- Минеральное питание

Содержание
- 2. Поступление в клетку. Перенос через плазмалемму. Электрофизиологический подход Кинетический подход Молекулярно биологический подход
- 3. Электрофизиологический подход Потенциал Нернста может быть использован, чтобы установить тип транспорта: активный или пасивный? μ =
- 4. Потенциал Нернста – градиент электрического потециала на мембране, уравновешенный градиентом химического потенциала для данного иона по
- 5. Электрофизиологический подход позволяет предсказать тип транспорта Пассивный транспорт – движение иона через мембрану в сторону его
- 6. Сравнение наблюдаемых и предсказанных равновесных концентраций ионов в тканях корней гороха (мМ L-1), мембранный потенциал –110
- 7. Пассивный транспорт – движение иона через мембрану по градиенту его электрохимического потенциала. Активный транспорт - перемещение
- 8. Пассивный ионный транспорт по каналу. Ионные каналы – интегральные мембранные белки, образующие «пору» в мембране Транспорт
- 9. Наружный раствор Устье Цитозоль Ворота (gate) + + + + + + + + (-) (-)
- 10. Воротный механизм «работы» канальных белков Два дискретных состояния канального белка: канал открыт/канал закрыт
- 11. 4 типа белков К+ каналов 1999 кл. дрожжей (8) кл. животных и растений (2) кл. животных
- 12. Новые типы белков К+ каналов у растений июнь, 1999 Впервые о двупоровых К каналах у растений
- 13. S1 S2 S3 S4 S5 S6 H5(Р) N- конец C-конец А + + + + +
- 14. К+-каналы Шейкерного типа у растений: 5 семейств по гомологии аминокислотных последовательностей Филогенетическое древо К-каналов Шейкерного типа
- 15. Расположение К+ каналов разных типов у Arabidopsis SKOR AKT1 KAT1 AKT2/3 Sabine Zimmermann* and Herve Sentenac†
- 16. Ионные каналы в растительных клетках Катионные и анионные “Входящие” (in) и “выходящие” (out) Селективность: специфические и
- 17. Активный транспорт ΔрН АТФ РРнн Первичный активный транспорт Вторичный активный транспорт Способы (или механизмы) активного поглощения
- 18. Первичный активный транспорт
- 19. Н-помпы отвечают за создание и поддержание потенциала на мембране Н+-АТФазы плазмалеммы Н+-АТФаза тонопласта Пирофосфатаза тонопласта Емб
- 20. Н+-АТФаза плазмалеммы Р-типа Один большой полипептид 100 - 106 кДа Регулируется по принципу «фосфорилирование/дефосфорилирование» по серину
- 21. Структура белка Н-АТФазы плазмалеммы Один полипептид 100 – 106 кДа Домен связывания Mg-АТФ Е1 связывает Н
- 22. В процессе работы Н+-АТФаза Р-типа подвергается фосфорилированию/дефосфорилированию, меняя при этом свою активность неактивная активная суперактивная
