- Онкогенні віруси людини

Содержание
- 2. Вперше вірусну природу пухлин продемонстрували: Еритробластоз курей (Ellerman та Bang, 1908), саркома курчат (Rous, 1911); Fujinami
- 3. Peyton Rouse and the cancer causing chicken virus Rockefeller University 1912
- 4. Таксономія онкогенних вірусів DNA viruses: paріlloma-poliomaviruses- hepadnaviruses herpesviruses adenoviruses poxviruses RNA viruses: retroviruses flaviviruses
- 5. Віруси людей , що асоціюються з трансформацією HPV 16, 18, 31, 33, 45 Рак шийки матки
- 6. Як віруси трансформують клітини ? Вірусна інфекція підвищує ризик розвитку раку Діє як “мутаген” Інші ко-фактори
- 7. Докази вірусного походження пухлин Присутність частини вірусного геному в пухлині та експресія деяких вірусних генів Інфекція
- 8. Фактори, що сприяють трансформації клітин вірусами Абортивна інфекція (переривання повного циклу репродукції) інфекція повним інфекційним вірусом
- 9. Основні положення Зільбер Л.О. 1945 Віруси можуть викликати пухлини у ссавців Онкогенні віруси частіше встановлюють персистентну
- 10. Віруси рідко бувають повністю канцерогенними Між первинною інфекцією та появою пухлин – тривалий латентний період Штами
- 11. В даний час відомо більше 150 онкогенних вірусів, які поділяються на великі групи: ДНК-і РНК-вмісні РНК-вмісні
- 12. ОНКОГЕННІ РНК ВІРУСИ
- 13. Alpharetrovirus: вірус лейкозу птахів, морфологічний тип-C; (B)Betaretrovirus:вірус раку молочних залоз мишей морфологічний тип-B; (C) Gammaretrovirus: вірус
- 14. Морфологічні типи тип А – розміром 70-75 нм, ендогенні, дефектні, мають вигляд електроннощільних шарів, що локалізуються
- 15. •більшість ретротранспозонів відрізняються від РВ відсутністю позаклітинної фази • Вони мають ген env , вірусоподіні частки
- 16. Біологічні властивості РВ 1.Різний ступінь онкогенності для природніх хазяїв Високий - для вірусів саркоматозно-лейкозного комплексу; помірний
- 17. Біологічні властивості РВ 2.Здатність долати тканинні, видові та класові бар’єри, що особливо притаманно вірусам типу С
- 18. Онкогенні РНК віруси
- 19. Формування віріонів
- 20. Зворотна Транскриптаза 1970 р. Д. Балтімор, С. Міцутані та Г. Темін виявили зворотну транскриптазу в частинках
- 21. Онкогенні віруси РНК : Диплоїдна має cap- та poly-А Позитивної полярності Вірусна РНК не може транслюватись,
- 22. Онкогенні РНК віруси POL: ферменти Зворотна транскриптаза (ЗТ), РНК-аза Н Інтеграза Протеаза Звичайні ретровіруси мають 3
- 23. Вірусна РНК Зворотна транскриптаза R U5 U3 R U3 R U5 U3 R U5 Утворились LTR
- 24. Онкогенні РНК віруси головне: відмінність двох форм РНК R U5 GAG POL ENV U3 R
- 25. Для вирішення питання про природу включення геному вірусу в ДНК клітини необхідно відповісти на питання: коли,
- 26. Ендонуклеаза зшиває ланцюги, при цьому втрачається по 2 нт, формується новий 3’вкорочений кінець З”єднання 3’ кінців
