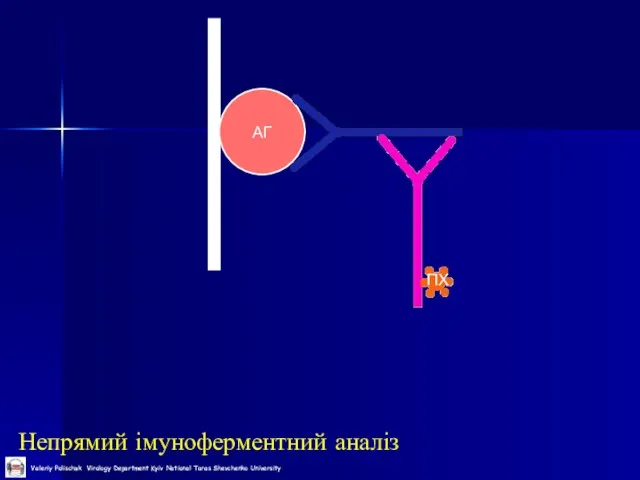
Слайд 42Непрямий імуноферментний аналіз
АГ
ПХ
Непрямий імуноферментний аналіз
АГ
ПХ
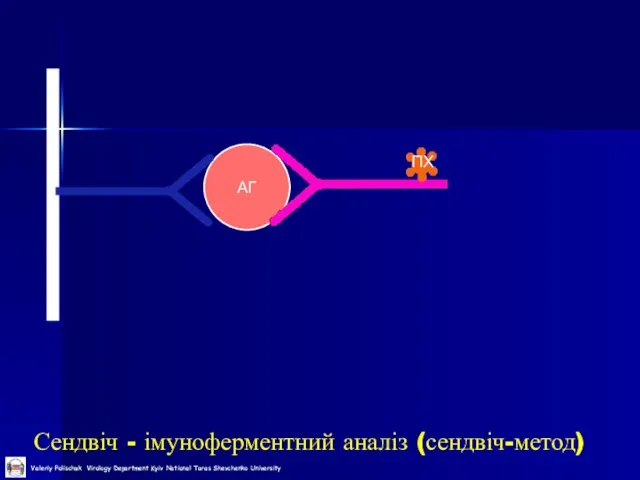
Слайд 43Сендвіч - імуноферментний аналіз (сендвіч-метод)
АГ
ПХ
Сендвіч - імуноферментний аналіз (сендвіч-метод)
АГ
ПХ
Слайд 46Імуноелектроблотинг (вестерн –блотинг)
Імуноелектроблотинг (вестерн –блотинг)
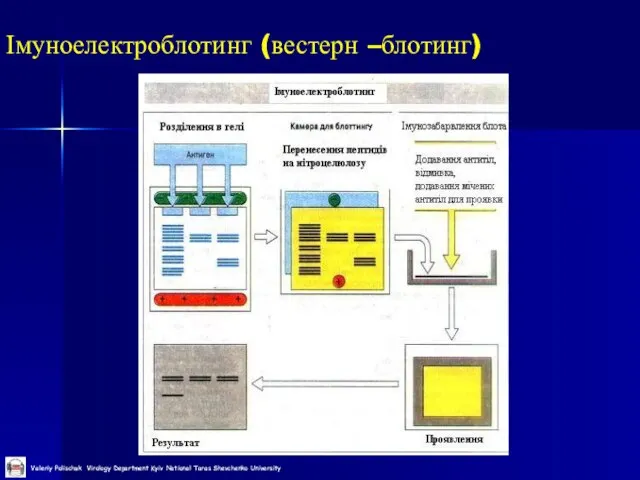
Слайд 47Імуноелектроблотинг (вестерн –блотинг)
1 етап – електрофорез білків в денатуруючих умовах (за Леммлі)
2
Імуноелектроблотинг (вестерн –блотинг)
1 етап – електрофорез білків в денатуруючих умовах (за Леммлі)
2

Слайд 48
Імуноелектроблотинг (вестерн –блотинг)
Імуноелектроблотинг (вестерн –блотинг)
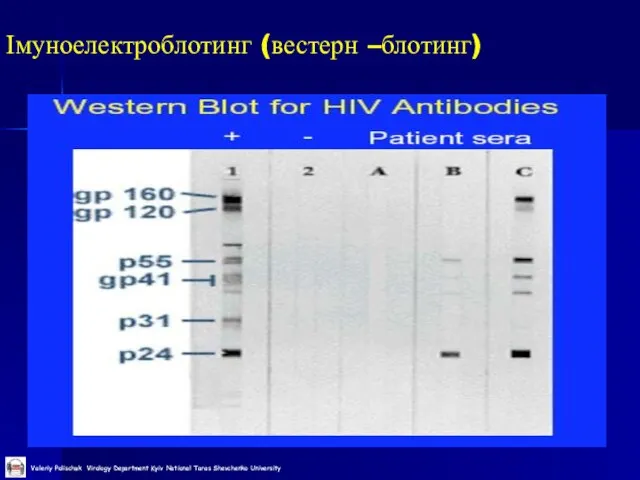
Слайд 49Молекулярно-генетичні методи детекції вірусів
Молекулярно-генетичні методи детекції вірусів

Слайд 50Генетичні методи детекції
Гібридизація нуклеїнових кислот (Саузерн,- нозерн–блотинг, гібридизація in situ)
Рестрикційний аналіз
Полімеразна ланцюгова
Генетичні методи детекції
Гібридизація нуклеїнових кислот (Саузерн,- нозерн–блотинг, гібридизація in situ)
Рестрикційний аналіз
Полімеразна ланцюгова