- 23. Многомерная структура 70 кДа, 60 кДа, 16 кДа (7-10 субъединиц) Анионзависимая (нитрат ингибирует, хлорид стимулирует) Нечувствительна
- 24. Пирофосфатаза тонопласта 64 – 67 кДа катионзависимая (стимулируется К+, ингибируется Na+, Са2+) высокоспецифична к пирофосфату зависит
- 25. Метаболическая (электрогенная) и диффузионная компоненты Емб
- 27. Са2+ -АТФазы Са2+АТФазы принадлежат к большому кругу АТФаз Р-типа Са2+АТФазы ПМ Са2+АТФазы ЭР У растений в
- 28. ABC-транспортеры Транспортные белки, которые используют энергию гидролиза АТФ для транспорта через мембраны самых разных химических агентов
- 29. Plant Physiol, March 2003, Vol. 131, pp. 1169-1177 UPDATE ON ATP-BINDING CASSETTE TRANSPORTERS The ATP-Binding Cassette
- 30. 1.Вклад в поддержание ΔЕ на мембране. 2. Обеспечение движущей силы для пассивного и активного транспорта веществ.
- 31. ΔЕ = Δψ + ΔрН Размен Δψ на ΔрН Вторичный активный транспорт (симпорт) Ан- Н+ Форма
- 32. Вторичный активный транспорт
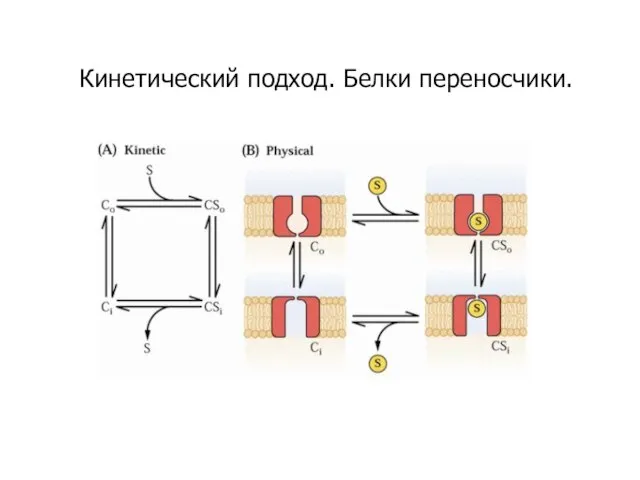
- 34. Кинетический подход. Белки переносчики.
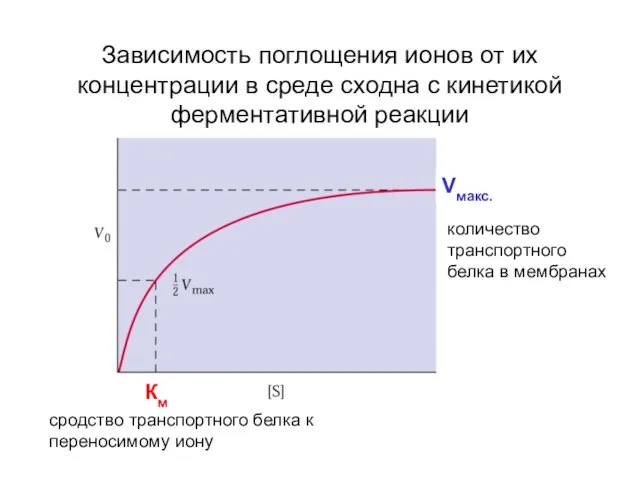
- 35. Зависимость поглощения ионов от их концентрации в среде сходна с кинетикой ферментативной реакции сродство транспортного белка
- 36. Кинетика поглощения ионов интактным растением 5 10 15 20 [Р] в растворе, мкМ⋅л-1 0 20 30
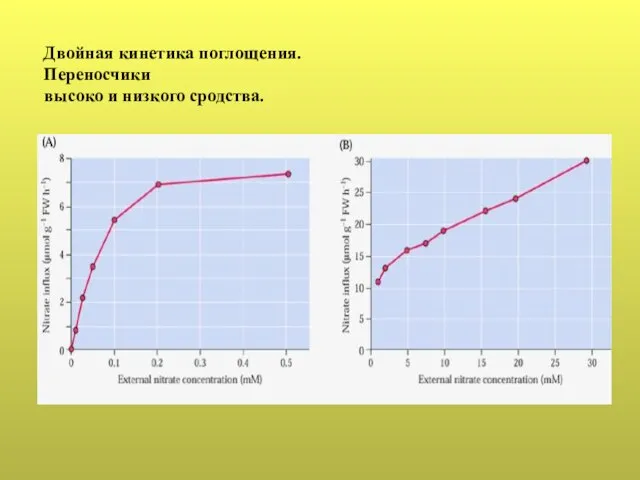
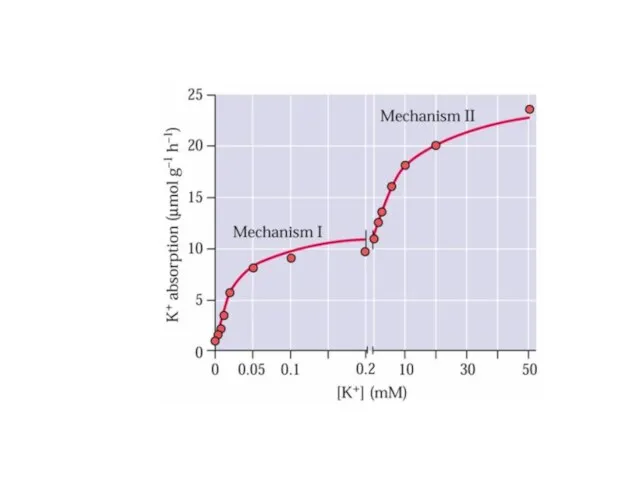
- 37. Двойная кинетика поглощения. Переносчики высоко и низкого сродства.
- 39. Сульфатные транспортеры высокого сродства Sultr1 или ST1 (Km порядка 10 мкмолей) низкого сродства Sultr2 или ST2
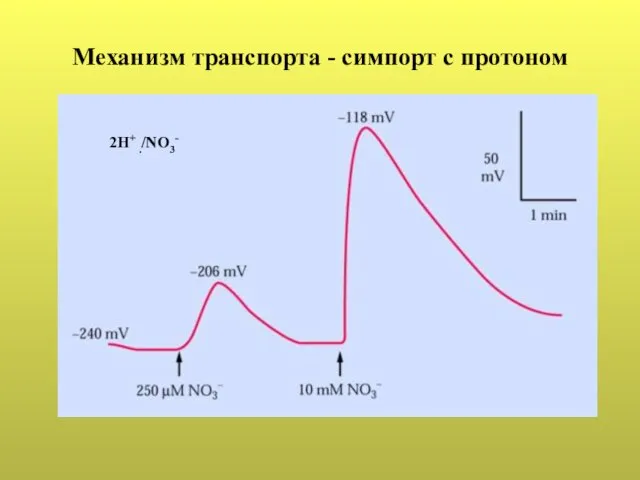
- 40. Механизм транспорта - симпорт с протоном 2Н+ ./NО3-
- 41. Вторичный активный транспорт (Сульфатный транспортер)
- 42. Переносчики, пермеазы, портеры, транспортеры Главное суперсемейство транспортных белков MFS (Major Facilitating Superfamily) – 12 трансмембранных доменов:
- 43. Микроэлементы: Mo, Zn, B, Cu, Mn Структура белка транспортера микроэлементов. ZIP семейство MFS (Major Facilitating Superfamily):
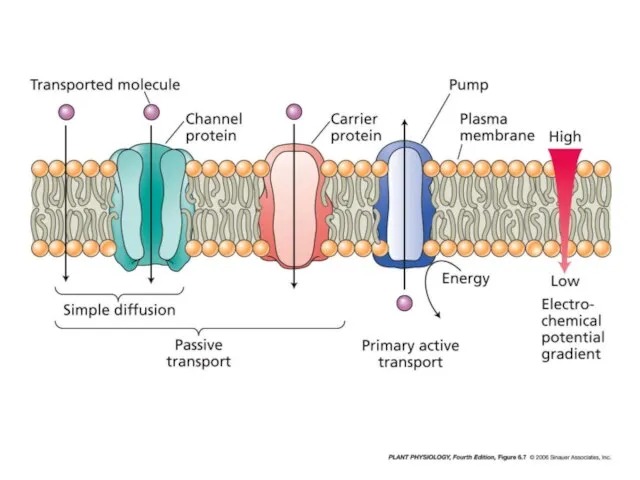
- 44. Активное поглощение и пассивное поглощение. Способы (или механизмы) активного поглощения различаются по формам потребляемой энергии.
- 46. Скачать презентацию
Слайд 2Поступление в клетку.
Перенос через плазмалемму.
Электрофизиологический подход
Кинетический подход
Молекулярно биологический подход
Поступление в клетку.
Перенос через плазмалемму.
Электрофизиологический подход
Кинетический подход
Молекулярно биологический подход