- 27. Онкогенні РНК віруси Батьківська РНК РНК/ДНК гібрид Лінійний DНК/DНК дублекс Циркулярний дублекс DНК Інтеграція Реплікація Транскрипція
- 28. Механізми клітинної трансформації ретровірусами 1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси) 2) Активація онкогена ретровірусною інсерцією (cis-acting
- 29. R U5 GAG POL ENV U3 R Деякі Ретровіруси мають додатковий ген “типовий ретровірус” SRC
- 30. Вірус саркоми птахів (FSV) R U5 dGAG FMS dENV U3 R Вірус мієлоцитозу птахів (MC29) R
- 31. Клітинний ген має гомологію з геном, знайденим у трансформуюго вірусу Клітинні онкогени можуть індукувати трансформацію тільки
- 32. вірус саркоми Рауса є результатом рекомбінацій між c-src і древнім стандартним ретровірусом птахів. функції нормальних генів
- 33. Структура дефектного РВ
- 34. Гостро трансформуючі ретровіруси src Rous sarcoma virus Chicken myc Avian myelocytomatosis virus Chicken erb A, erb
- 35. Трансдукуючі ретровіруси Включення клітинного прото-онкогену в вірусний геном замість кодуючих вірусних послідовностей (виняток RSV= src- oнкоген)
- 36. Наслідки ретровірусної трансдукції “Простий удар” канцерогенезу (одна подія) Поліклональна : пухлинний ріст починається в кожній інфікованій
- 37. Retroviruses and cancer
- 39. Білкові продукти прото-онкогенів та онкогенів: ~100 різних онкогенів ідентифіковано. Всі відомі прото-онкогени залучені в позитивному контролі
- 40. Характеристика вірусних онкогенів (Virology 2000)
- 41. Трансдуковні гени – це гени сигнального шляху Зовнішні сигнальні молекули або фактори росту (sis) Клітинні рецептори
- 42. Будова та локалізація pp60-src
- 43. Активація src-гену
- 44. SRC Окисний стрес Мітоз ФР АГ Цитокіни Позаклітинний матрикс Мітотичні функції Стрес. механізм RAS MYC Реорганізація
- 45. C-SRC V-SRC
- 46. Структурна зміна в онкогені PP PP P c-Erb B (EGFR) v-Erb B Змінений v-Erb B діє
- 47. Мутації протоонкогенів Ras характерні для пухлин людини Мутації в кодонах 12, 13, 61 генів K-RAS, N-RAS,
- 48. Мутації протоонкогенів Ras викликають перманентну стимуляцію розмноження неопластичних клітин Ras Ras ГДФ ГТФ SOS GAP Ras
- 49. 1. Sis – фактор росту тромбоцитів (PDGF) cДНК диких мавп та котів викликає саркому Трансформовані клітини
- 50. інтегрини (a5b1 и др) Рецептори мітогенів/мотогенів (Met.) PI3K Raf ERK Rho Ras MEK Rac PAK MEKK
- 51. За функціональною активністю онкогени можна розділити на групи 1.а) з тирозинкіназною активністю : abl, erb B,
- 52. Онкогени як сигнали трансдукції Фактор росту v-sis, int-1, int-2, hst, fgf-5 Рецептори фактору росту v-erb-B, v-fms,
- 53. Існує три гіпотези щодо можливих механізмів неопластичного перетворення клітин за допомогою онкогенів: -«кількісна» - збільшення кількості
