- Механизмы репарации ДНК

Содержание
- 2. Механизмы репарации ДНК
- 3. Объекты для изучения репарации Escherichia coli Saccharomyces cerevisiae Caenorhabditis elegans Arabidopsis Thaliana Мыши, крысы Клеточные культуры
- 4. 1. 1. Мисматч репарация (MMR) Этапы 1 - 3
- 5. MMR. Этапы 4 - 5
- 6. А. Репарация мисматчей у бактерий VSP- very short patch repair Short patch repair Long patch repair
- 7. VSP- very short patch repair – 1 В основном удаляется Т из мисматчей G/T и C/T.
- 8. VSP- very short patch repair – 2 MutY заменяет А из мисматчей C/A и G/A. Это
- 9. MMR млекопитающих 9 генов: MLH1, MLH3, PMS1-2, MSH2-6 MSH – гомолог MutS MLH – гомолог MutL
- 10. MMR человека На примере болезни HNPCC (heredity non-polyposis colorectal cancer) в 1993-1994 гг. У человека найдено
- 11. MMR человека На примере болезни HNPCC (heredity non-polyposis colorectal cancer) в 1993-1994 гг. Пациенты с HNPCC
- 12. Комбинация генов при репарации мисматчей
- 13. Показатели риска заболевания раком (Standardized incidence ratios - SIRs) на основании популяционных и клинических исследований дефекта
- 14. Механизмы, осуществляющие вклад в специфичность клеточных типов, чувствительных к дефициту MMR
- 15. 1. 2. UVR репарация
- 16. SOS-мутагенез у бактерий
- 17. 2. Прямая репарация Репарируются О6-метилгуанин и О4-метилгуанин ферментом МТаза (MGMT). У Е. coli 2 фермента (гены
- 18. Пример реакции
- 19. 3. BER-репарация. Этапы 1-2
- 20. BER-репарация. Этапы 3-4
- 21. BER-репарация. Этап 5
- 23. 4. NER-репарация 1. TCR – transcription coupled repair (преимущественная репарация траснкрибируемых цепей гена) 2. GGR –
- 24. Больные пигментной ксеродермой ( Выявлена в 1968 г. Дефект одного из 7 или более XP генов
- 25. Больные TTD трихотиодистрофией (А) и CS кокаиновым синдромом (В)
- 26. Этапы NER. 1-3
- 27. Этапы NER. 4-6
- 28. Биохимия NER (Этапы 1-3)
- 29. Специфическая активность ХР нуклеаз
- 30. Биохимия NER (Этапы 4-5)
- 31. Повреждение ХР при болезнях ХР – мутации в генах XP A-D,F,G TTD – серо-дефицитные хрупкие волосы,
- 32. Вклад NER генов в развитие сквамозной карциномы головы и шеи
- 33. Роль pol lI в репарации Когда pol II взаимодействует с промотором, она находится в гипофосфорилированном статусе
- 34. Rad26 создает NER комплекс в сайте повреждения. В то же время Rsp5 (и Ubc5, не показано)
- 35. Аддукты ДНК с цис-платином
- 36. Репарация аддуктов ДНК с цис-платином
- 37. 5. Другие виды репарации ДНК
- 38. Альтернативный механизм репарации у дрожжей S. pombe
- 39. Основные этапы репарации
- 40. Белки ATM ATM (="ataxia telangiectasia mutated") получила название от болезни, пациенты, среди прочего, имеют высокий риск
- 41. Примеры ICL, вызванных антираковыми агентами
- 42. Клеточный ответ на ICLs
- 43. Репарация ICL у млекопитающих
- 44. Fanconi Anemia (FA)путь репарации У пациентов с FA повреждено, по крайней мере, 13 генов: FANCA, B,
- 45. Сравнение FA генов у человека, Drosophila, Dictyostelium and C. elegans
- 46. FA путь у C. elegans.
- 47. Fanconi Anemia путь регулирует репарацию ICLs ДНК с помощью гомологичной рекомбинации
- 49. SUMO (small ubiquitin-related modifier) конъюгация Несколько SUMO E3 лигаз выявлено: SP-RING (secretory protein with a RING
- 50. Моделирование влияния SUMO конъюгации на BRCA1 Генотоксический стресс запускает SUMO модификации BRCA1 через активность UBC9–PIAS1 и
- 51. IY. Эпигенетические модификации ДНК Модификации хроматина, Метилирование ДНК, Геномный импринтинг.
- 52. Нуклеосомная организация ДНК
- 53. Регуляции транскрипции ацетилированием гистонов Гистон-деацетилаза (HDACs) деацетилирует лизиновые остатки, создавая предпосылки для метилирования HMT. ДНК может
- 54. Метилирование ДНК
- 56. Распределение метилирования
- 57. Статус метилирования и функциональные особенности промоторов, содержащих CpG-островки
- 59. Функции ДНК-МТаз
- 60. Активация транскрипции метилированием ДНК
- 62. Метилирование ДНК и рак
- 63. Морфологические изменения в хроматине (a) Нормальный эпителий кишечника: ядра разделены, одинаковы по форме и размеру (мономорфны).
- 64. Общие эпигенетические изменения при раке
- 65. Метилирование ДНК и рак
- 66. Эпигенетическая модель рака кишечника
- 67. Примеры гиперметилирования некоторых генов
- 68. Примеры гиперметилирования некоторых генов
- 70. Скачать презентацию

Слайд 3Объекты для изучения репарации
Escherichia coli
Saccharomyces cerevisiae
Caenorhabditis elegans
Arabidopsis Thaliana
Мыши, крысы
Клеточные культуры человека
Объекты для изучения репарации
Escherichia coli
Saccharomyces cerevisiae
Caenorhabditis elegans
Arabidopsis Thaliana
Мыши, крысы
Клеточные культуры человека

Слайд 41. 1. Мисматч репарация (MMR) Этапы 1 - 3
1. 1. Мисматч репарация (MMR) Этапы 1 - 3
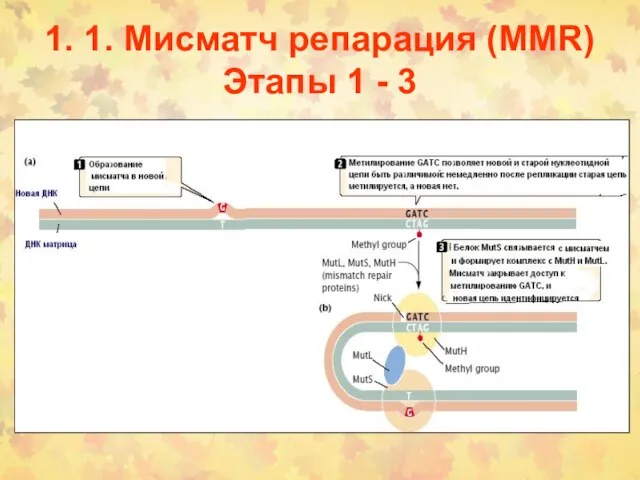
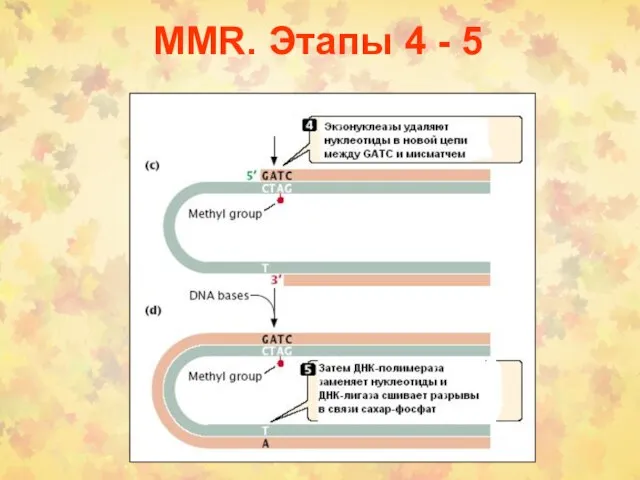
Слайд 5MMR. Этапы 4 - 5
MMR. Этапы 4 - 5
Слайд 6А. Репарация мисматчей
у бактерий
VSP- very short patch repair
Short patch repair
Long patch
А. Репарация мисматчей
у бактерий
VSP- very short patch repair
Short patch repair
Long patch