Слайд 3Электрофизиологический подход
Потенциал Нернста может быть использован, чтобы установить тип транспорта: активный
Электрофизиологический подход
Потенциал Нернста может быть использован, чтобы установить тип транспорта: активный

Слайд 4Потенциал Нернста – градиент электрического потециала на мембране, уравновешенный градиентом химического потенциала
Потенциал Нернста – градиент электрического потециала на мембране, уравновешенный градиентом химического потенциала
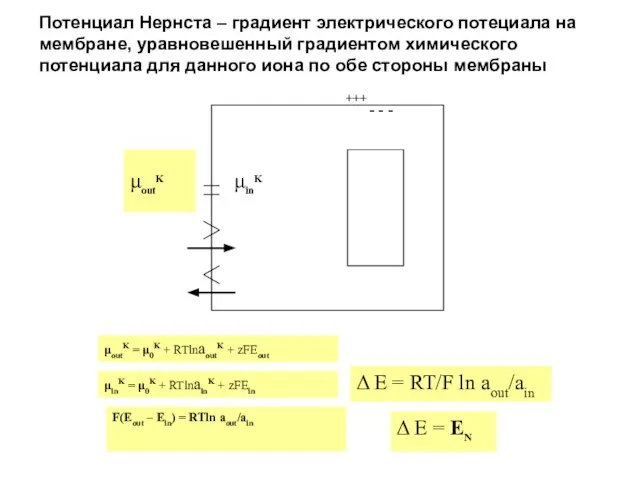
Слайд 5Электрофизиологический подход позволяет предсказать тип транспорта
Пассивный транспорт – движение иона через мембрану
Электрофизиологический подход позволяет предсказать тип транспорта
Пассивный транспорт – движение иона через мембрану

Слайд 6Сравнение наблюдаемых и предсказанных равновесных концентраций ионов в тканях корней гороха (мМ
Сравнение наблюдаемых и предсказанных равновесных концентраций ионов в тканях корней гороха (мМ
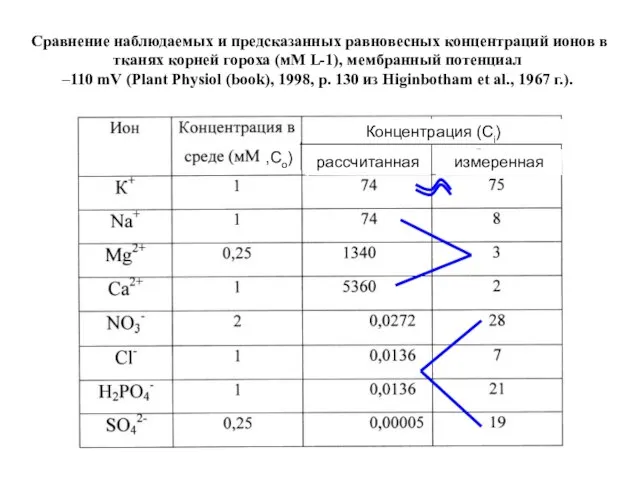
Слайд 7Пассивный транспорт – движение иона через мембрану по градиенту его электрохимического потенциала.
Активный
Пассивный транспорт – движение иона через мембрану по градиенту его электрохимического потенциала.
Активный

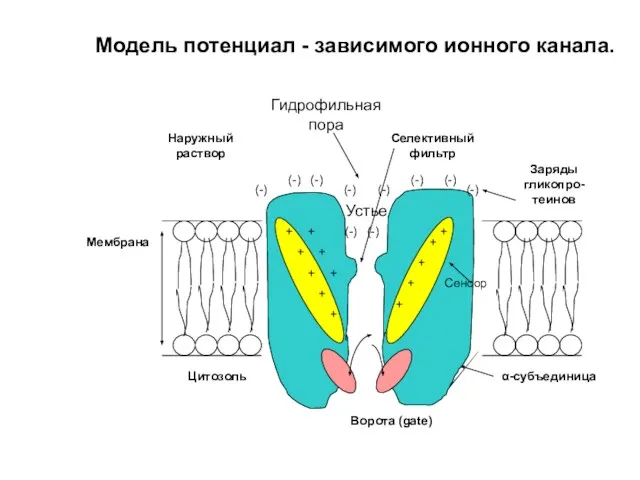
Слайд 8Пассивный ионный транспорт по каналу.
Ионные каналы – интегральные мембранные белки, образующие «пору»
Пассивный ионный транспорт по каналу.
Ионные каналы – интегральные мембранные белки, образующие «пору»