Слайд 51Використання ПЛР
ПЛР використовується не тільки для детекції аномальних генів та вірусів, але
Використання ПЛР
ПЛР використовується не тільки для детекції аномальних генів та вірусів, але

Слайд 52Процес ампліфікації
Процес ампліфікації складається з циклів, що повторюються:
температурної денатурації ДНК,
відпалу праймерів
Процес ампліфікації
Процес ампліфікації складається з циклів, що повторюються:
температурної денатурації ДНК,
відпалу праймерів

Слайд 53
1й крок
1й крок
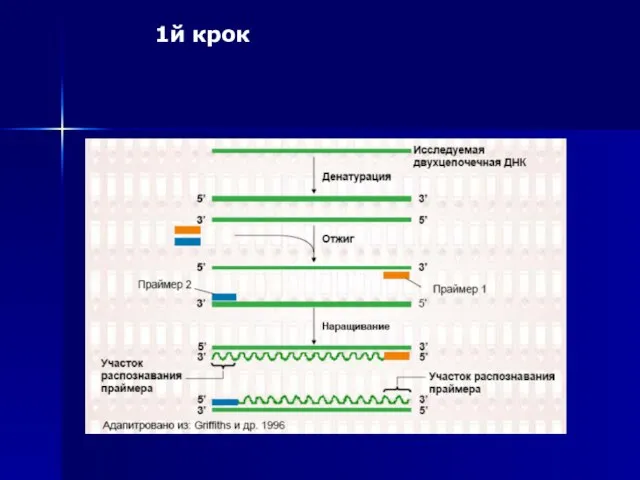
Слайд 54
2й крок
2й крок
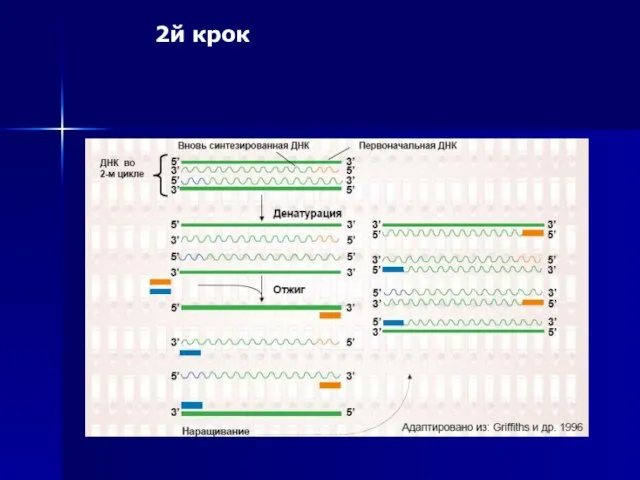
Слайд 55
3й крок
3й крок
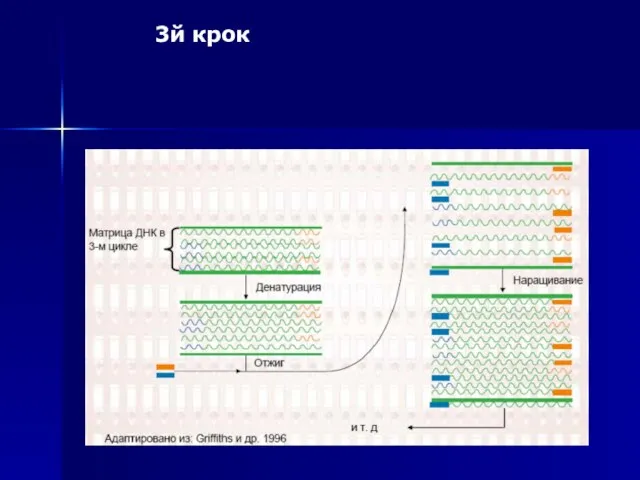
Слайд 56Експоненційне збільшення ділянок ДНК, які утворюються за допомогою ПЛР.
А= М x(2n-n-1) ≈
Експоненційне збільшення ділянок ДНК, які утворюються за допомогою ПЛР.
А= М x(2n-n-1) ≈

Слайд 57Обладнання для ПЛР
Реакції зазвичай проводять у 0,2 чи 0,5 мл мікропробірках Епендорф.
Обладнання для ПЛР
Реакції зазвичай проводять у 0,2 чи 0,5 мл мікропробірках Епендорф.

Слайд 58Компоненти реакції
Для проведення ПЛР необхідні такі складові частини:
послідовність ДНК, що досліджується;
буфер;
дезоксирибонуклеозидтрифосфати
Компоненти реакції
Для проведення ПЛР необхідні такі складові частини:
послідовність ДНК, що досліджується;
буфер;
дезоксирибонуклеозидтрифосфати

Слайд 59Послідовність ДНК, що досліджується, має бути попередньо підготована для аналізу (повинно бути
Послідовність ДНК, що досліджується, має бути попередньо підготована для аналізу (повинно бути

Слайд 60Параметри температурних циклів
ПЛР передбачає інкубацію зразків при трьох температурах, які відповідають трьом
Параметри температурних циклів
ПЛР передбачає інкубацію зразків при трьох температурах, які відповідають трьом

Слайд 61Аналіз ПЛР-ампліфікованої ДНК
Для аналізу продукту, отриманого в ПЛР, використовують різні методи, такі
Аналіз ПЛР-ампліфікованої ДНК
Для аналізу продукту, отриманого в ПЛР, використовують різні методи, такі

