- МОБИЛЬНЫЕ ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ

Содержание
- 2. В геномах плазмид, бактерий и эукариот широко распространены особые генетические элементы, способные перемещаться из одного участка
- 3. В большинстве своем мобильные элементы прокариот и эукариот построены по сходному плану. Сами элементы состоят из
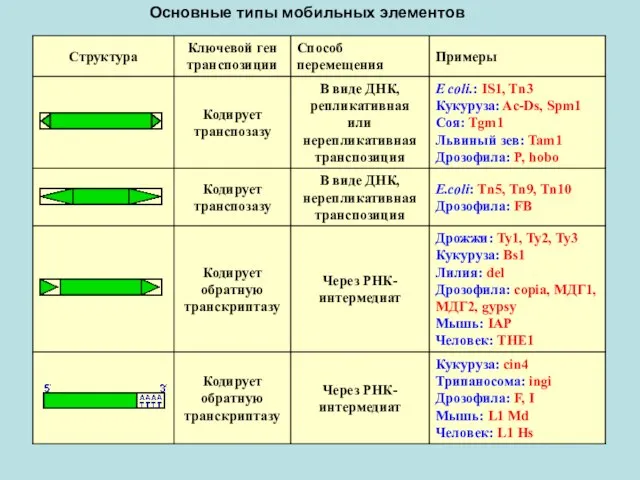
- 4. Основные типы мобильных элементов
- 5. ИП абсолютно необходимы для транспозиции, поскольку именно их концы связываются транспозазой, и по ним происходит рекомбинация.
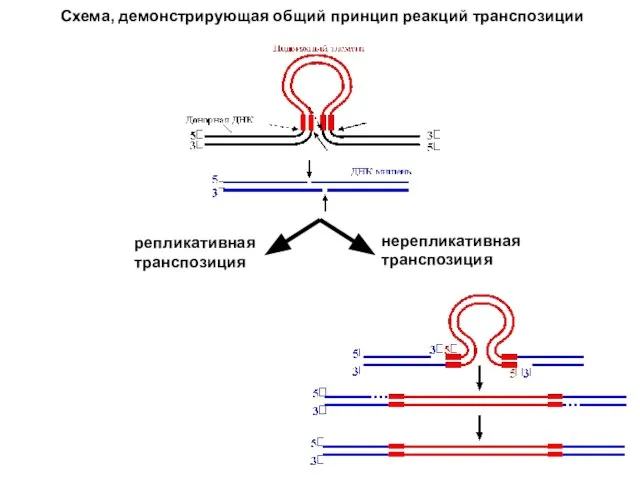
- 6. Структура мобильных элементов определяет механизмы их перемещений. Хотя эти механизмы различаются в деталях, имеется общий принцип
- 7. репликативная транспозиция нерепликативная транспозиция Схема, демонстрирующая общий принцип реакций транспозиции
- 8. МОБИЛЬНЫЕ ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ ПРОКАРИОТ: IS-элементы, транспозоны Для бактерий и плазмид характерны мобильные элементы с короткими или
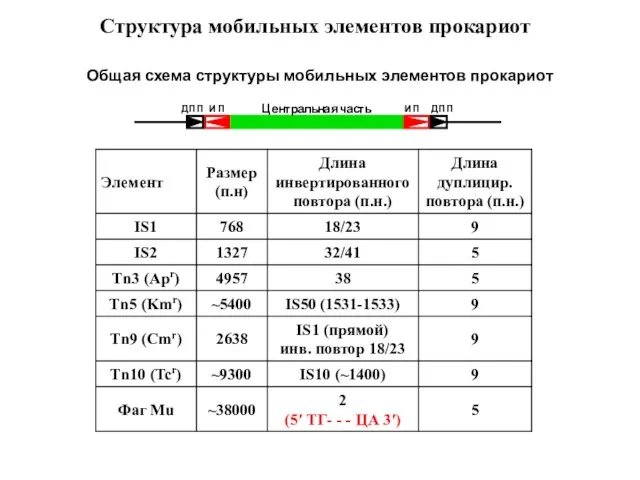
- 9. Структура мобильных элементов прокариот Общая схема структуры мобильных элементов прокариот
- 10. Теперь рассмотрим детали. Основные механизмы транспозиций изображены на рисунках, следующих ниже. Репликативная транспозиция отличается тем, что
- 11. Структура транспозона Tn3
- 13. Транспозон Tn3 представляет семейство мобильных элементов с короткими ИП (35-50 п.н.), перемещающимися с помощью репликативной транспозиции
- 14. Большинство прокариотических мобильных элементов перемещается с помощью нерепликативной транспозиции. Нерепликативная транспозиция заключается в вырезании элемента и
- 15. МОБИЛЬНЫЕ ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ ЭУКАРИОТ Мобильные элементы эукариот значительно разнообразнее прокариотических элементов. У эукариот распространены разнообразные мобильные