Слайд 9Наружный раствор
Устье
Цитозоль
Ворота (gate)
+
+
+
+
+
+
+
+
(-)
(-)
(-)
(-)
(-)
Заряды гликопро-теинов
(-)
Селективный фильтр
α-субъединица
+
+
+
+
+
(-)
(-)
(-)
(-)
Сенсор
Мембрана
Гидрофильная пора
Модель потенциал - зависимого ионного канала.
Наружный раствор
Устье
Цитозоль
Ворота (gate)
+
+
+
+
+
+
+
+
(-)
(-)
(-)
(-)
(-)
Заряды гликопро-теинов
(-)
Селективный фильтр
α-субъединица
+
+
+
+
+
(-)
(-)
(-)
(-)
Сенсор
Мембрана
Гидрофильная пора
Модель потенциал - зависимого ионного канала.
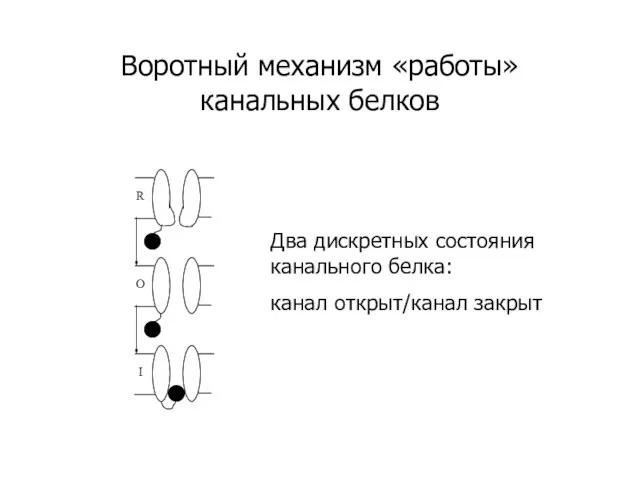
Слайд 10Воротный механизм «работы» канальных белков
Два дискретных состояния канального белка:
канал открыт/канал закрыт
Воротный механизм «работы» канальных белков
Два дискретных состояния канального белка:
канал открыт/канал закрыт
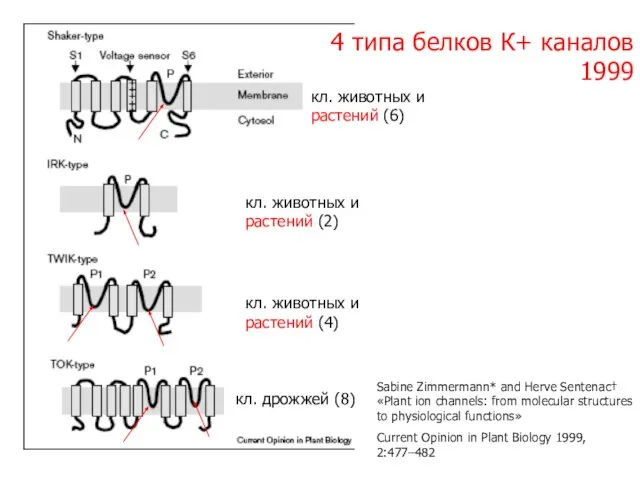
Слайд 114 типа белков К+ каналов
1999
кл. дрожжей (8)
кл. животных и растений (2)
кл.
4 типа белков К+ каналов
1999
кл. дрожжей (8)
кл. животных и растений (2)
кл.
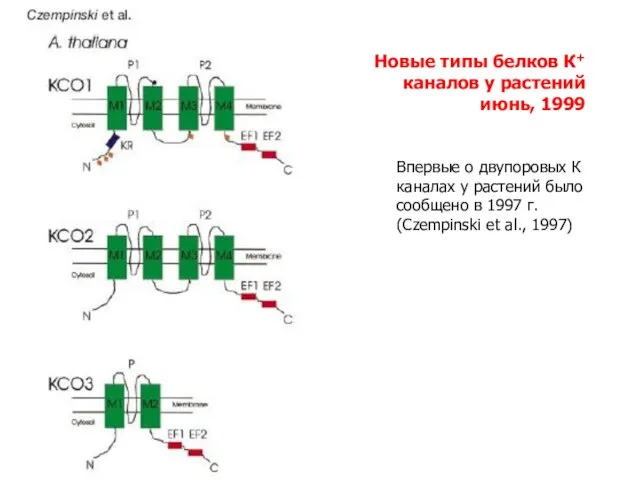
Слайд 12Новые типы белков К+ каналов у растений
июнь, 1999
Впервые о двупоровых К каналах
Новые типы белков К+ каналов у растений
июнь, 1999
Впервые о двупоровых К каналах
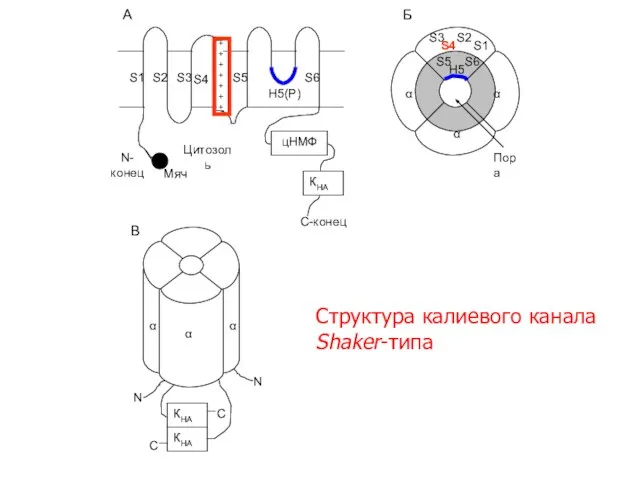
Слайд 13 S1
S2
S3
S4
S5
S6
H5(Р)
N-
конец
C-конец
А
+
+
+
+
S1
S2
S3
S4
S5
S6
H5(Р)
N-
конец
C-конец
А
+
+
+
+
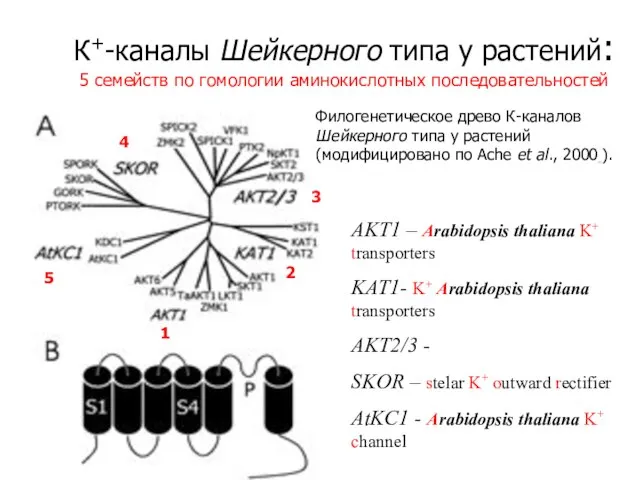
Слайд 14К+-каналы Шейкерного типа у растений:
5 семейств по гомологии аминокислотных последовательностей
Филогенетическое древо К-каналов
К+-каналы Шейкерного типа у растений:
5 семейств по гомологии аминокислотных последовательностей
Филогенетическое древо К-каналов
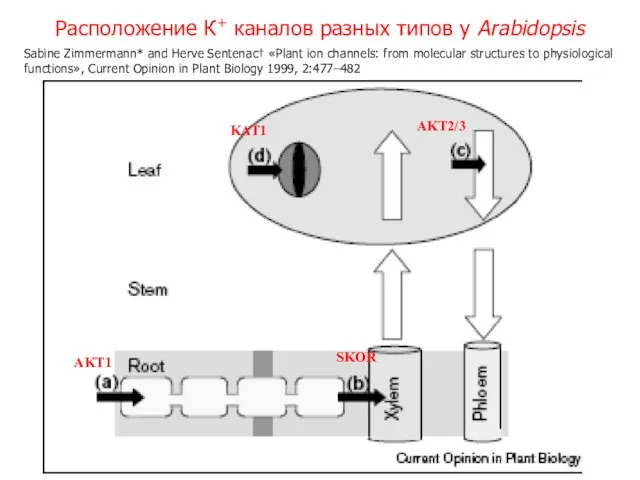
Слайд 15Расположение К+ каналов разных типов у Arabidopsis
SKOR
AKT1
KAT1
AKT2/3
Sabine Zimmermann* and Herve Sentenac†
Расположение К+ каналов разных типов у Arabidopsis
SKOR
AKT1
KAT1
AKT2/3
Sabine Zimmermann* and Herve Sentenac†
Слайд 16Ионные каналы в растительных клетках
Катионные и анионные
“Входящие” (in) и “выходящие” (out)
Селективность: специфические
Ионные каналы в растительных клетках
Катионные и анионные
“Входящие” (in) и “выходящие” (out)
Селективность: специфические