Слайд 7VSP- very short patch repair – 1
В основном удаляется Т из мисматчей
VSP- very short patch repair – 1
В основном удаляется Т из мисматчей

Слайд 8VSP- very short patch repair – 2
MutY заменяет А из мисматчей C/A
VSP- very short patch repair – 2
MutY заменяет А из мисматчей C/A

Слайд 9MMR млекопитающих
9 генов:
MLH1, MLH3, PMS1-2, MSH2-6
MSH – гомолог MutS
MLH – гомолог MutL
MSH2-6
MMR млекопитающих
9 генов:
MLH1, MLH3, PMS1-2, MSH2-6
MSH – гомолог MutS
MLH – гомолог MutL
MSH2-6

Слайд 10MMR человека
На примере болезни HNPCC (heredity
non-polyposis colorectal cancer) в 1993-1994 гг.
У
MMR человека
На примере болезни HNPCC (heredity
non-polyposis colorectal cancer) в 1993-1994 гг.
У

Слайд 11MMR человека
На примере болезни HNPCC (heredity non-polyposis colorectal cancer) в 1993-1994 гг.
Пациенты
MMR человека
На примере болезни HNPCC (heredity non-polyposis colorectal cancer) в 1993-1994 гг.
Пациенты

Слайд 12Комбинация генов при репарации мисматчей
Комбинация генов при репарации мисматчей
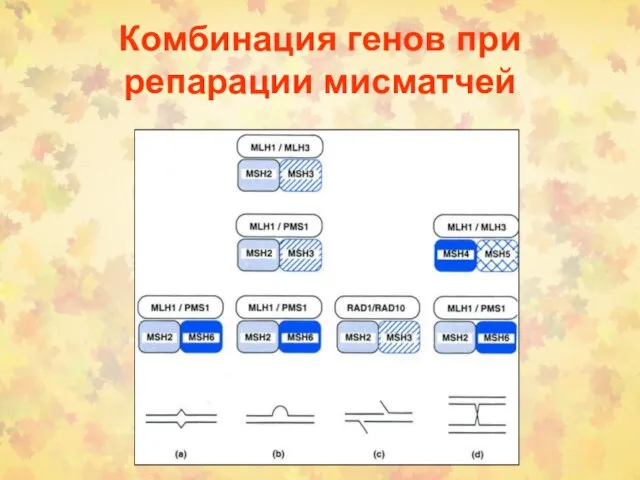
Слайд 13Показатели риска заболевания раком
(Standardized incidence ratios - SIRs) на основании популяционных
Показатели риска заболевания раком (Standardized incidence ratios - SIRs) на основании популяционных
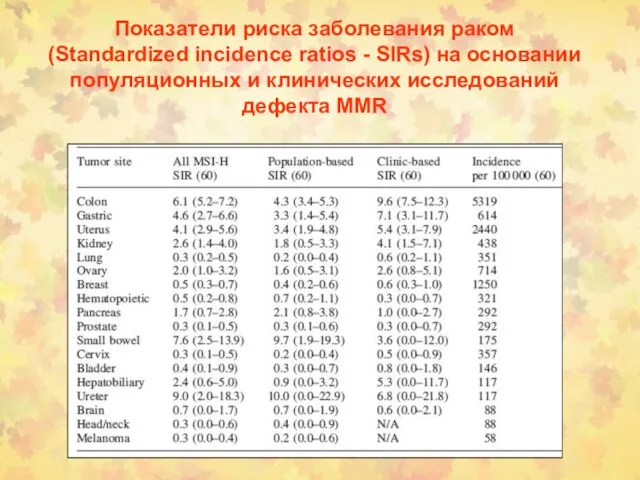
Слайд 14Механизмы, осуществляющие вклад в специфичность клеточных типов, чувствительных к дефициту MMR
Механизмы, осуществляющие вклад в специфичность клеточных типов, чувствительных к дефициту MMR

Слайд 151. 2. UVR репарация
1. 2. UVR репарация
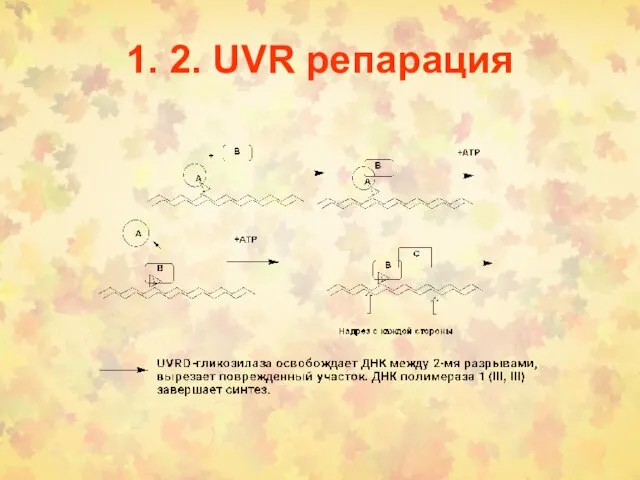
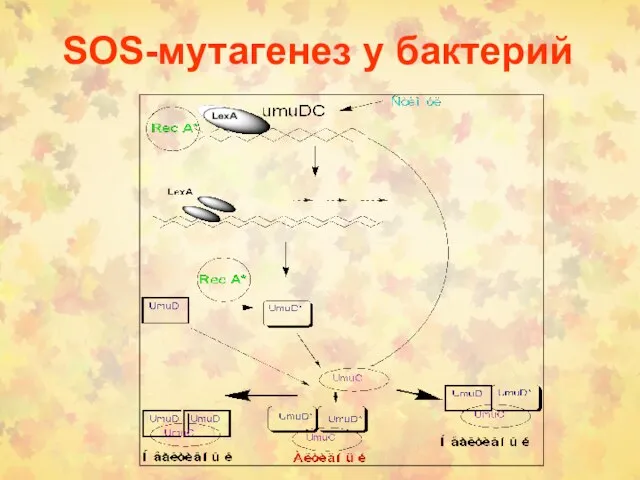
Слайд 16SOS-мутагенез у бактерий
SOS-мутагенез у бактерий
Слайд 172. Прямая репарация
Репарируются О6-метилгуанин и О4-метилгуанин ферментом МТаза (MGMT).
У Е. coli 2
2. Прямая репарация
Репарируются О6-метилгуанин и О4-метилгуанин ферментом МТаза (MGMT).
У Е. coli 2