- 55. Механізми клітинної трансформації ретровірусами 1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси) 2) Активація онкогена ретровірусною інсерцією (cis-acting
- 56. Cis-активуючі ретровіруси Не мають онкогена Зберігають всі вірусні гени Компетентні за реплікацією
- 57. Повільно трансформуючі (хронічно) ретровіруси викликають пухлини протягом тривалого латентного періоду Немає онкогена! – Яким чином утворюється
- 58. Пухлина походить з однієї клітини - моноклональна ALV може інтегруватись в геном клітини БAГАТЬОХ місцях Але
- 59. Чи специфічні ці сайти? Вірус Myelocytoma виділений від різних птахів завжди знаходять в одному й тому
- 60. Як C-onc залучаються в не вірусний канцерогенез?
- 61. From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006 Pearson Benjamin Cummings.
- 62. Протоонкогени та перебудова хромомсом Лімфома Беркітта t(8;14), t(8;22), t(2;8) - мішень c-myc. інсерція промотора Ig тяжкого
- 63. Механізм клітинної трансфорації cis-активуючих ретровірусів Випадкова ретровірусна інтеграція в клітинний геном Інсерційна активація (або дезактивація) с-onc
- 65. Наслідки активації онкогену при інсерції Ретровірусів Клітинна трансформація відбувається завдяки інсерції поблизу потенційного онкогена Пухлини моноклональні:
- 66. Механізми клітинної трансформації ретровірусами 1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси) 2) Активація онкогена ретровірусною інсерцією (cis-acting
- 67. Human T cell Leukemia Virus type I (HTLV-I) Асоційований з 2 фатальними хворобами людей Т-клітинний лейкоз
- 68. HTLV-I та ATL 1980 Галло ізолював тип C retrovirus (HTLV1) від хворого з “різкою T клітинною
- 69. Міжклітинний контакт Статевий (60% від чоловіка до жінки проти 1% від жінки до чоловіка) Продуктами крові
- 70. Онкогенез, опосередкований важливим ретровірусним білком Виключення з парадігми ретровірусного онкогенезу (HTLV-1) HTLV-1 не має послідовностей клітинного
- 71. HTLV-I genome 9 kilobase РНК геном Унікальні регуляторні білки Tax та Rev Важливі для вірусної реплікації
- 72. Tax тa Oнкогенез Tax необхідний для вірусної реплікації 40kda фосфопротеїн Tранскрипційний активатор геному HTLV-I Взаємодія з
- 73. Tax – множинний трансактиватор Зв”язування з клітинними транскрипційними факторами посилює їх взаємодію з клітинними промоторами Дисоціює
- 74. Білки регулятори клітинного циклу – мішені дії Tax Iнактивація p53 (G1/S контроль рестрикції) Активує циклін D,
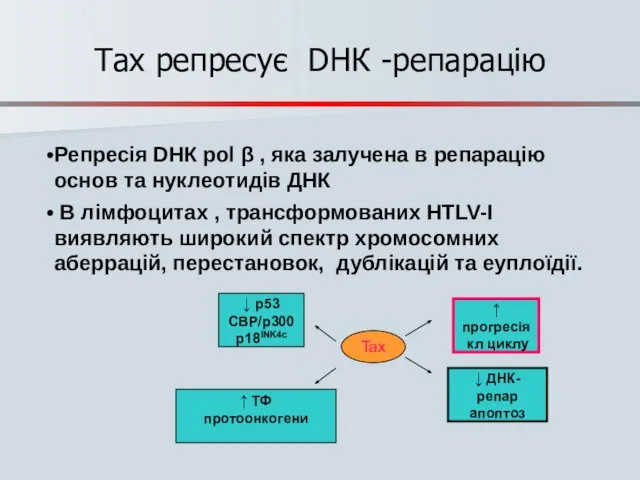
- 75. Tax репресує DНК -репарацію Репресія DНК pol β , яка залучена в репарацію основ та нуклеотидів
- 76. Механізми клітинної трансформації ретровірусами
- 77. Онкогенні РНК- віруси
- 79. Скачать презентацию
Слайд 2 Вперше вірусну природу пухлин продемонстрували:
Еритробластоз курей (Ellerman та Bang, 1908),
Вперше вірусну природу пухлин продемонстрували:
Еритробластоз курей (Ellerman та Bang, 1908),