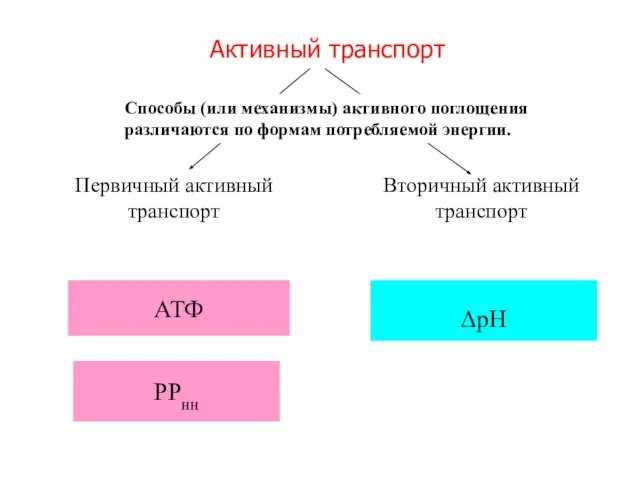
Слайд 17Активный транспорт
ΔрН
АТФ
РРнн
Первичный активный транспорт
Вторичный активный транспорт
Способы (или механизмы) активного поглощения различаются по
Активный транспорт
ΔрН
АТФ
РРнн
Первичный активный транспорт
Вторичный активный транспорт
Способы (или механизмы) активного поглощения различаются по
Слайд 18Первичный активный транспорт
Первичный активный транспорт

Слайд 19Н-помпы
отвечают за создание и поддержание потенциала на мембране
Н+-АТФазы плазмалеммы
Н+-АТФаза тонопласта
Пирофосфатаза тонопласта
Емб
Н-помпы
отвечают за создание и поддержание потенциала на мембране
Н+-АТФазы плазмалеммы
Н+-АТФаза тонопласта
Пирофосфатаза тонопласта
Емб

Слайд 20Н+-АТФаза плазмалеммы Р-типа
Один большой полипептид 100 - 106 кДа
Регулируется по принципу «фосфорилирование/дефосфорилирование»
Н+-АТФаза плазмалеммы Р-типа
Один большой полипептид 100 - 106 кДа
Регулируется по принципу «фосфорилирование/дефосфорилирование»