Слайд 18Пример реакции
Пример реакции
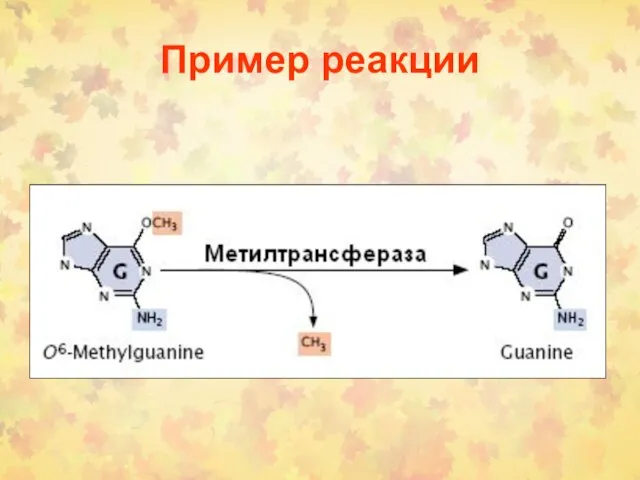
Слайд 193. BER-репарация. Этапы 1-2
3. BER-репарация. Этапы 1-2
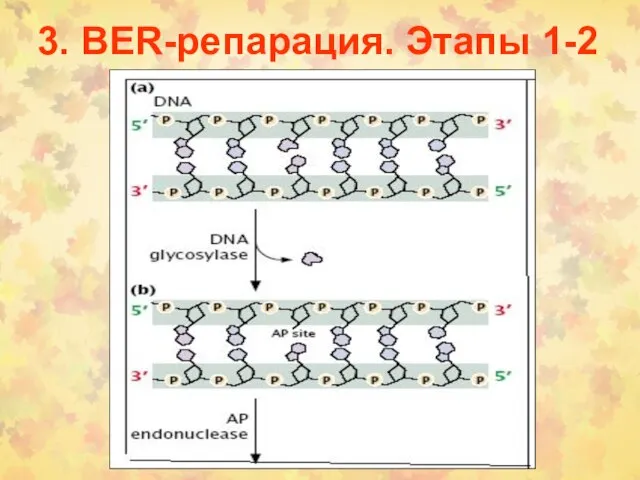
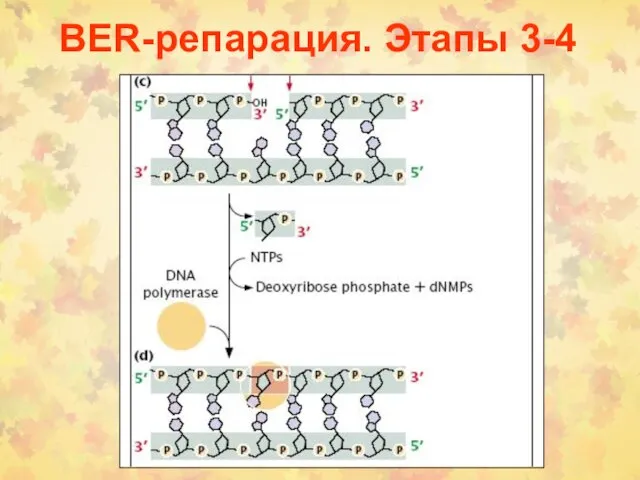
Слайд 20BER-репарация. Этапы 3-4
BER-репарация. Этапы 3-4
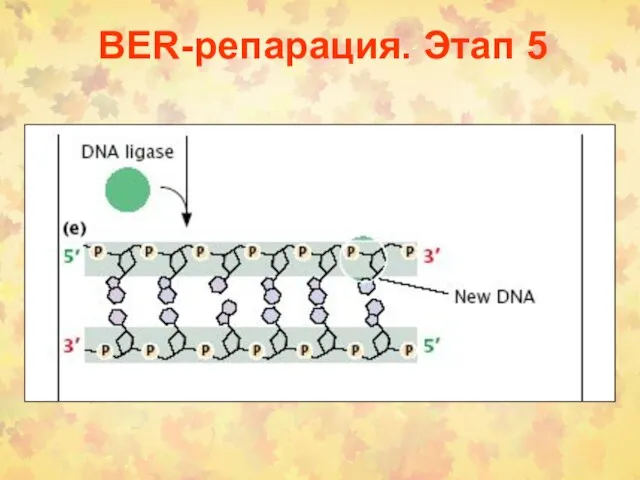
Слайд 21BER-репарация. Этап 5
BER-репарация. Этап 5
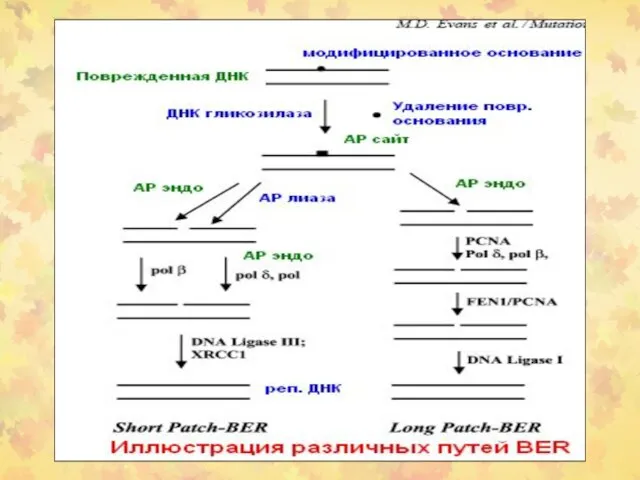
Слайд 234. NER-репарация
1. TCR – transcription coupled repair (преимущественная репарация траснкрибируемых цепей гена)
2.
4. NER-репарация
1. TCR – transcription coupled repair (преимущественная репарация траснкрибируемых цепей гена)
2.

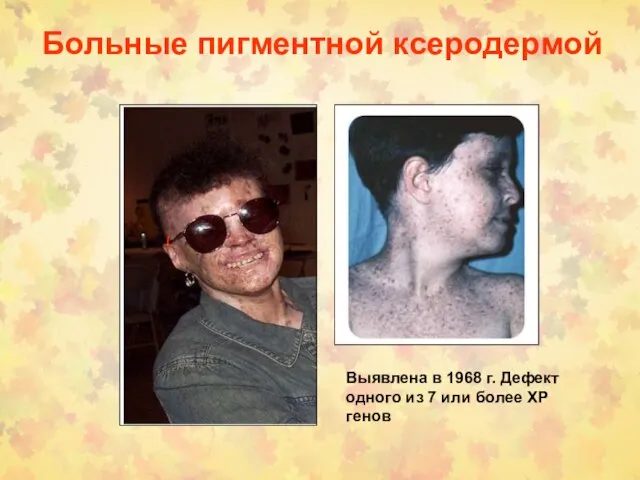
Слайд 24Больные пигментной ксеродермой
(
Выявлена в 1968 г. Дефект одного из 7 или более
Больные пигментной ксеродермой
(
Выявлена в 1968 г. Дефект одного из 7 или более
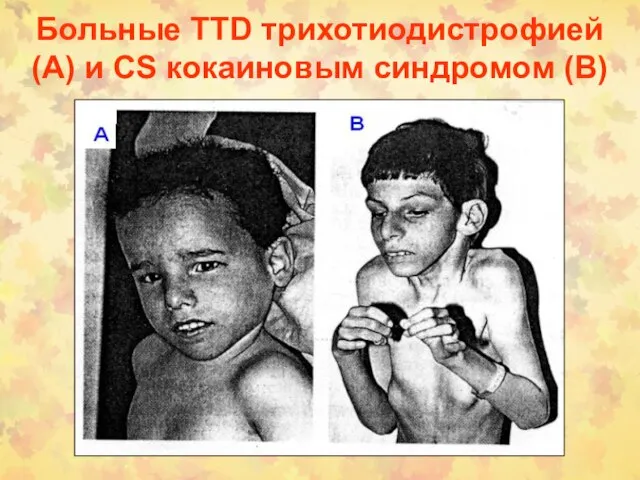
Слайд 25Больные TTD трихотиодистрофией (А) и CS кокаиновым синдромом (В)
Больные TTD трихотиодистрофией (А) и CS кокаиновым синдромом (В)
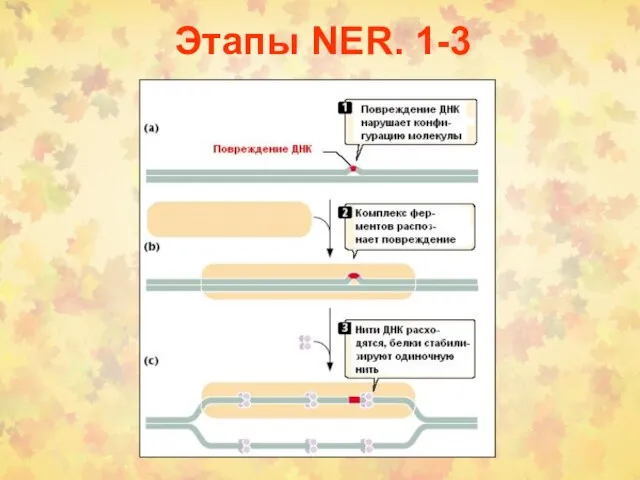
Слайд 26Этапы NER. 1-3
Этапы NER. 1-3
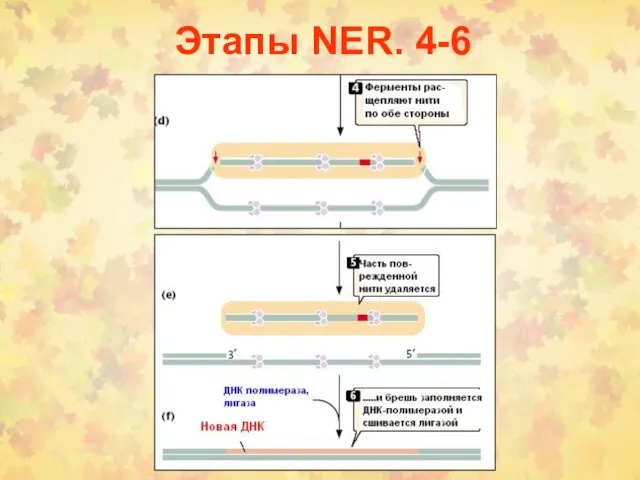
Слайд 27Этапы NER. 4-6
Этапы NER. 4-6
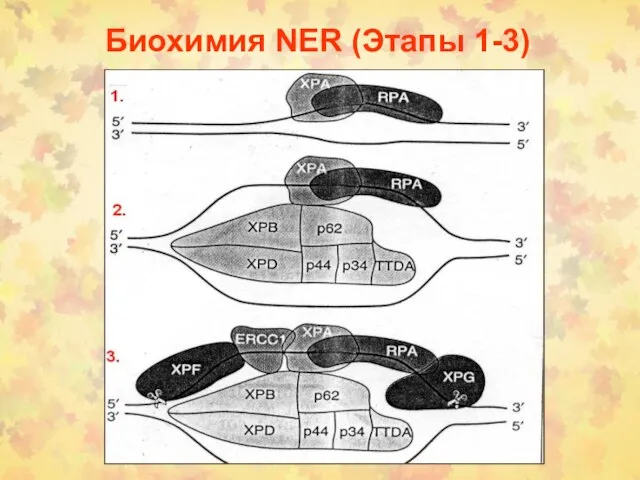
Слайд 28Биохимия NER (Этапы 1-3)
Биохимия NER (Этапы 1-3)
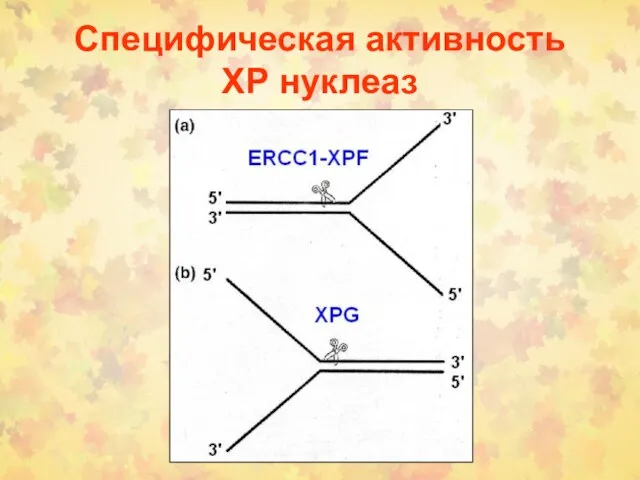
Слайд 29Специфическая активность
ХР нуклеаз
Специфическая активность
ХР нуклеаз
Слайд 30Биохимия NER (Этапы 4-5)
Биохимия NER (Этапы 4-5)