Слайд 62Діагностика вірусу сказу
Діагностика вірусу сказу
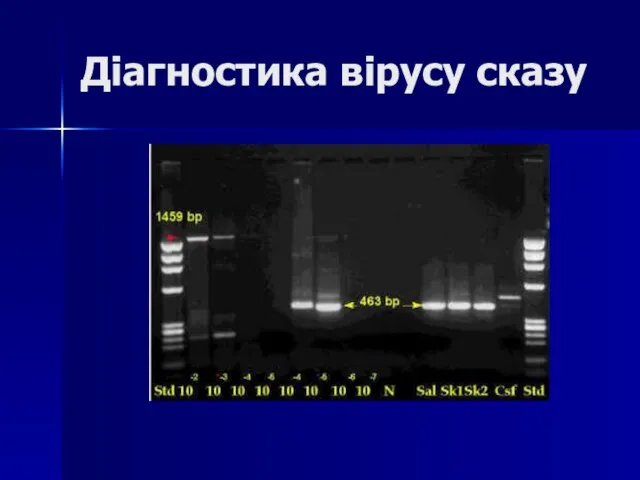
Слайд 63Праймери, специфічні до послідовності гену капсидного білка ВШМЯ
left - 5’GTGAGGAGGTGATGGGTAAT3’;
right -
Праймери, специфічні до послідовності гену капсидного білка ВШМЯ
left - 5’GTGAGGAGGTGATGGGTAAT3’;
right -

Слайд 64 1 2 3 4 5 6 7 8 9 10 11
1 2 3 4 5 6 7 8 9 10 11


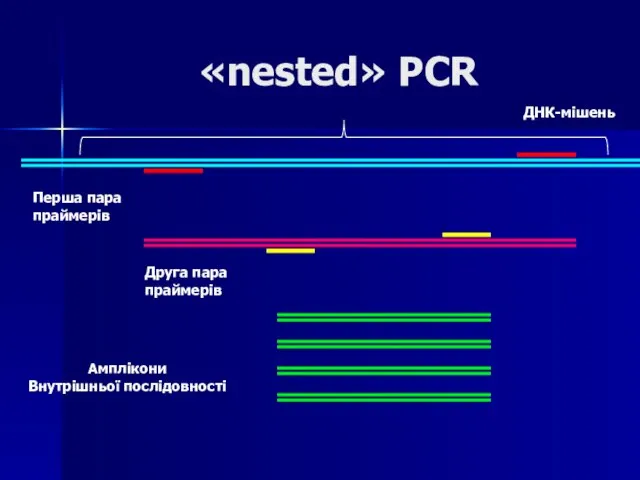
Слайд 66«nested» PCR
Для збільшення чутливості та специфічності методу ПЛР була розроболена так звана
«nested» PCR
Для збільшення чутливості та специфічності методу ПЛР була розроболена так звана

Слайд 67«nested» PCR
ДНК-мішень
Перша пара
праймерів
Друга пара
праймерів
Амплікони
Внутрішньої послідовності
«nested» PCR
ДНК-мішень
Перша пара
праймерів
Друга пара
праймерів
Амплікони
Внутрішньої послідовності
Слайд 6810 причин,
з яких ПЛР може «не йти»
Неякісний дизайн праймерів
Невірна концентрація праймерів
Занадто
10 причин,
з яких ПЛР може «не йти»
Неякісний дизайн праймерів
Невірна концентрація праймерів
Занадто

Слайд 69Real-time PCR
ПЛР з детекцією накопичення продуктів
в режимі реального часу
Основний принцип:
Накопичення
Real-time PCR
ПЛР з детекцією накопичення продуктів
в режимі реального часу
Основний принцип:
Накопичення

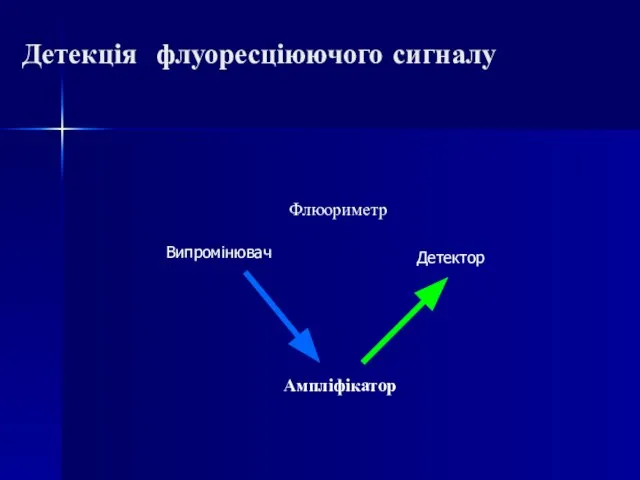
Слайд 70Детекція флуоресціюючого сигналу
Ампліфікатор
Флюориметр
Випромінювач
Детектор
Детекція флуоресціюючого сигналу
Ампліфікатор
Флюориметр
Випромінювач
Детектор
Слайд 71Оптична система iQ5™
Збудження: вольфрам-галогенова лампа
Детектування: 12-bit CCD камера
Оптична система iQ5™
Збудження: вольфрам-галогенова лампа
Детектування: 12-bit CCD камера

Слайд 72Real-time PCR
Інтеркалюючі фарбники
SYBR green
Гібридизаційні зонди
Taqman
Molecular beacons
FRET probes
Real-time PCR
Інтеркалюючі фарбники
SYBR green
Гібридизаційні зонди
Taqman
Molecular beacons
FRET probes