- 16. Примерами мобильных элементов класса II.1 у дрозофилы являются элементы Р и hobo. Р-элемент содержится в количестве
- 17. К этому же типу транспозонов относятся многие мобильные элементы растений: элементы Spm кукурузы, Tgm1 сои, Tam1
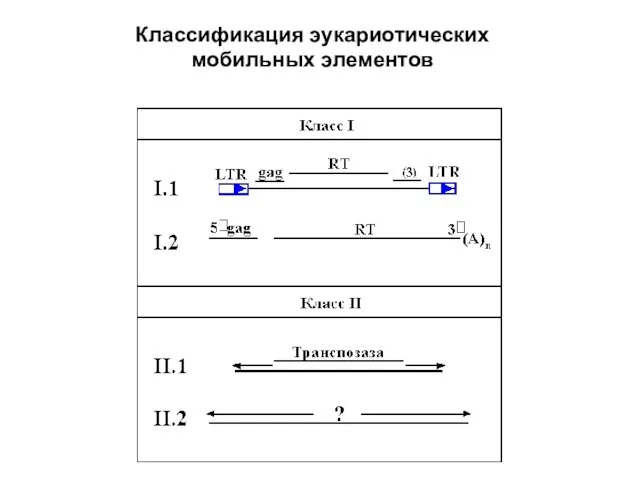
- 18. Классификация эукариотических мобильных элементов
- 20. У эукариот широко распространены ретротранспозоны, в транспозициях которых задействованы фермент обратная транскриптаза (ревертаза) и РНК-копия элемента
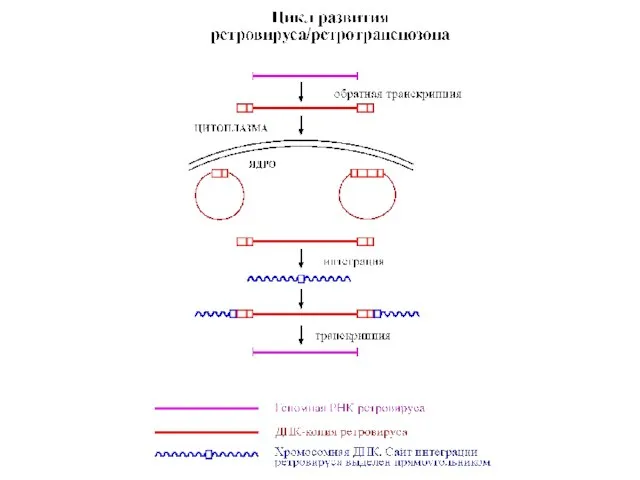
- 21. Ретровирусы являются «прототипами» ретротранспозонов. Их цикл развития состоит из чередования РНК- и ДНК-стадий. Вирионный геном представлен
- 22. В последние годы А. И. Ким и др. открыли, что мобильный элемент МДГ-4 (gypsy), содержит ген
- 23. У ретроэлементов с ДКП транспозиция происходит по схеме, включающей РНК-интермедиат. С геномной ДНК элемента транскрибируется РНК-копия,
- 25. Элементы без длинных концевых последовательностей: LINE и SINE Другая группа ретротранспозонов – элементы класса I.2 (ретропозоны).
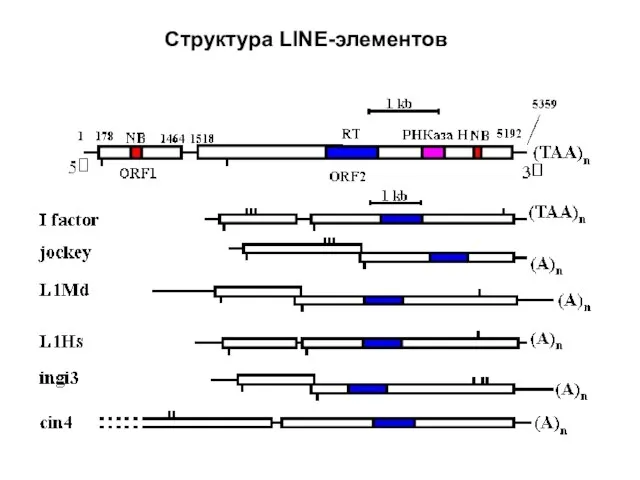
- 26. Структура LINE-элементов
- 27. LINE-элементы широко распространены как у беспозвоночных, так и у позвоночных. У млекопитающих LINE и SINE являются
- 28. Механизм перемещения LINE- и SINE-элементов представлен на рисунке. В отличие от ретротранспозонов I типа, здесь реакцию
- 29. Перемещение мобильного элемента LINE-типа
- 30. Мобильные ретроэлементы имеют большое биологическое значение. Как и все мобильные элементы, они вызывают хромосомные перестройки и
- 31. У представителей рода Drosophila, D.melanogaster и D.virilis теломеры, в отличие от других организмов, формируются путем последовательных
- 33. Скачать презентацию
Слайд 2В геномах плазмид, бактерий и эукариот широко распространены особые генетические элементы, способные
В геномах плазмид, бактерий и эукариот широко распространены особые генетические элементы, способные