Слайд 3Peyton Rouse and the cancer causing chicken virus
Rockefeller University 1912
Peyton Rouse and the cancer causing chicken virus
Rockefeller University 1912

Слайд 4Таксономія онкогенних вірусів
DNA viruses: paріlloma-poliomaviruses- hepadnaviruses herpesviruses adenoviruses poxviruses
RNA viruses:
Таксономія онкогенних вірусів
DNA viruses: paріlloma-poliomaviruses- hepadnaviruses herpesviruses adenoviruses poxviruses
RNA viruses:

Слайд 5Віруси людей , що асоціюються з трансформацією
HPV 16, 18, 31, 33,
Віруси людей , що асоціюються з трансформацією
HPV 16, 18, 31, 33,

Слайд 6Як віруси трансформують клітини ?
Вірусна інфекція підвищує ризик розвитку раку
Діє
Як віруси трансформують клітини ?
Вірусна інфекція підвищує ризик розвитку раку
Діє

Слайд 7Докази вірусного походження пухлин
Присутність частини вірусного геному в пухлині та експресія
Докази вірусного походження пухлин
Присутність частини вірусного геному в пухлині та експресія

Слайд 8Фактори, що сприяють трансформації клітин вірусами
Абортивна інфекція (переривання повного циклу репродукції)
інфекція повним
Фактори, що сприяють трансформації клітин вірусами
Абортивна інфекція (переривання повного циклу репродукції)
інфекція повним

Слайд 9Основні положення
Зільбер Л.О. 1945
Віруси можуть викликати пухлини у ссавців
Онкогенні віруси частіше
Основні положення
Зільбер Л.О. 1945
Віруси можуть викликати пухлини у ссавців
Онкогенні віруси частіше

Слайд 10Віруси рідко бувають повністю канцерогенними
Між первинною інфекцією та появою пухлин – тривалий
Віруси рідко бувають повністю канцерогенними
Між первинною інфекцією та появою пухлин – тривалий

Слайд 11В даний час відомо більше 150 онкогенних вірусів, які поділяються на великі
В даний час відомо більше 150 онкогенних вірусів, які поділяються на великі

Слайд 12ОНКОГЕННІ РНК ВІРУСИ
ОНКОГЕННІ РНК ВІРУСИ

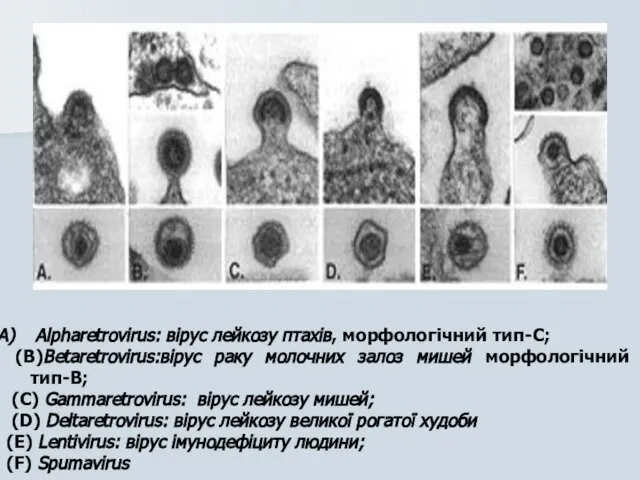
Слайд 13Alpharetrovirus: вірус лейкозу птахів, морфологічний тип-C;
(B)Betaretrovirus:вірус раку молочних залоз мишей морфологічний
(B)Betaretrovirus:вірус раку молочних залоз мишей морфологічний
Слайд 14Морфологічні типи
тип А – розміром 70-75 нм, ендогенні, дефектні, мають вигляд електроннощільних
Морфологічні типи
тип А – розміром 70-75 нм, ендогенні, дефектні, мають вигляд електроннощільних

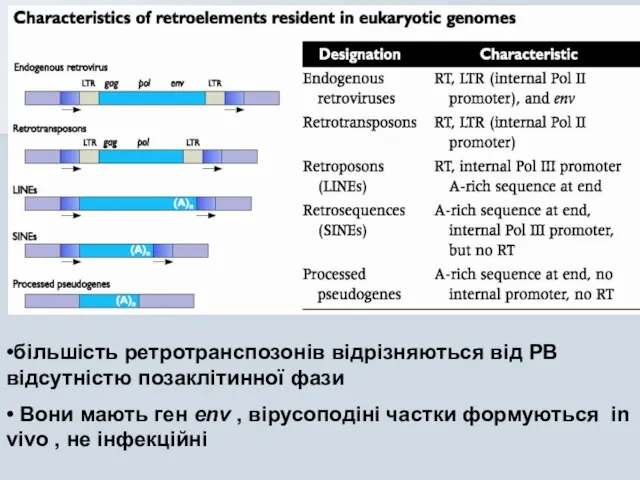
Слайд 15•більшість ретротранспозонів відрізняються від РВ відсутністю позаклітинної фази
• Вони мають ген env
•більшість ретротранспозонів відрізняються від РВ відсутністю позаклітинної фази
• Вони мають ген env
Слайд 16Біологічні властивості РВ
1.Різний ступінь онкогенності для природніх хазяїв Високий - для вірусів
Біологічні властивості РВ
1.Різний ступінь онкогенності для природніх хазяїв Високий - для вірусів