Слайд 73Real-time PCR
Інтеркалюючі фарбники
SYBR green
Гібридизаційні зонди
Taqman
Molecular beacons
FRET probes
Real-time PCR
Інтеркалюючі фарбники
SYBR green
Гібридизаційні зонди
Taqman
Molecular beacons
FRET probes

Слайд 74SYBR green
Цей спосіб детекції заснований на тому факті, що флуоресценція бромистого етидію
SYBR green
Цей спосіб детекції заснований на тому факті, що флуоресценція бромистого етидію

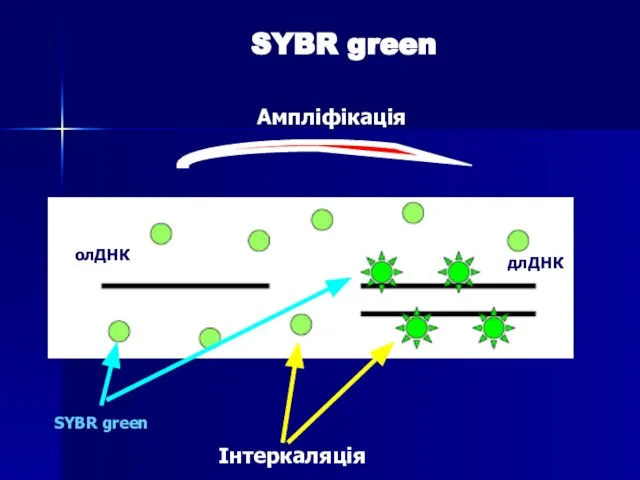
Слайд 75SYBR green
Ампліфікація
Інтеркаляція
SYBR green
олДНК
длДНК
SYBR green
Ампліфікація
Інтеркаляція
SYBR green
олДНК
длДНК
Слайд 76
SYBR green
SYBR green
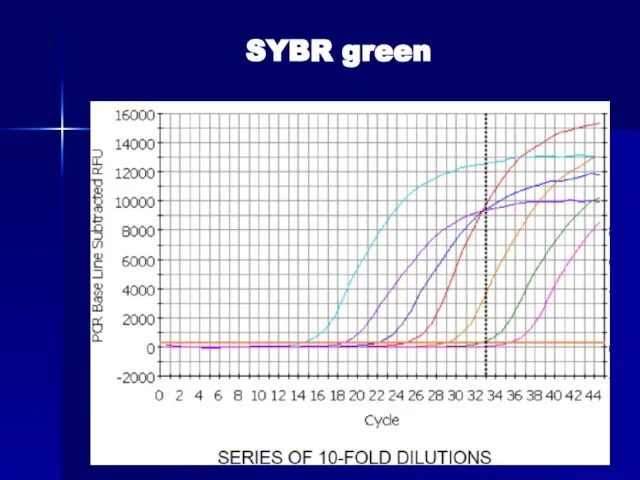
Слайд 77Van der Velden, Leukemia 2003 (www)
Van der Velden, Leukemia 2003 (www)
Слайд 78Real-time PCR
Інтеркалюючі фарбники
SYBR green
Гібридизаційні зонди
Taqman
Molecular beacons
FRET probes
Real-time PCR
Інтеркалюючі фарбники
SYBR green
Гібридизаційні зонди
Taqman
Molecular beacons
FRET probes

Слайд 79PCR Real - Time
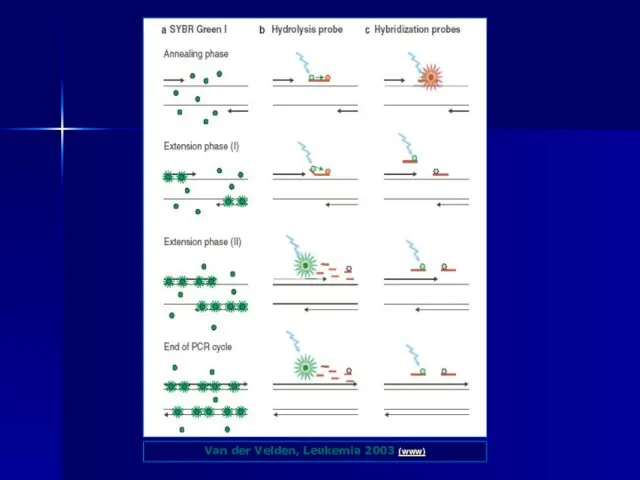
На відміну від класичного ПЛР методу в реакції ПЛР
PCR Real - Time
На відміну від класичного ПЛР методу в реакції ПЛР
Слайд 80Mocellin et al.
Trends Mol Med 2003 (www)
5' Екзонуклеазна активність ДНК-полімерази (TaqMan
Mocellin et al.
Trends Mol Med 2003 (www)
5' Екзонуклеазна активність ДНК-полімерази (TaqMan
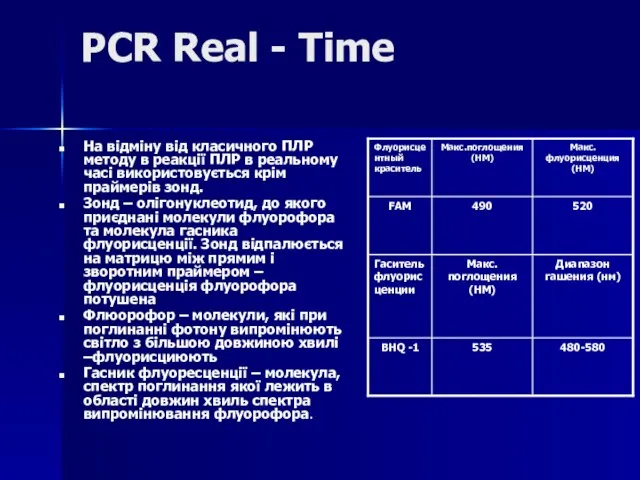
Слайд 81Дана методика заснована на використанні 5'-екзонуклеазної активності полімерази. В реакційну суміш додають
Дана методика заснована на використанні 5'-екзонуклеазної активності полімерази. В реакційну суміш додають