Слайд 31Повреждение ХР при болезнях
ХР – мутации в генах XP A-D,F,G
TTD – серо-дефицитные
Повреждение ХР при болезнях
ХР – мутации в генах XP A-D,F,G
TTD – серо-дефицитные

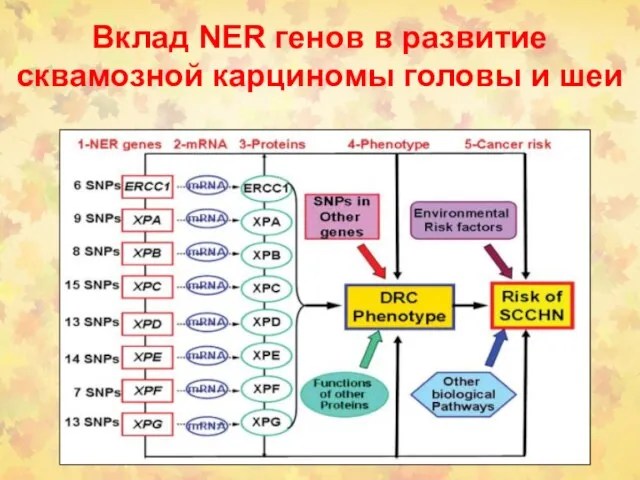
Слайд 32Вклад NER генов в развитие сквамозной карциномы головы и шеи
Вклад NER генов в развитие сквамозной карциномы головы и шеи
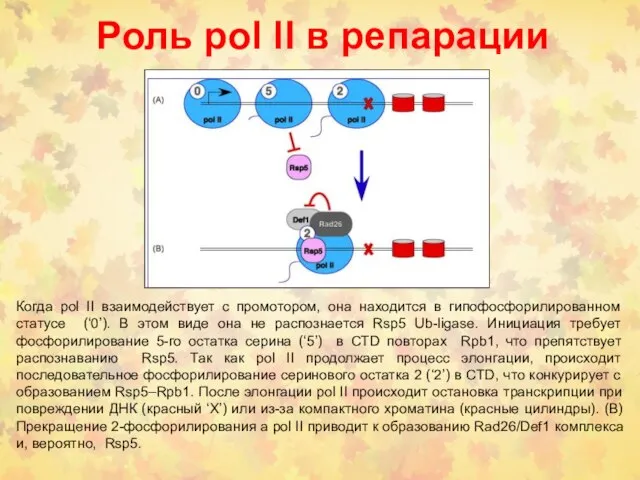
Слайд 33Роль pol lI в репарации
Когда pol II взаимодействует с промотором, она находится
Роль pol lI в репарации
Когда pol II взаимодействует с промотором, она находится
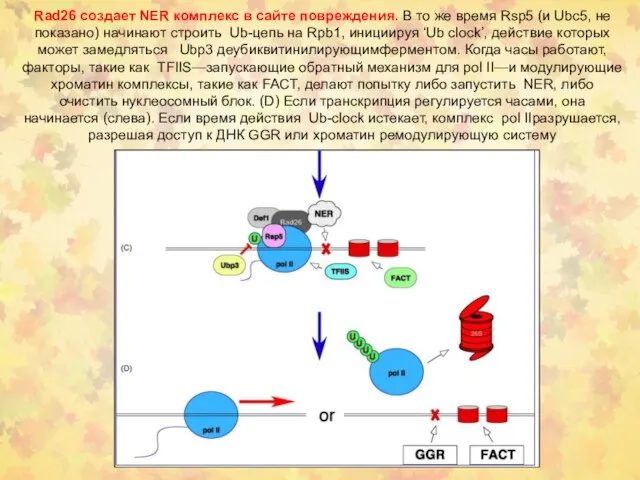
Слайд 34Rad26 создает NER комплекс в сайте повреждения. В то же время Rsp5
Rad26 создает NER комплекс в сайте повреждения. В то же время Rsp5
Слайд 35Аддукты ДНК с цис-платином
Аддукты ДНК с цис-платином
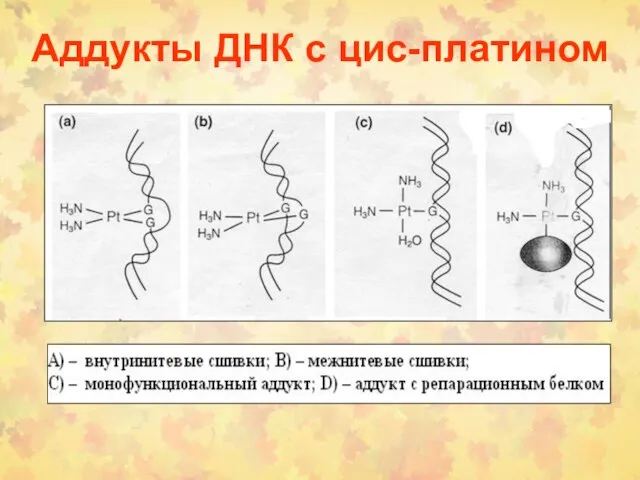
Слайд 36Репарация аддуктов ДНК с цис-платином
Репарация аддуктов ДНК с цис-платином
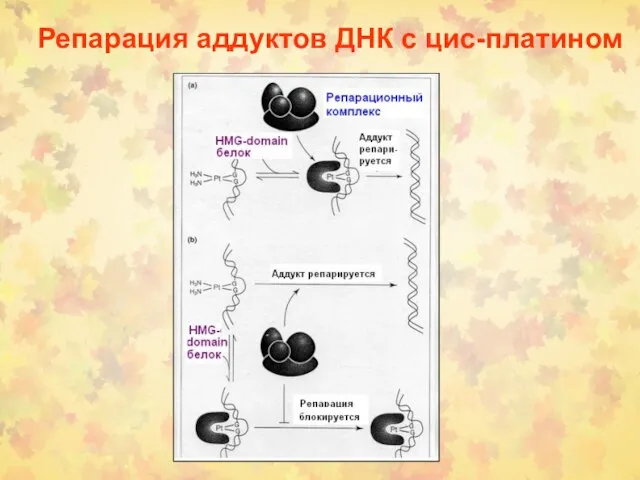
Слайд 375. Другие виды репарации ДНК
5. Другие виды репарации ДНК