Слайд 3В большинстве своем мобильные элементы прокариот и эукариот построены по сходному плану.
В большинстве своем мобильные элементы прокариот и эукариот построены по сходному плану.

Слайд 4
Основные типы мобильных элементов
Основные типы мобильных элементов
Слайд 5ИП абсолютно необходимы для транспозиции, поскольку именно их концы связываются транспозазой, и
ИП абсолютно необходимы для транспозиции, поскольку именно их концы связываются транспозазой, и

Слайд 6Структура мобильных элементов определяет механизмы их перемещений. Хотя эти механизмы различаются в
Структура мобильных элементов определяет механизмы их перемещений. Хотя эти механизмы различаются в

Слайд 7репликативная транспозиция
нерепликативная транспозиция
Схема, демонстрирующая общий принцип реакций транспозиции
репликативная транспозиция
нерепликативная транспозиция
Схема, демонстрирующая общий принцип реакций транспозиции
Слайд 8 МОБИЛЬНЫЕ ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ ПРОКАРИОТ:
IS-элементы, транспозоны
Для бактерий и плазмид характерны мобильные элементы
МОБИЛЬНЫЕ ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ ПРОКАРИОТ:
IS-элементы, транспозоны
Для бактерий и плазмид характерны мобильные элементы

Слайд 9Структура мобильных элементов прокариот
Общая схема структуры мобильных элементов прокариот
Структура мобильных элементов прокариот
Общая схема структуры мобильных элементов прокариот
Слайд 10Теперь рассмотрим детали. Основные механизмы транспозиций изображены на рисунках, следующих ниже.
Репликативная
Теперь рассмотрим детали. Основные механизмы транспозиций изображены на рисунках, следующих ниже.
Репликативная

Слайд 11Структура транспозона Tn3
Структура транспозона Tn3
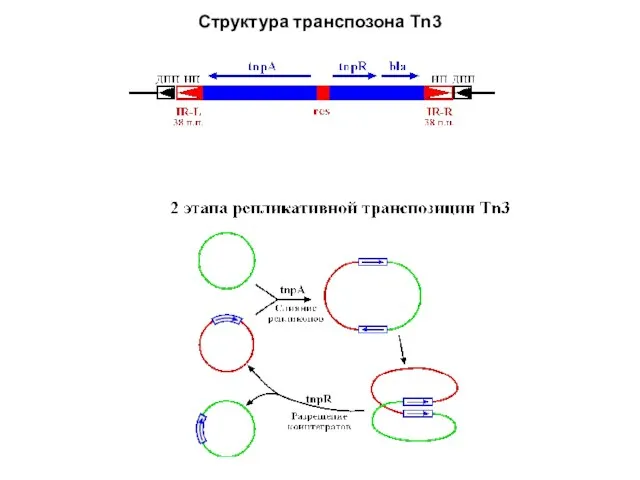
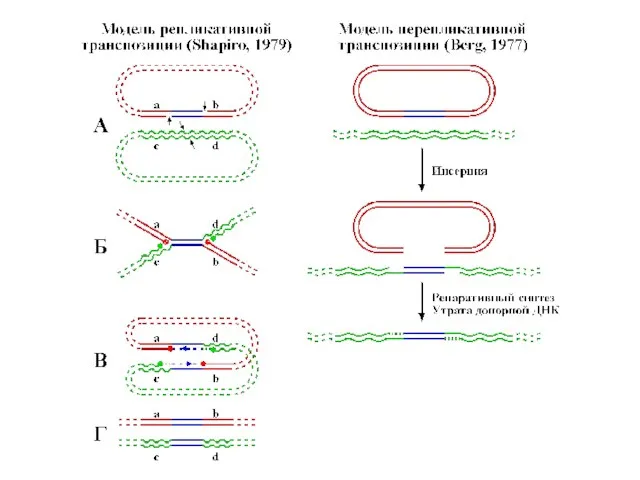
Слайд 13Транспозон Tn3 представляет семейство мобильных элементов с короткими ИП (35-50 п.н.), перемещающимися
Транспозон Tn3 представляет семейство мобильных элементов с короткими ИП (35-50 п.н.), перемещающимися