Слайд 82В ході ПЛР під час стадії відпалу праймерів відбувається приєднання ДНК-зонду до
В ході ПЛР під час стадії відпалу праймерів відбувається приєднання ДНК-зонду до

Слайд 83The TaqMan 5’ Exonuclease Assay
In addition to two conventional PCR primers, P1
The TaqMan 5’ Exonuclease Assay
In addition to two conventional PCR primers, P1
Слайд 84Схема розщеплення зонду під час реакції
Схема розщеплення зонду під час реакції
Слайд 85
Схема розщеплення зонду під час реакції
Схема розщеплення зонду під час реакції

Слайд 86Схема розщеплення зонду під час реакції
Схема розщеплення зонду під час реакції

Слайд 87Molecular beacons
Дана методика відрізняється від попередньої (TaqMan Assay) тим, що кінцеві послідовності
Molecular beacons
Дана методика відрізняється від попередньої (TaqMan Assay) тим, що кінцеві послідовності

Слайд 88Mocellin et al. Trends Mol Med 2003 (www)
Molecular beacons
Mocellin et al. Trends Mol Med 2003 (www)
Molecular beacons
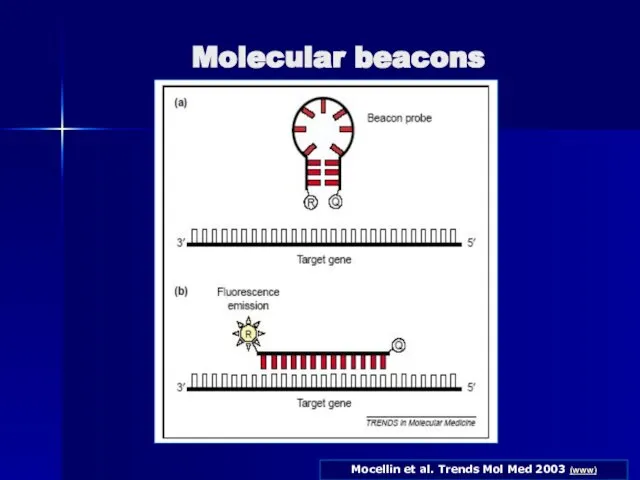
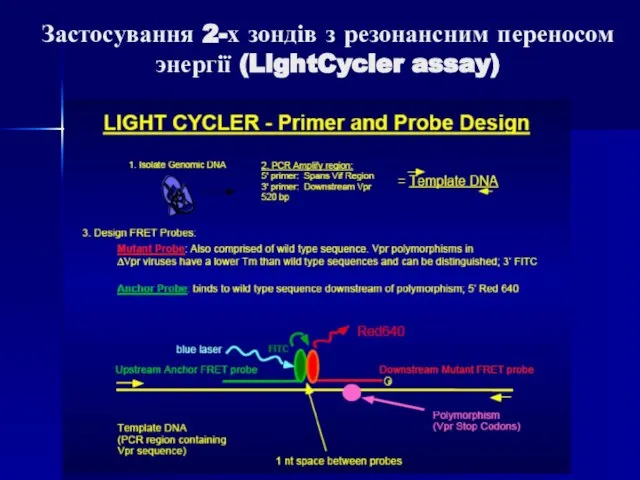
Слайд 89Застосування 2-х зондів з резонансним переносом энергії (LightCycler assay)
Даний спосіб детекції накопичення
Застосування 2-х зондів з резонансним переносом энергії (LightCycler assay)
Даний спосіб детекції накопичення