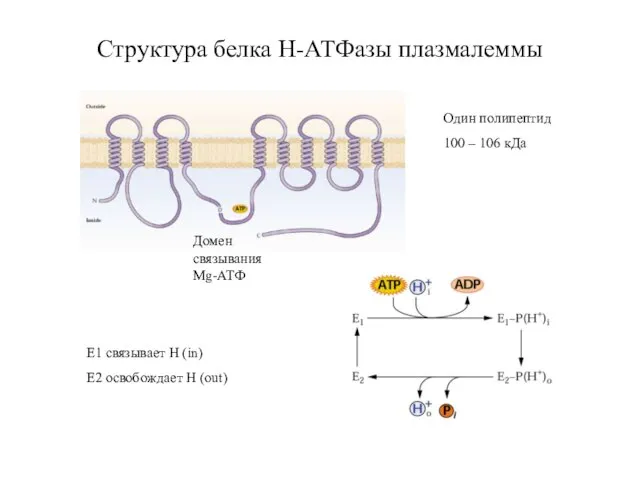
Слайд 21Структура белка Н-АТФазы плазмалеммы
Один полипептид
100 – 106 кДа
Домен связывания Mg-АТФ
Е1 связывает Н
Структура белка Н-АТФазы плазмалеммы
Один полипептид
100 – 106 кДа
Домен связывания Mg-АТФ
Е1 связывает Н
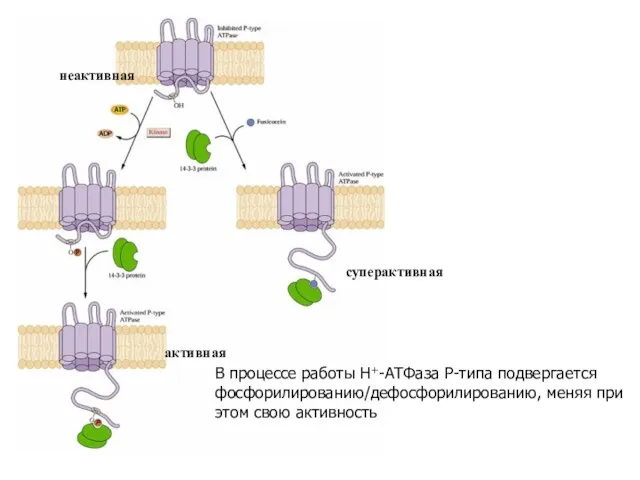
Слайд 22В процессе работы Н+-АТФаза Р-типа подвергается фосфорилированию/дефосфорилированию, меняя при этом свою активность
неактивная
активная
суперактивная
В процессе работы Н+-АТФаза Р-типа подвергается фосфорилированию/дефосфорилированию, меняя при этом свою активность
неактивная
активная
суперактивная
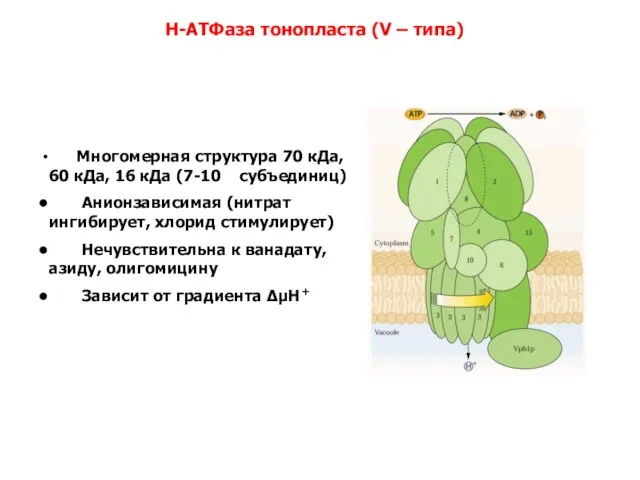
Слайд 23 Многомерная структура 70 кДа, 60 кДа, 16 кДа (7-10 субъединиц)
Анионзависимая
Многомерная структура 70 кДа, 60 кДа, 16 кДа (7-10 субъединиц)
Анионзависимая
Слайд 24Пирофосфатаза тонопласта
64 – 67 кДа
катионзависимая (стимулируется К+, ингибируется Na+, Са2+)
Пирофосфатаза тонопласта
64 – 67 кДа
катионзависимая (стимулируется К+, ингибируется Na+, Са2+)

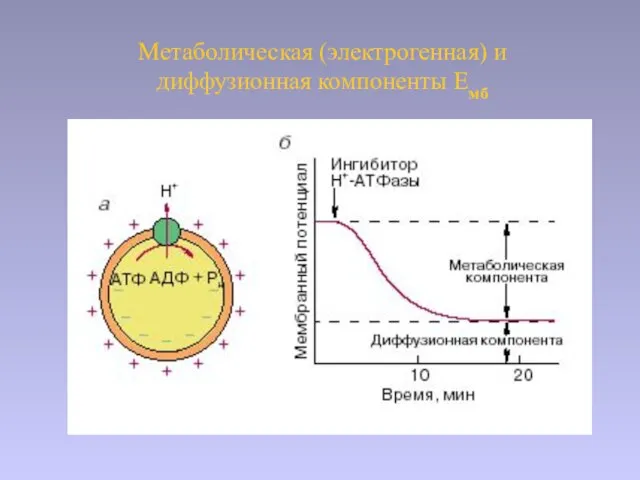
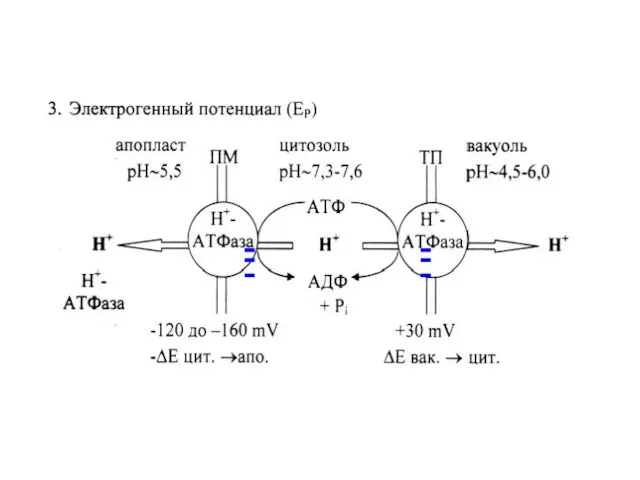
Слайд 25Метаболическая (электрогенная) и диффузионная компоненты Емб
Метаболическая (электрогенная) и диффузионная компоненты Емб
Слайд 27Са2+ -АТФазы
Са2+АТФазы принадлежат к большому кругу АТФаз Р-типа
Са2+АТФазы ПМ
Са2+АТФазы ЭР
У
Са2+ -АТФазы
Са2+АТФазы принадлежат к большому кругу АТФаз Р-типа
Са2+АТФазы ПМ
Са2+АТФазы ЭР
У