Слайд 17Біологічні властивості РВ
2.Здатність долати тканинні, видові та класові бар’єри, що особливо притаманно
Біологічні властивості РВ
2.Здатність долати тканинні, видові та класові бар’єри, що особливо притаманно

Слайд 18Онкогенні РНК віруси
Онкогенні РНК віруси
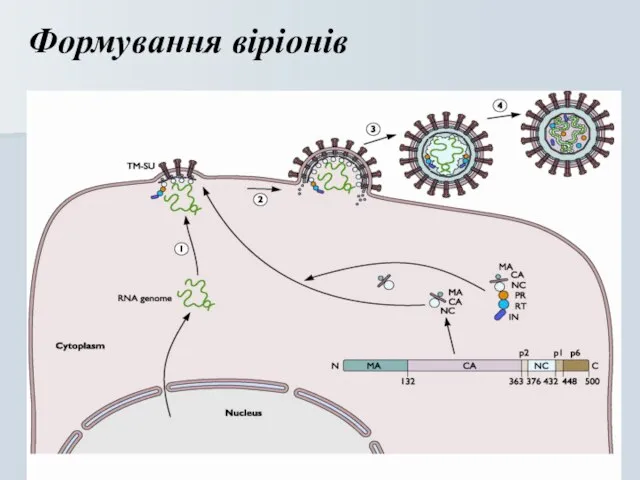
Слайд 19Формування віріонів
Формування віріонів
Слайд 20Зворотна Транскриптаза
1970 р.
Д. Балтімор, С. Міцутані та Г. Темін виявили зворотну
Зворотна Транскриптаза
1970 р.
Д. Балтімор, С. Міцутані та Г. Темін виявили зворотну

Слайд 21Онкогенні віруси
РНК :
Диплоїдна має cap- та poly-А
Позитивної полярності
Вірусна РНК не
Онкогенні віруси
РНК :
Диплоїдна має cap- та poly-А
Позитивної полярності
Вірусна РНК не

Слайд 22Онкогенні РНК віруси
POL: ферменти
Зворотна транскриптаза (ЗТ), РНК-аза Н
Інтеграза
Протеаза
Звичайні ретровіруси мають
3
Онкогенні РНК віруси
POL: ферменти
Зворотна транскриптаза (ЗТ), РНК-аза Н
Інтеграза
Протеаза
Звичайні ретровіруси мають
3

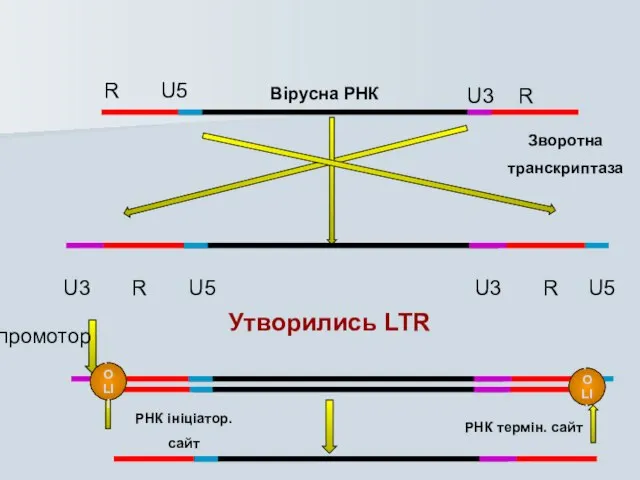
Слайд 23Вірусна РНК
Зворотна
транскриптаза
R U5
U3 R
U3 R U5
U3 R U5
Утворились LTR
POLII
Вірусна РНК
Зворотна
транскриптаза
R U5
U3 R
U3 R U5
U3 R U5
Утворились LTR
POLII
Слайд 24Онкогенні РНК віруси
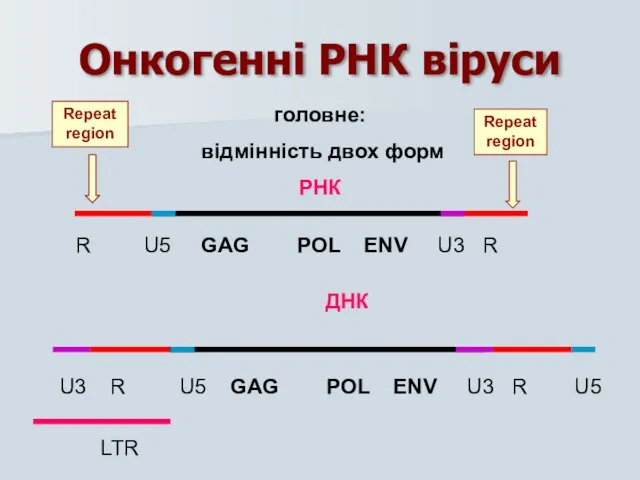
головне:
відмінність двох форм
РНК
R U5 GAG POL ENV U3 R
Онкогенні РНК віруси
головне:
відмінність двох форм
РНК
R U5 GAG POL ENV U3 R
Слайд 25Для вирішення питання про природу включення геному вірусу в ДНК клітини необхідно
Для вирішення питання про природу включення геному вірусу в ДНК клітини необхідно