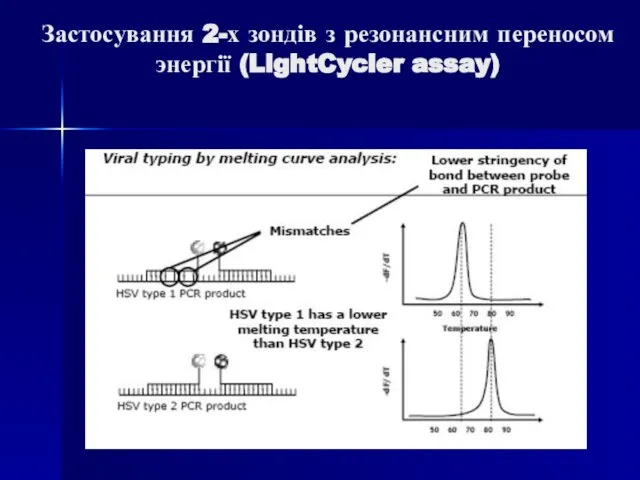
Слайд 90Застосування 2-х зондів з резонансним переносом энергії (LightCycler assay)
Застосування 2-х зондів з резонансним переносом энергії (LightCycler assay)
Слайд 91Застосування 2-х зондів з резонансним переносом энергії (LightCycler assay)
Застосування 2-х зондів з резонансним переносом энергії (LightCycler assay)
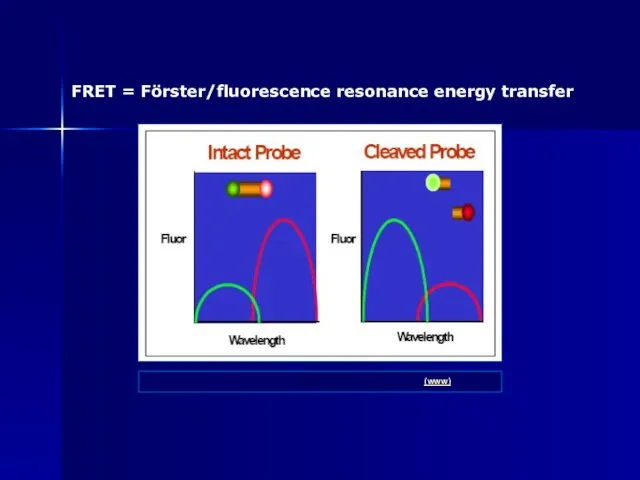
Слайд 92FRET = Förster/fluorescence resonance energy transfer
ABI: Real-Time PCR vs Traditional PCR
FRET = Förster/fluorescence resonance energy transfer
ABI: Real-Time PCR vs Traditional PCR
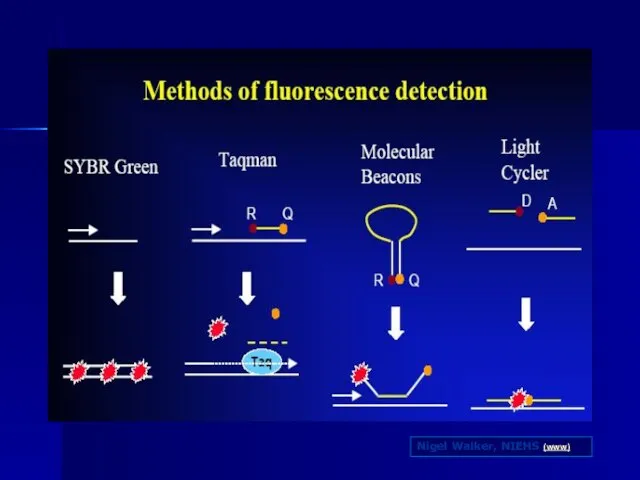
Слайд 93Nigel Walker, NIEHS (www)
Nigel Walker, NIEHS (www)
Слайд 94
ДНК-чіпи
(DNA-array)
ДНК-чіпи
(DNA-array)

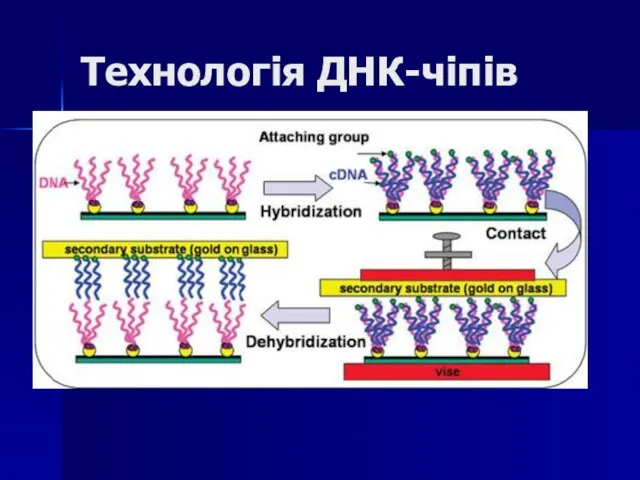
Слайд 95Технологія ДНК-чіпів
Технологія ДНК-чіпів
Слайд 97ДНК-чіп
ДНК-чіп

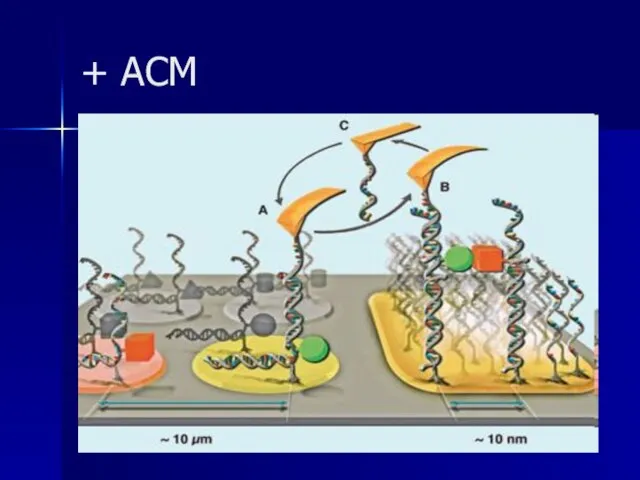
Слайд 98+ АСМ
+ АСМ
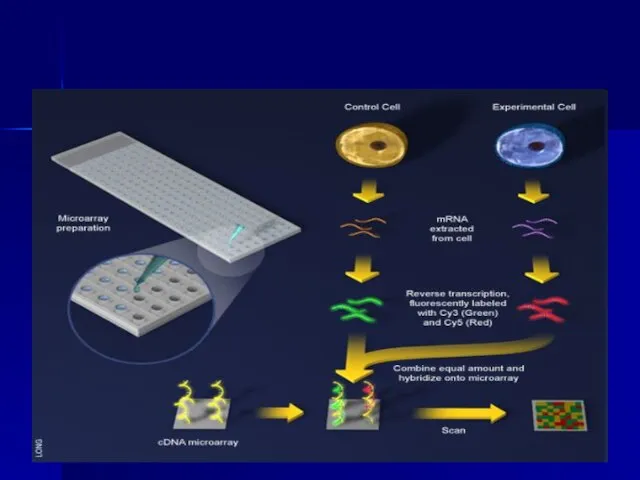
Слайд 99Microarrays
Microarrays