Слайд 28ABC-транспортеры
Транспортные белки, которые используют энергию гидролиза АТФ для транспорта через мембраны самых
ABC-транспортеры
Транспортные белки, которые используют энергию гидролиза АТФ для транспорта через мембраны самых

Слайд 29Plant Physiol, March 2003, Vol. 131, pp. 1169-1177
UPDATE ON ATP-BINDING CASSETTE
Plant Physiol, March 2003, Vol. 131, pp. 1169-1177
UPDATE ON ATP-BINDING CASSETTE
Слайд 301.Вклад в поддержание ΔЕ на мембране.
2. Обеспечение движущей силы для пассивного и
1.Вклад в поддержание ΔЕ на мембране.
2. Обеспечение движущей силы для пассивного и

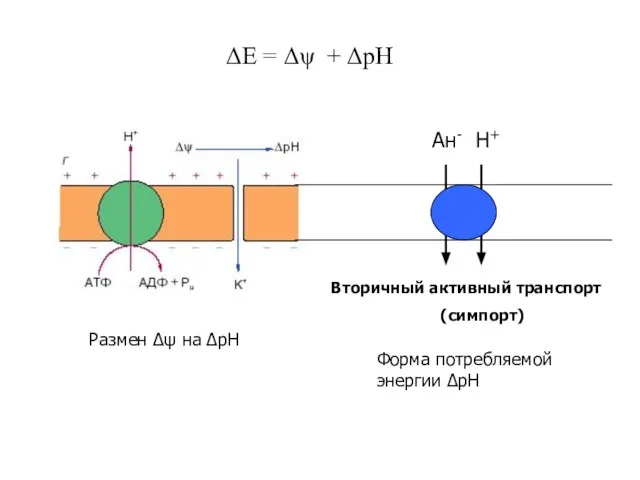
Слайд 31ΔЕ = Δψ + ΔрН
Размен Δψ на ΔрН
Вторичный активный транспорт
(симпорт)
Ан- Н+
Форма потребляемой
ΔЕ = Δψ + ΔрН
Размен Δψ на ΔрН
Вторичный активный транспорт
(симпорт)
Ан- Н+
Форма потребляемой
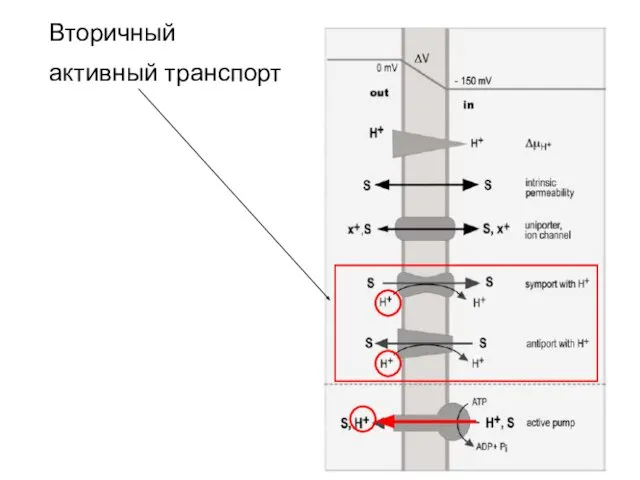
Слайд 32Вторичный
активный транспорт
Вторичный
активный транспорт
Слайд 34Кинетический подход. Белки переносчики.
Кинетический подход. Белки переносчики.
Слайд 35Зависимость поглощения ионов от их концентрации в среде сходна с кинетикой ферментативной
Зависимость поглощения ионов от их концентрации в среде сходна с кинетикой ферментативной
Слайд 36Кинетика поглощения ионов интактным растением
5
10
15
20
[Р] в растворе,
мкМ⋅л-1
0
20
30
10
40
Imax
½ Imax
Cmin
Kmкаж
Еn
Поступление
Кинетика поглощения ионов интактным растением
5
10
15
20
[Р] в растворе,
мкМ⋅л-1
0
20
30
10
40
Imax
½ Imax
Cmin
Kmкаж
Еn
Поступление
![Кинетика поглощения ионов интактным растением 5 10 15 20 [Р] в растворе,](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/377469/slide-35.jpg)
Слайд 37Двойная кинетика поглощения.
Переносчики
высоко и низкого сродства.
Двойная кинетика поглощения.
Переносчики
высоко и низкого сродства.
Слайд 39Сульфатные транспортеры
высокого сродства Sultr1 или ST1 (Km порядка 10 мкмолей)
низкого сродства
Сульфатные транспортеры
высокого сродства Sultr1 или ST1 (Km порядка 10 мкмолей)
низкого сродства