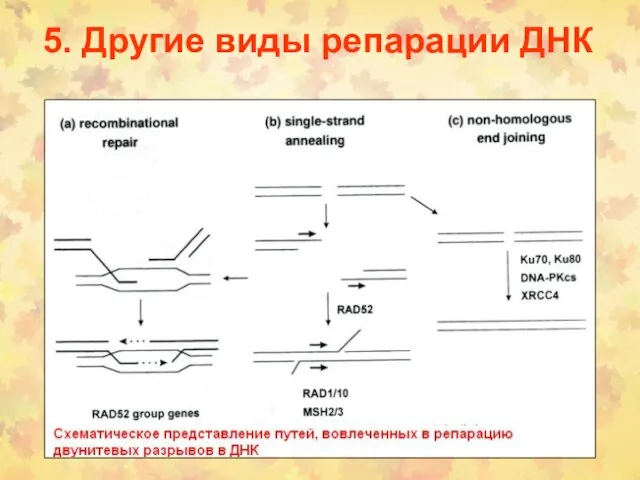
Слайд 38Альтернативный механизм репарации у дрожжей S. pombe
Альтернативный механизм репарации у дрожжей S. pombe
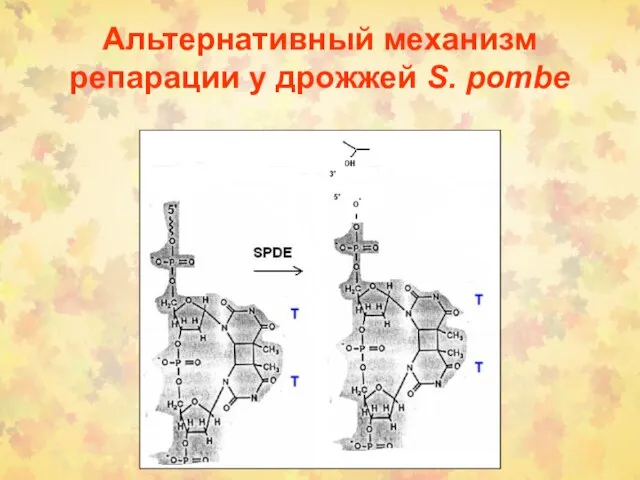
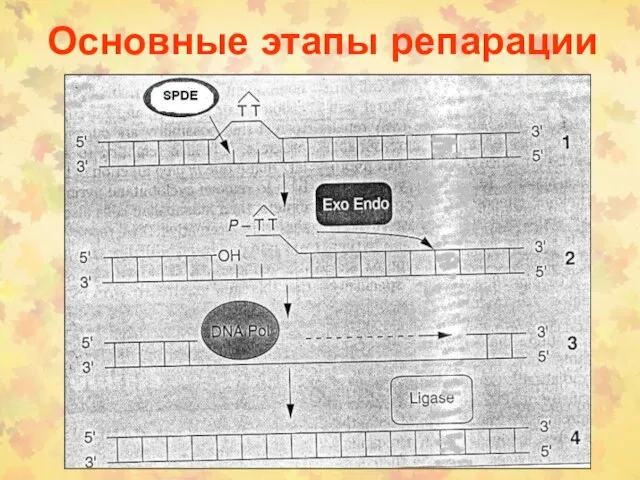
Слайд 39Основные этапы репарации
Основные этапы репарации
Слайд 40Белки ATM
ATM (="ataxia telangiectasia mutated") получила название от болезни, пациенты, среди
Белки ATM
ATM (="ataxia telangiectasia mutated") получила название от болезни, пациенты, среди

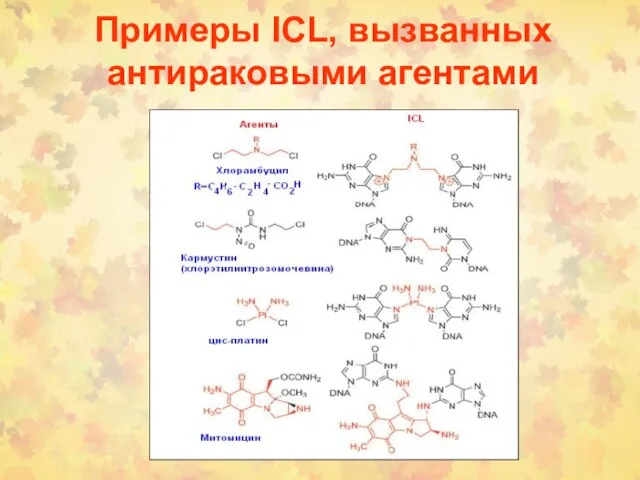
Слайд 41Примеры ICL, вызванных антираковыми агентами
Примеры ICL, вызванных антираковыми агентами
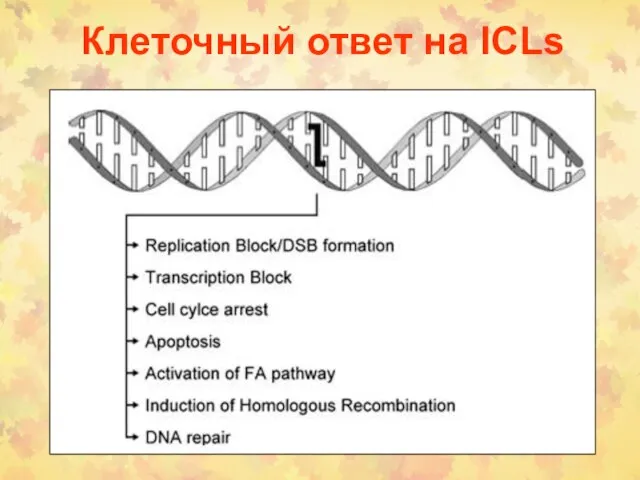
Слайд 42Клеточный ответ на ICLs
Клеточный ответ на ICLs
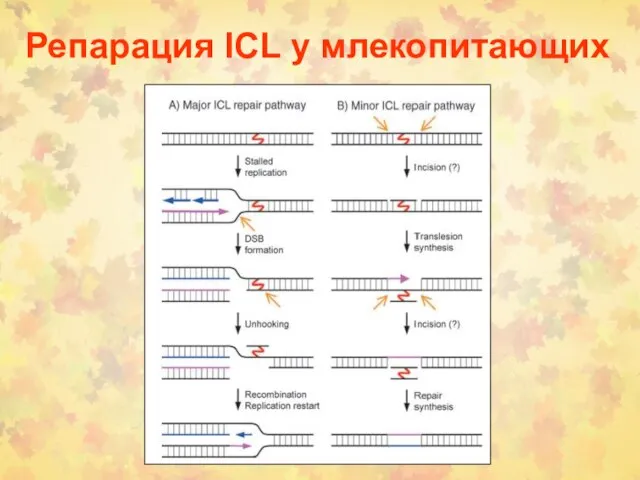
Слайд 43Репарация ICL у млекопитающих
Репарация ICL у млекопитающих
Слайд 44Fanconi Anemia (FA)путь репарации
У пациентов с FA повреждено, по крайней мере, 13
Fanconi Anemia (FA)путь репарации
У пациентов с FA повреждено, по крайней мере, 13