Слайд 101
Визначення філогенетичних зв’язків українських ізолятів вірусів рослин за допомогою порівняльного та
Визначення філогенетичних зв’язків українських ізолятів вірусів рослин за допомогою порівняльного та

Слайд 102 побудова філогенетичних дерев
The time will come, I believe, though I shall
побудова філогенетичних дерев
The time will come, I believe, though I shall

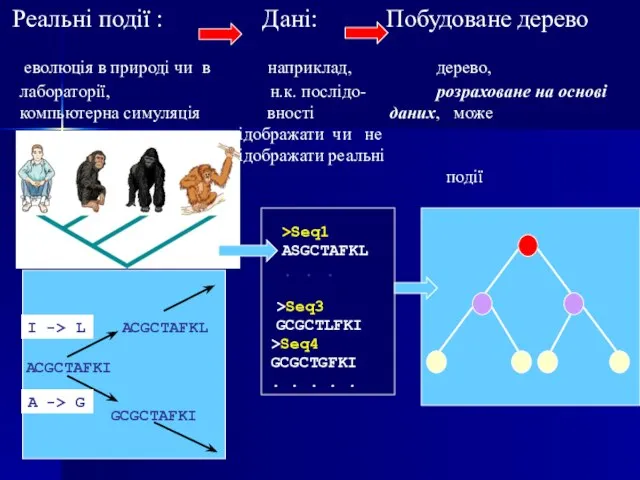
Слайд 103 Реальні події : Дані: Побудоване дерево
еволюція в природі чи
Реальні події : Дані: Побудоване дерево
еволюція в природі чи
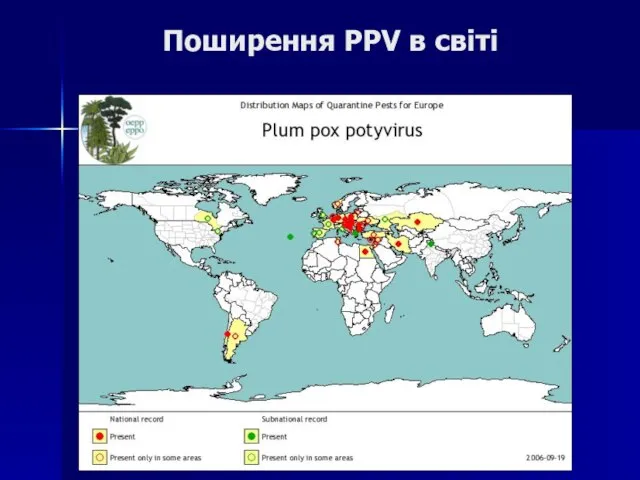
Слайд 104Поширення PPV в світі
Поширення PPV в світі
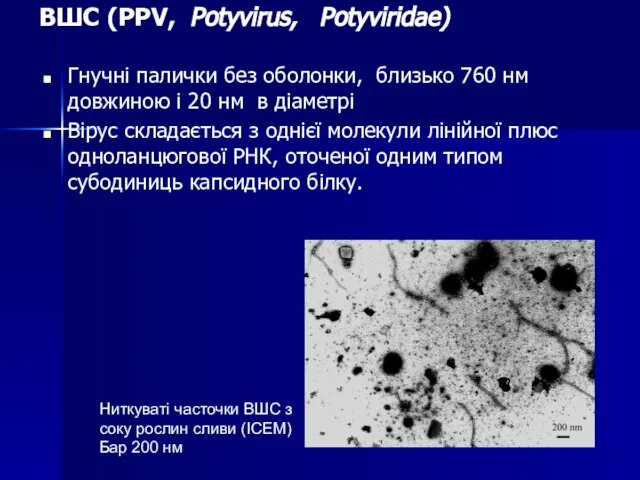
Слайд 105ВШС (PPV, Potyvirus, Potyviridae)
Гнучні палички без оболонки, близько 760 нм довжиною і
ВШС (PPV, Potyvirus, Potyviridae)
Гнучні палички без оболонки, близько 760 нм довжиною і
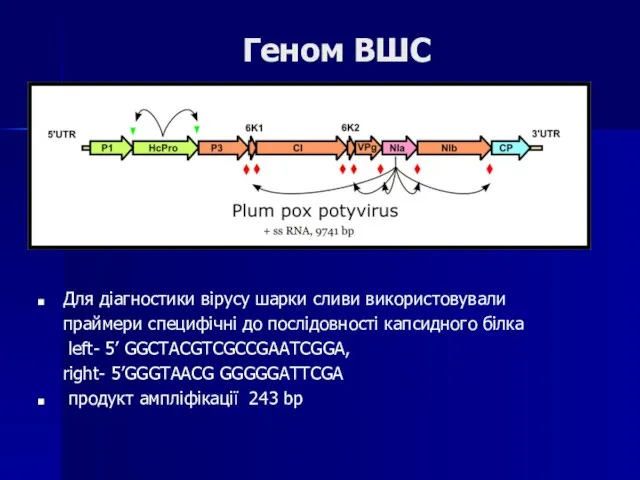
Слайд 106Геном ВШС
Для діагностики вірусу шарки сливи використовували
праймери специфічні до послідовності капсидного білка
Геном ВШС
Для діагностики вірусу шарки сливи використовували
праймери специфічні до послідовності капсидного білка
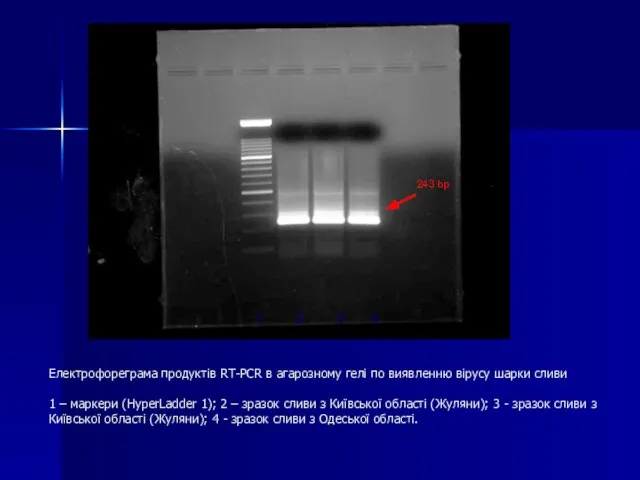
Слайд 107Електрофореграма продуктів RT-PCR в агарозному гелі по виявленню вірусу шарки сливи
1 –
Електрофореграма продуктів RT-PCR в агарозному гелі по виявленню вірусу шарки сливи 1 –
Слайд 108 TGCATGTTGCGATTAACGTCACCAGCGGTGTGTCTCTCTGTGTCCTCTTCTTGTGTTCCGACGTTTCCATCCAAGCCAAATAAACGATTTTGAACATTTCTCAATGCTGCTGCCTTCATCTGGATATGAGCTTCACGTGCCCGTACGGGTGTCGTTGAAGTCATTTCGTAAAAATCAAAGGCATATCTGGCGAGGCTGTAGTCT
Нуклеотидна послідовність амплікону, отриманого з інфікованих зразків сливи
Сиквенс кДНК
TGCATGTTGCGATTAACGTCACCAGCGGTGTGTCTCTCTGTGTCCTCTTCTTGTGTTCCGACGTTTCCATCCAAGCCAAATAAACGATTTTGAACATTTCTCAATGCTGCTGCCTTCATCTGGATATGAGCTTCACGTGCCCGTACGGGTGTCGTTGAAGTCATTTCGTAAAAATCAAAGGCATATCTGGCGAGGCTGTAGTCT
Нуклеотидна послідовність амплікону, отриманого з інфікованих зразків сливи
Сиквенс кДНК