Слайд 40Механизм транспорта - симпорт с протоном
2Н+ ./NО3-
Механизм транспорта - симпорт с протоном
2Н+ ./NО3-
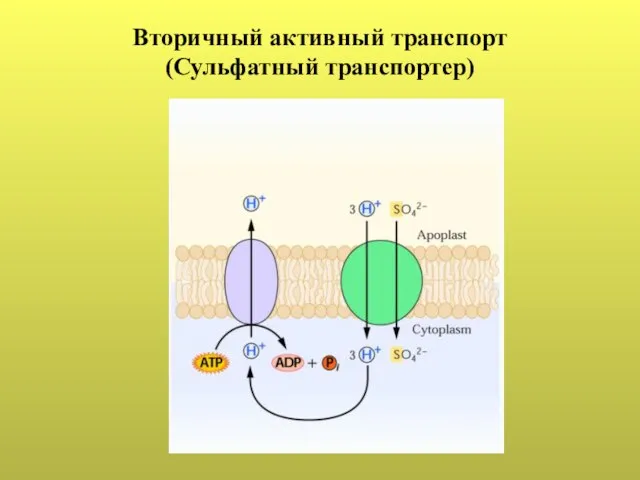
Слайд 41Вторичный активный транспорт
(Сульфатный транспортер)
Вторичный активный транспорт
(Сульфатный транспортер)
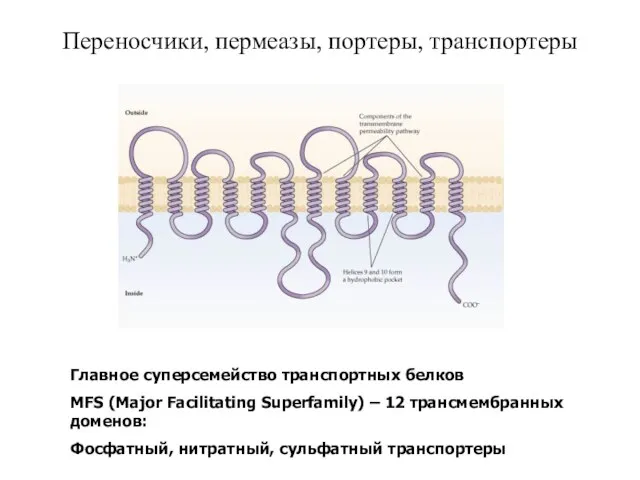
Слайд 42Переносчики, пермеазы, портеры, транспортеры
Главное суперсемейство транспортных белков
MFS (Major Facilitating Superfamily) –
Переносчики, пермеазы, портеры, транспортеры
Главное суперсемейство транспортных белков
MFS (Major Facilitating Superfamily) –
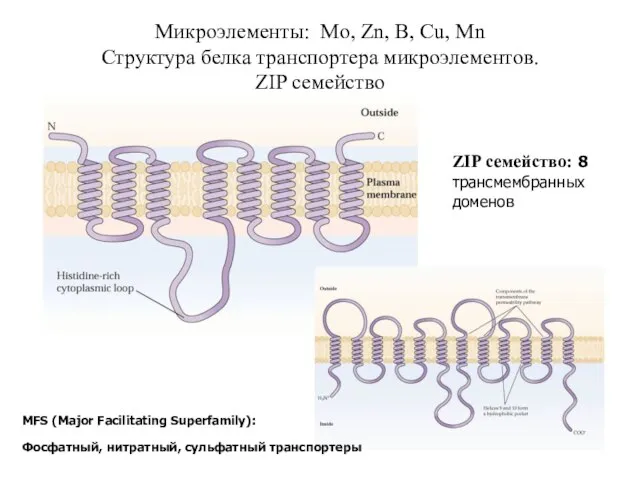
Слайд 43Микроэлементы: Mo, Zn, B, Cu, Mn
Структура белка транспортера микроэлементов.
ZIP семейство
MFS (Major Facilitating
Микроэлементы: Mo, Zn, B, Cu, Mn
Структура белка транспортера микроэлементов.
ZIP семейство
MFS (Major Facilitating
Слайд 44Активное поглощение и пассивное поглощение.
Способы (или механизмы) активного поглощения различаются по
Активное поглощение и пассивное поглощение.
Способы (или механизмы) активного поглощения различаются по

 Презентация на тему КРЫЛАТЫЕ СЛОВА
Презентация на тему КРЫЛАТЫЕ СЛОВА  Правописание приставок 6 класс
Правописание приставок 6 класс Аудитория украинского интернета: статистика и тенденции
Аудитория украинского интернета: статистика и тенденции Нормативно-правовое обеспечение электронной коммерции
Нормативно-правовое обеспечение электронной коммерции Презентация на тему Механическая работа и мощность (7 класс)
Презентация на тему Механическая работа и мощность (7 класс) Астероиды - космические лилипуты
Астероиды - космические лилипуты Питание и органы пищеварения
Питание и органы пищеварения Проект «Организация сбора макулатуры в районе Северное Измайлово»
Проект «Организация сбора макулатуры в районе Северное Измайлово» Технология продаж. Уровень 1
Технология продаж. Уровень 1 Тема: Моделирование физических явлений при помощи пакета Mathematica
Тема: Моделирование физических явлений при помощи пакета Mathematica Моллюски. Обитатели морского дна
Моллюски. Обитатели морского дна Новые возможности увеличения дохода РА и повышения лояльности клиентов за счёт использования партнёрских программ
Новые возможности увеличения дохода РА и повышения лояльности клиентов за счёт использования партнёрских программ экономика батецкий
экономика батецкий Презентация на тему Что такое плотность вещества (исследовательская работа)?
Презентация на тему Что такое плотность вещества (исследовательская работа)?  Школа личностного роста детей – инвалидов в условиях общественного благотворительного учреждения « Центр – Благодать»
Школа личностного роста детей – инвалидов в условиях общественного благотворительного учреждения « Центр – Благодать» Шаблон проекта
Шаблон проекта Giving presentations
Giving presentations Презентация на тему Мы - россияне
Презентация на тему Мы - россияне  Мозырский райисполком проводит открытый аукцион на право заключения договора аренды земельного участка
Мозырский райисполком проводит открытый аукцион на право заключения договора аренды земельного участка Презентация на тему Природный и антропогенный ландшафт
Презентация на тему Природный и антропогенный ландшафт Открытия российского кинематографа
Открытия российского кинематографа Презентация проекта
Презентация проекта ПРЕЗЕНТАЦИЯ рабочих мест в учреждениях образования Боровичского муниципального района
ПРЕЗЕНТАЦИЯ рабочих мест в учреждениях образования Боровичского муниципального района Планеты в домах гороскопа
Планеты в домах гороскопа Совершенствование системы ЛО
Совершенствование системы ЛО Детская агрессия
Детская агрессия Решение заданий ЕГЭ математика В6
Решение заданий ЕГЭ математика В6 Клочкова Нина Федоровна
Клочкова Нина Федоровна