Слайд 14Большинство прокариотических мобильных элементов перемещается с помощью нерепликативной транспозиции.
Нерепликативная транспозиция заключается
Большинство прокариотических мобильных элементов перемещается с помощью нерепликативной транспозиции.
Нерепликативная транспозиция заключается

Слайд 15МОБИЛЬНЫЕ ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ ЭУКАРИОТ
Мобильные элементы эукариот значительно разнообразнее прокариотических элементов.
У эукариот распространены
МОБИЛЬНЫЕ ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ ЭУКАРИОТ
Мобильные элементы эукариот значительно разнообразнее прокариотических элементов.
У эукариот распространены

Слайд 16Примерами мобильных элементов класса II.1 у дрозофилы являются элементы Р и hobo.
Примерами мобильных элементов класса II.1 у дрозофилы являются элементы Р и hobo.

Слайд 17К этому же типу транспозонов относятся многие мобильные элементы растений: элементы Spm
К этому же типу транспозонов относятся многие мобильные элементы растений: элементы Spm

Слайд 18Классификация эукариотических
мобильных элементов
Классификация эукариотических
мобильных элементов

Слайд 20У эукариот широко распространены ретротранспозоны, в транспозициях которых задействованы фермент обратная транскриптаза
У эукариот широко распространены ретротранспозоны, в транспозициях которых задействованы фермент обратная транскриптаза

Слайд 21Ретровирусы являются «прототипами» ретротранспозонов. Их цикл развития состоит из чередования РНК- и

Слайд 22В последние годы А. И. Ким и др. открыли, что мобильный элемент МДГ-4 (gypsy),
В последние годы А. И. Ким и др. открыли, что мобильный элемент МДГ-4 (gypsy),

Слайд 23У ретроэлементов с ДКП транспозиция происходит по схеме, включающей РНК-интермедиат.
С геномной
У ретроэлементов с ДКП транспозиция происходит по схеме, включающей РНК-интермедиат.
С геномной

Слайд 25Элементы без длинных концевых последовательностей: LINE и SINE
Другая группа ретротранспозонов – элементы
Элементы без длинных концевых последовательностей: LINE и SINE
Другая группа ретротранспозонов – элементы

Слайд 26Структура LINE-элементов
Структура LINE-элементов
Слайд 27LINE-элементы широко распространены как у беспозвоночных, так и у позвоночных. У млекопитающих
LINE-элементы широко распространены как у беспозвоночных, так и у позвоночных. У млекопитающих

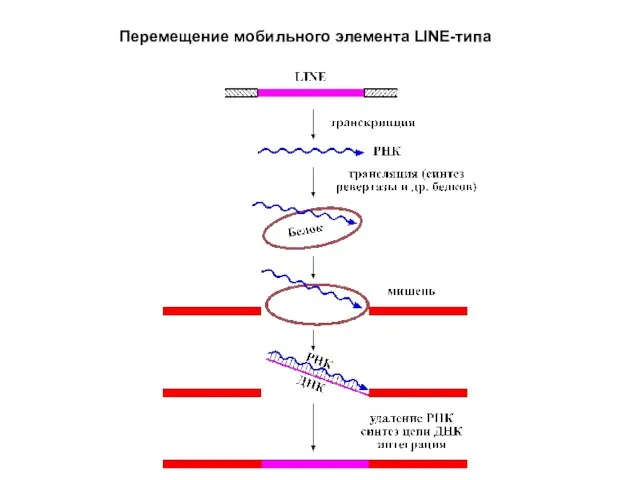
Слайд 28Механизм перемещения LINE- и SINE-элементов представлен на рисунке. В отличие от ретротранспозонов
Механизм перемещения LINE- и SINE-элементов представлен на рисунке. В отличие от ретротранспозонов

Слайд 29Перемещение мобильного элемента LINE-типа
Перемещение мобильного элемента LINE-типа
Слайд 30Мобильные ретроэлементы имеют большое биологическое значение.
Как и все мобильные элементы, они
Мобильные ретроэлементы имеют большое биологическое значение.
Как и все мобильные элементы, они