Слайд 45Сравнение FA генов у человека, Drosophila, Dictyostelium and C. elegans
Сравнение FA генов у человека, Drosophila, Dictyostelium and C. elegans

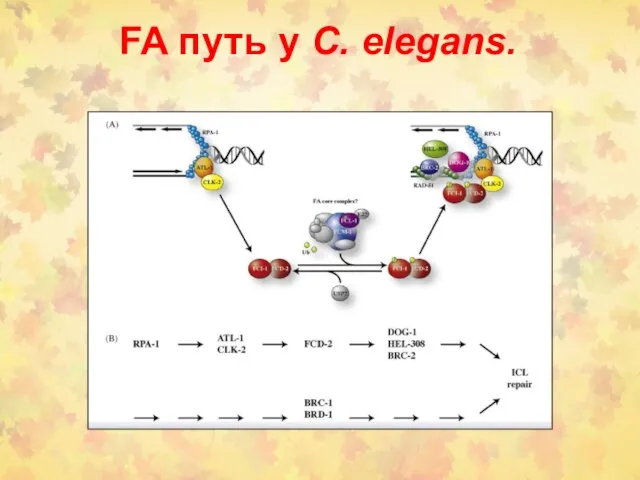
Слайд 46FA путь у C. elegans.
FA путь у C. elegans.
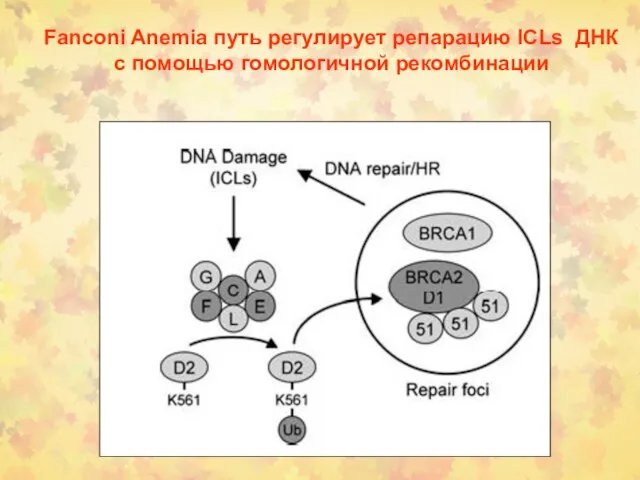
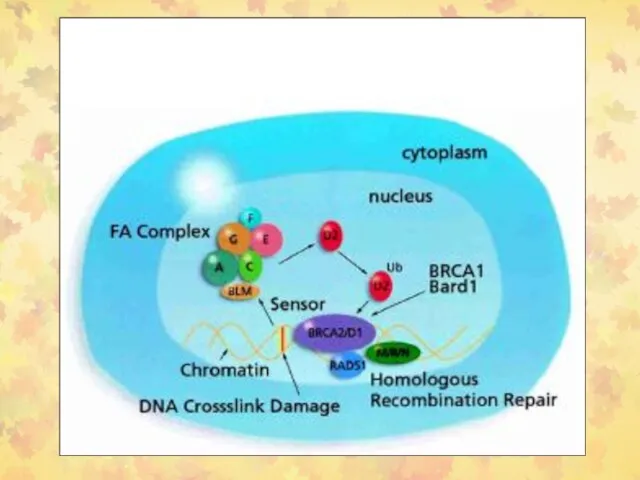
Слайд 47Fanconi Anemia путь регулирует репарацию ICLs ДНК с помощью гомологичной рекомбинации
Fanconi Anemia путь регулирует репарацию ICLs ДНК с помощью гомологичной рекомбинации
Слайд 49SUMO
(small ubiquitin-related modifier) конъюгация
Несколько SUMO E3 лигаз выявлено: SP-RING (secretory protein
SUMO
(small ubiquitin-related modifier) конъюгация
Несколько SUMO E3 лигаз выявлено: SP-RING (secretory protein

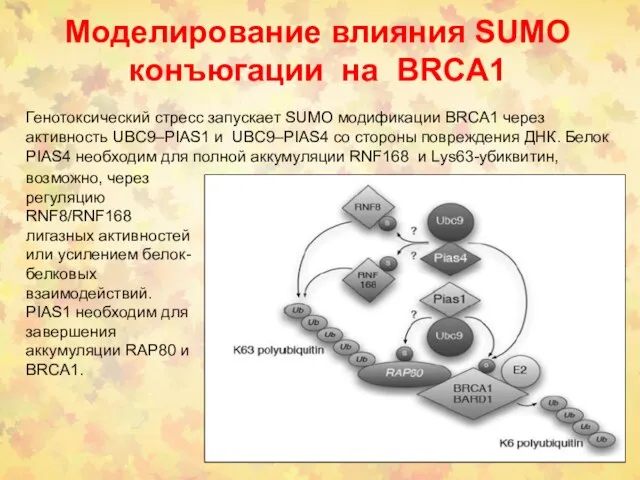
Слайд 50Моделирование влияния SUMO конъюгации на BRCA1
Генотоксический стресс запускает SUMO модификации BRCA1 через
Моделирование влияния SUMO конъюгации на BRCA1
Генотоксический стресс запускает SUMO модификации BRCA1 через
Слайд 51IY. Эпигенетические модификации ДНК
Модификации хроматина,
Метилирование ДНК,
Геномный импринтинг.
IY. Эпигенетические модификации ДНК
Модификации хроматина,
Метилирование ДНК,
Геномный импринтинг.