Слайд 109
Вигляд таблиці результатів вирівнювання нуклеотидних послідовностей гену капсидного білка з послідовностями
Вигляд таблиці результатів вирівнювання нуклеотидних послідовностей гену капсидного білка з послідовностями

Слайд 110Філогенетичне дерево нуклеотидних послідовностей капсидного білка українського ізоляту та відомих ізолятів ВШС,
Філогенетичне дерево нуклеотидних послідовностей капсидного білка українського ізоляту та відомих ізолятів ВШС,

Слайд 111Піросеквенування
Піросеквенування
 Архитектура и функционирование персонального компьютера
Архитектура и функционирование персонального компьютера Каталог TopFranchise. Коммерческое предложение по сотрудничеству
Каталог TopFranchise. Коммерческое предложение по сотрудничеству Лот 16, г. Хабаровск, ул. Сысоева, 21, кв. 32
Лот 16, г. Хабаровск, ул. Сысоева, 21, кв. 32 Тема. Предмет исследования (шаблон)
Тема. Предмет исследования (шаблон) Воссоздающее и творческое воображение
Воссоздающее и творческое воображение Специальные звания и сроки выслуги
Специальные звания и сроки выслуги Федеральный государственный образовательный стандарт основного общего образования
Федеральный государственный образовательный стандарт основного общего образования Ректорат_26092022
Ректорат_26092022 Стиль в искусстве – это мироощущение времени.
Стиль в искусстве – это мироощущение времени. «Ах, как я зол!.. Как я зла, Как я зла!», или Превратности любви
«Ах, как я зол!.. Как я зла, Как я зла!», или Превратности любви Польза мёда
Польза мёда Анксиолитки. Тревожные расстройства
Анксиолитки. Тревожные расстройства Present Simple (Indefinite) and Present Continuous(Progressive)
Present Simple (Indefinite) and Present Continuous(Progressive) Презентация на тему Демографическая ситуация в России
Презентация на тему Демографическая ситуация в России Severe flooding in Brazil and Australia
Severe flooding in Brazil and Australia  През. Введение в предмет информатики
През. Введение в предмет информатики 18.08 план развития музея Древние курганы Салбыкской степи (1)
18.08 план развития музея Древние курганы Салбыкской степи (1) Год культурного наследия народов России
Год культурного наследия народов России Создание электронного курса
Создание электронного курса ПЛАНИРОВАНИЕ СЕБЕСТОИМОСТИ, ПРИБЫЛИ И РЕНТАБЕЛЬНОСТИ
ПЛАНИРОВАНИЕ СЕБЕСТОИМОСТИ, ПРИБЫЛИ И РЕНТАБЕЛЬНОСТИ Скульптура и рельеф Древней Греции
Скульптура и рельеф Древней Греции Отдых в Крыму – Профилакторий “Солнечный”
Отдых в Крыму – Профилакторий “Солнечный” Т.Г. Шевченко-художник
Т.Г. Шевченко-художник Личностно-ориентированное воспитание младших школьников как фактор их социализации
Личностно-ориентированное воспитание младших школьников как фактор их социализации Элементы баскетбола на уроках в начальной школе
Элементы баскетбола на уроках в начальной школе Жанры цирка
Жанры цирка Презентация на тему Традиционный японский дом
Презентация на тему Традиционный японский дом  Подарочные наборы
Подарочные наборы