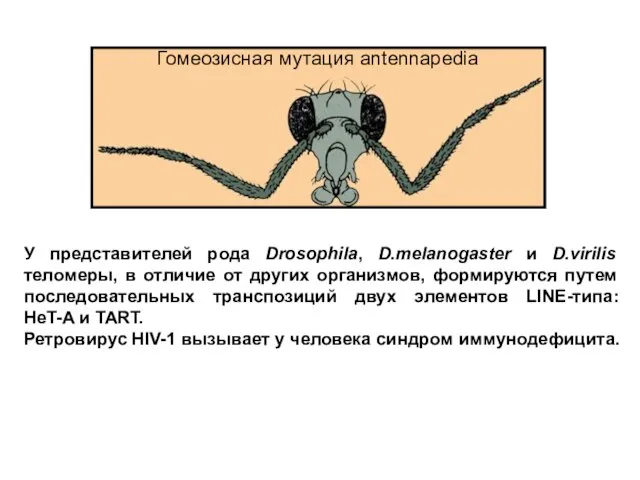
Слайд 31У представителей рода Drosophila, D.melanogaster и D.virilis теломеры, в отличие от других
У представителей рода Drosophila, D.melanogaster и D.virilis теломеры, в отличие от других
 Динамические методы оценки эффективности инвестиционных проектов (NPV, IRR)
Динамические методы оценки эффективности инвестиционных проектов (NPV, IRR) История арифметической и геометрической прогрессий
История арифметической и геометрической прогрессий Читать человека с первого взгляда. Чакры
Читать человека с первого взгляда. Чакры День Государственного флага Донецкой Народной Республики
День Государственного флага Донецкой Народной Республики Роль международного разделения труда в системе современной мировой экономики и международных экономических отношений
Роль международного разделения труда в системе современной мировой экономики и международных экономических отношений  Квадратные уравнения
Квадратные уравнения Каждый голос имеет значение Как надёжно подготовить сайт к нашествию посетителей
Каждый голос имеет значение Как надёжно подготовить сайт к нашествию посетителей modal_verbs_game_4_teacher_switcher
modal_verbs_game_4_teacher_switcher «Развитие детей в театрализованной деятельности» Музыкальный руководитель: Ахизова Елена Михайловна
«Развитие детей в театрализованной деятельности» Музыкальный руководитель: Ахизова Елена Михайловна Учебно-методический комплекс (УМК) по дисциплине «Инженерная и компьютерная графика»
Учебно-методический комплекс (УМК) по дисциплине «Инженерная и компьютерная графика» Презентация Выпускной
Презентация Выпускной ЧЕХОСЛОВАЦКИЕ ВОЕННЫЕ ФОРМИРОВАНИЯ В РОССИИ (1914-1920 ГГ.)Презентация магистерской работы Габрусевича С.А.
ЧЕХОСЛОВАЦКИЕ ВОЕННЫЕ ФОРМИРОВАНИЯ В РОССИИ (1914-1920 ГГ.)Презентация магистерской работы Габрусевича С.А. Анимализм в Англии XVIII в. Творчество Томаса Гейнсборо
Анимализм в Англии XVIII в. Творчество Томаса Гейнсборо Презентация на тему Жизнь и творчество Ломоносова
Презентация на тему Жизнь и творчество Ломоносова Датчик уровня наполненности
Датчик уровня наполненности ИНФОРМАЦИОНН ОЕОБЕСПЕЧЕНИЕ ИС
ИНФОРМАЦИОНН ОЕОБЕСПЕЧЕНИЕ ИС Изображение человека в движении. Эскиз модели
Изображение человека в движении. Эскиз модели Красота человека
Красота человека 984967
984967 История ЭВМ Автор: Николаева О. А.МОУ СОШ № 4 п. Хинганск
История ЭВМ Автор: Николаева О. А.МОУ СОШ № 4 п. Хинганск Первый раз, в первый класс!
Первый раз, в первый класс! Система мотивации продавцов
Система мотивации продавцов Инженер по эксплуатации железных дорог
Инженер по эксплуатации железных дорог Родник "Горный хрусталь"
Родник "Горный хрусталь" Для меня жизнь – Христос
Для меня жизнь – Христос функції мови
функції мови Презентация на тему Театр и музыка Античности
Презентация на тему Театр и музыка Античности Аттестационная работа. Проектная и исследовательская деятельность как способ формирования метапредметных результатов обучения
Аттестационная работа. Проектная и исследовательская деятельность как способ формирования метапредметных результатов обучения