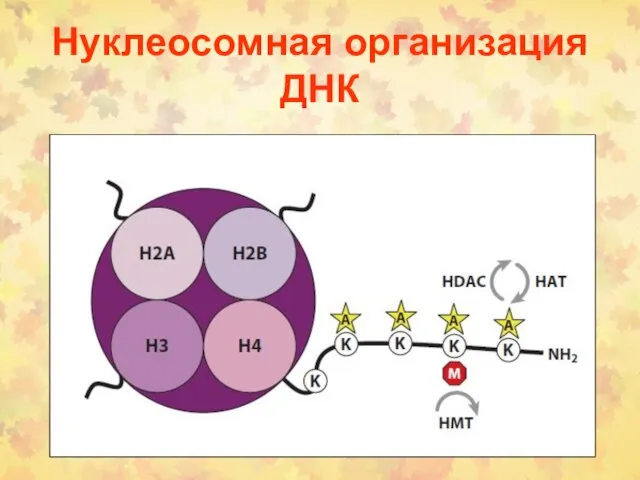
Слайд 52Нуклеосомная организация ДНК
Нуклеосомная организация ДНК
Слайд 53Регуляции транскрипции ацетилированием гистонов
Гистон-деацетилаза (HDACs) деацетилирует лизиновые остатки, создавая предпосылки для метилирования
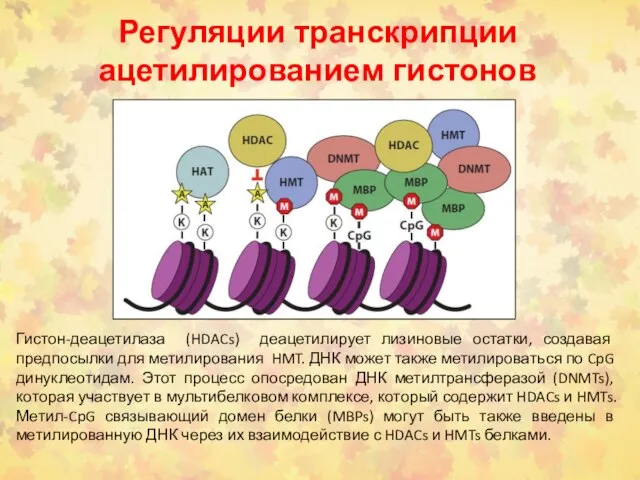
Регуляции транскрипции ацетилированием гистонов
Гистон-деацетилаза (HDACs) деацетилирует лизиновые остатки, создавая предпосылки для метилирования
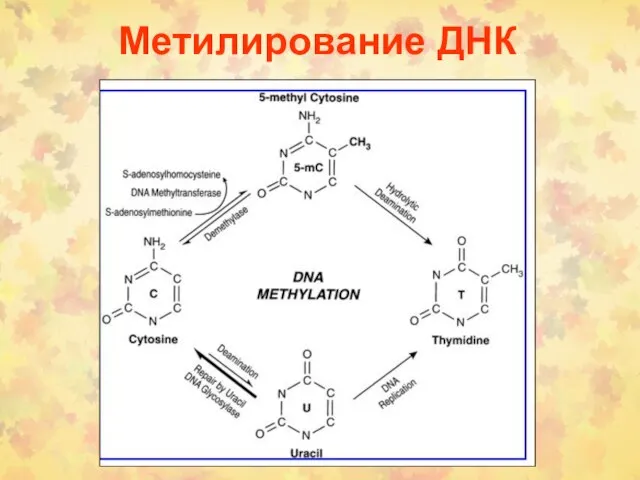
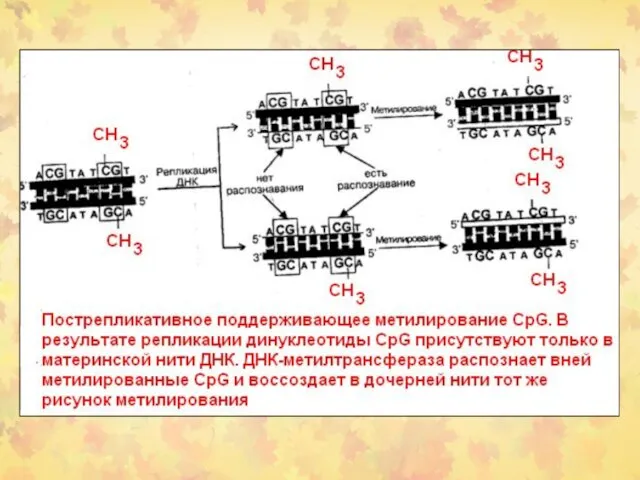
Слайд 54Метилирование ДНК
Метилирование ДНК
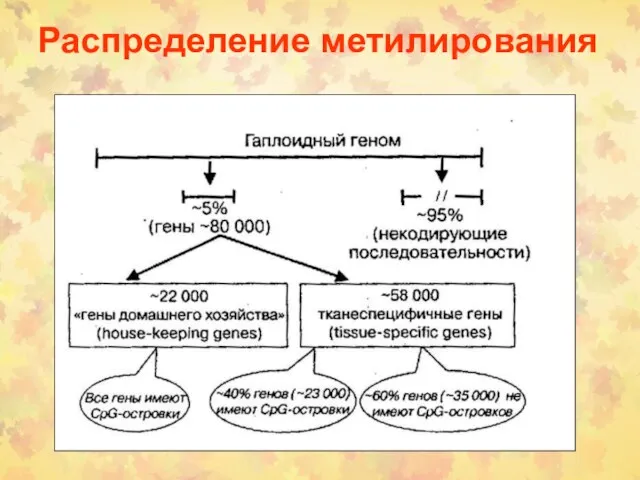
Слайд 56Распределение метилирования
Распределение метилирования
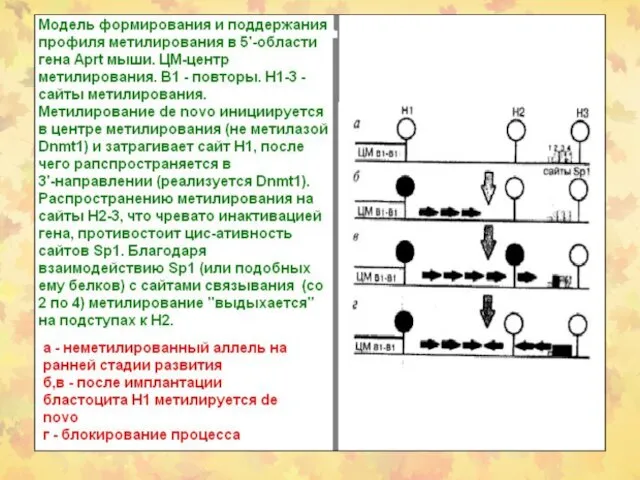
Слайд 57Статус метилирования и функциональные особенности промоторов, содержащих CpG-островки
Статус метилирования и функциональные особенности промоторов, содержащих CpG-островки

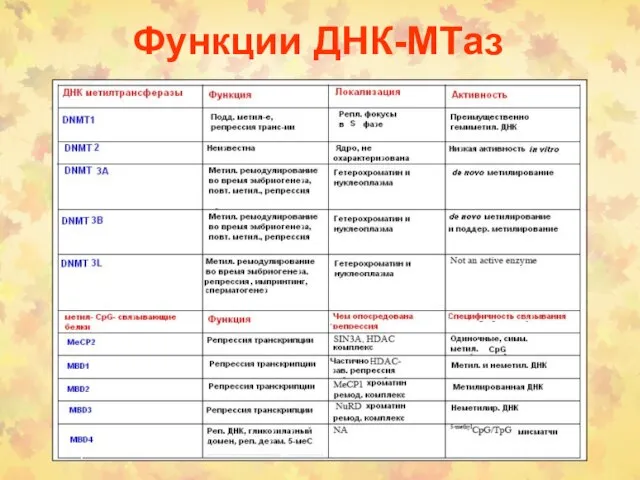
Слайд 59Функции ДНК-МТаз
Функции ДНК-МТаз
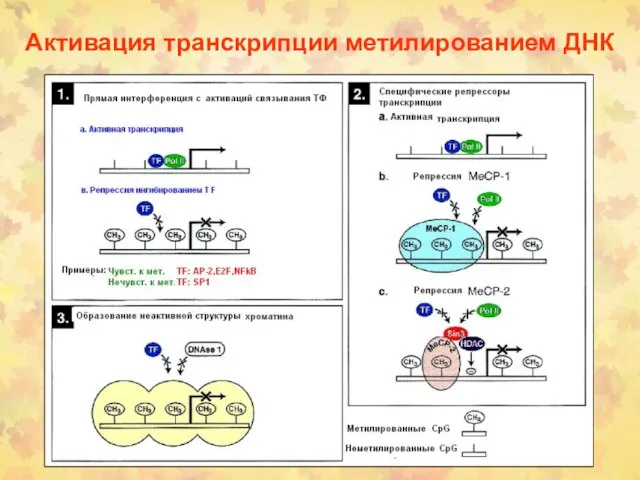
Слайд 60Активация транскрипции метилированием ДНК
Активация транскрипции метилированием ДНК

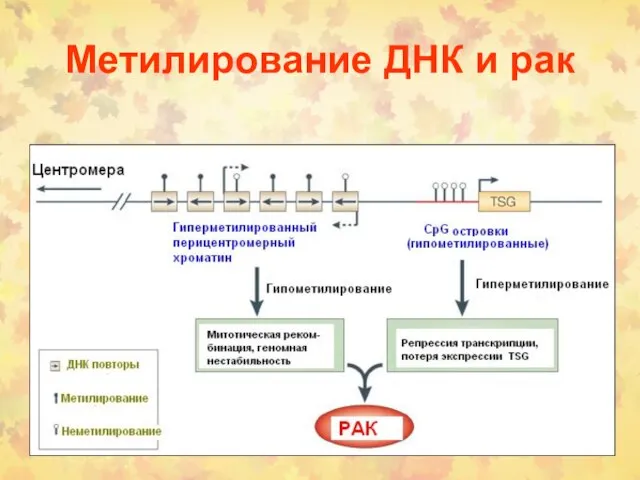
Слайд 62Метилирование ДНК и рак
Метилирование ДНК и рак
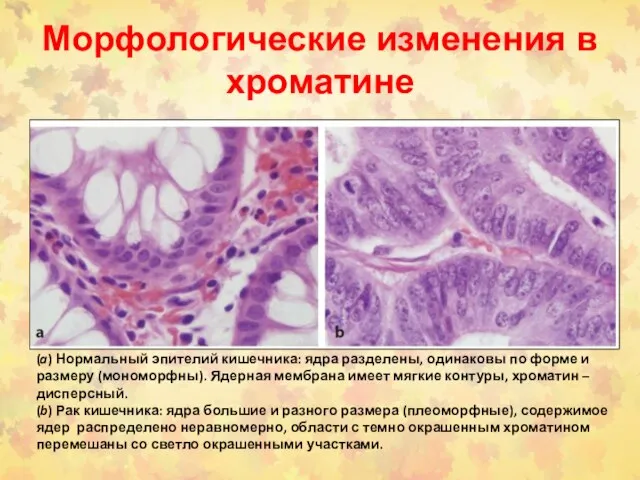
Слайд 63Морфологические изменения в хроматине
(a) Нормальный эпителий кишечника: ядра разделены, одинаковы по форме
Морфологические изменения в хроматине
(a) Нормальный эпителий кишечника: ядра разделены, одинаковы по форме
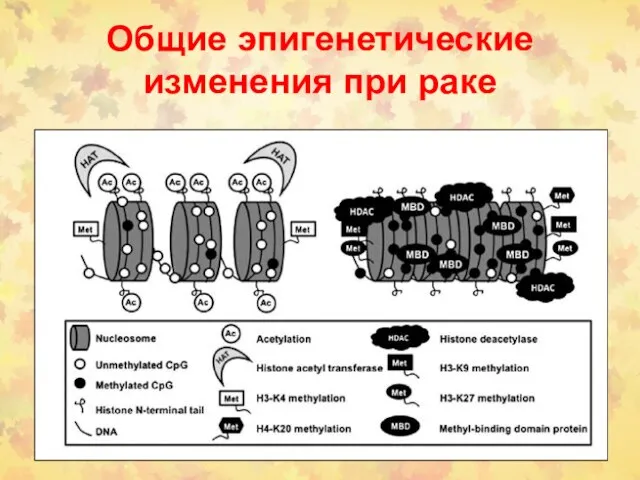
Слайд 64Общие эпигенетические изменения при раке
Общие эпигенетические изменения при раке
Слайд 65Метилирование ДНК и рак
Метилирование ДНК и рак

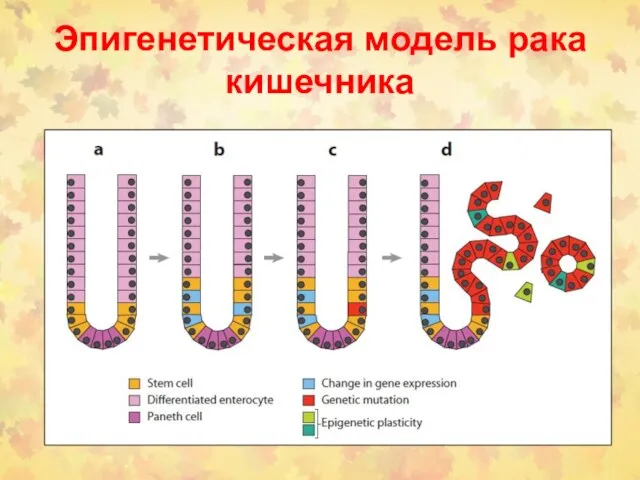
Слайд 66Эпигенетическая модель рака кишечника
Эпигенетическая модель рака кишечника
Слайд 67Примеры гиперметилирования некоторых генов
Примеры гиперметилирования некоторых генов
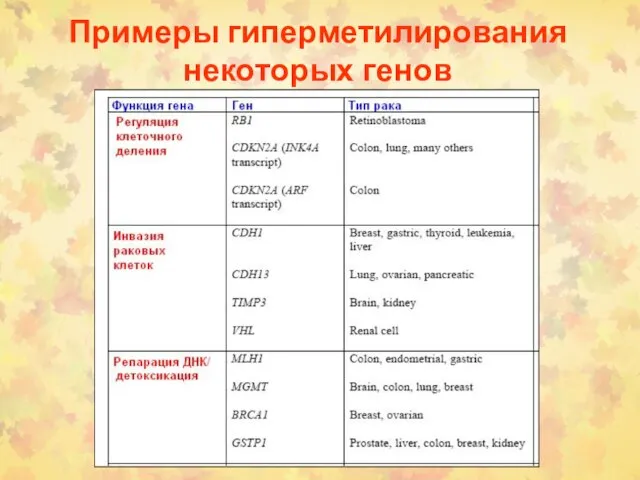
Слайд 68Примеры гиперметилирования некоторых генов
Примеры гиперметилирования некоторых генов

 Данную презентацию использовать для изучения в 11 классе темы «Излучения и спектры. Спектральный анализ» Темы рассматриваются в
Данную презентацию использовать для изучения в 11 классе темы «Излучения и спектры. Спектральный анализ» Темы рассматриваются в  Животные живого уголка
Животные живого уголка Пирин, Рила и Централен Балкан са едни от най- големите и ценни защитени територии в Европа. В тях се опазват естествени екосистеми,
Пирин, Рила и Централен Балкан са едни от най- големите и ценни защитени територии в Европа. В тях се опазват естествени екосистеми,  Музыкальные инструменты
Музыкальные инструменты Интернет – образование
Интернет – образование Ковчег
Ковчег Разработка корпоративной структуры
Разработка корпоративной структуры Gli ospiti di Jerry. Chi è?
Gli ospiti di Jerry. Chi è? Круговая теория любви А. Рейса
Круговая теория любви А. Рейса Тема 1
Тема 1 Проверочная работа по теме «Строение клетки»
Проверочная работа по теме «Строение клетки» Не храм, не золотое зданье, Не круг отобранных друзей, - Христова Церковь есть собранье Крестом искупленных людей.
Не храм, не золотое зданье, Не круг отобранных друзей, - Христова Церковь есть собранье Крестом искупленных людей. Презентация на тему Виды компьютерных вирусов Антивирусные программы
Презентация на тему Виды компьютерных вирусов Антивирусные программы  Это у нас, это гордость Вятки
Это у нас, это гордость Вятки Новогодний калейдоскоп
Новогодний калейдоскоп День космонавтики (12 апреля)
День космонавтики (12 апреля) Катаракта и современные методы лечения
Катаракта и современные методы лечения Футбол моя любимая игра
Футбол моя любимая игра Афинский Акрополь
Афинский Акрополь Презентация на тему Самопрезентация учителя
Презентация на тему Самопрезентация учителя Writing a book report
Writing a book report Возможности «1С:Зарплата и управление персоналом 8 КОРП» для автоматизации крупных предприятий и холдингов
Возможности «1С:Зарплата и управление персоналом 8 КОРП» для автоматизации крупных предприятий и холдингов Бонусная программа общероссийского профсоюза образования
Бонусная программа общероссийского профсоюза образования Презентация на тему Адам Смит
Презентация на тему Адам Смит Преодоление страхов и психолого-эмоционального напряжения средствами данстерапии
Преодоление страхов и психолого-эмоционального напряжения средствами данстерапии «Использование информационных технологий в составлении оценочных средств. Тестовые задания»
«Использование информационных технологий в составлении оценочных средств. Тестовые задания» Бизнес план магазина одежды New Style
Бизнес план магазина одежды New Style Презентация на тему MS Excel основы работы
Презентация на тему MS Excel основы работы