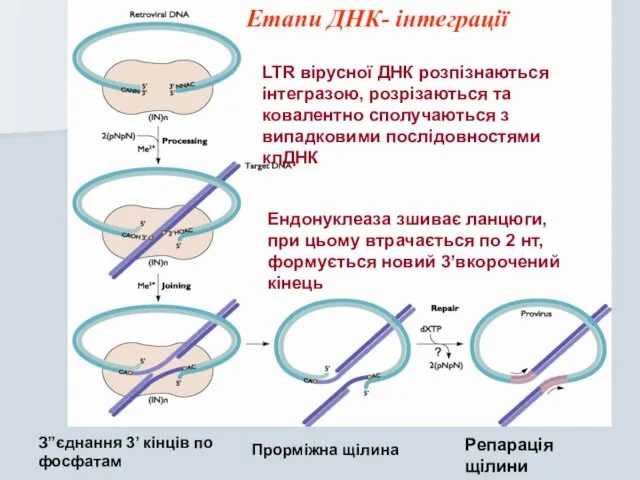
Слайд 26Ендонуклеаза зшиває ланцюги, при цьому втрачається по 2 нт, формується новий 3’вкорочений
Ендонуклеаза зшиває ланцюги, при цьому втрачається по 2 нт, формується новий 3’вкорочений
Слайд 27Онкогенні РНК віруси
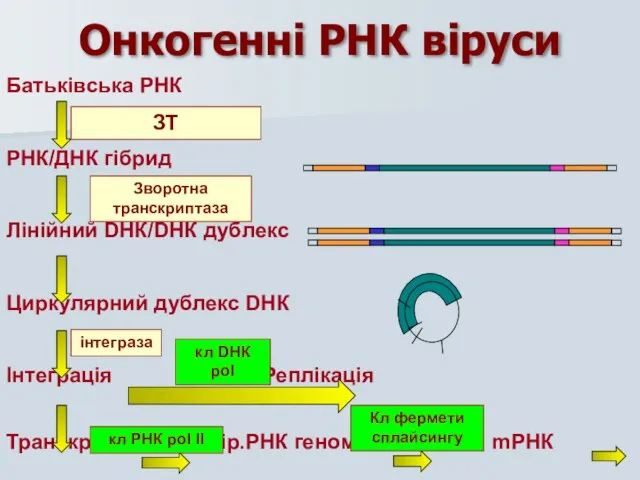
Батьківська РНК
РНК/ДНК гібрид
Лінійний DНК/DНК дублекс
Циркулярний дублекс DНК
Інтеграція Реплікація
Транскрипція вір.РНК
Онкогенні РНК віруси
Батьківська РНК
РНК/ДНК гібрид
Лінійний DНК/DНК дублекс
Циркулярний дублекс DНК
Інтеграція Реплікація
Транскрипція вір.РНК
Слайд 28Механізми клітинної трансформації ретровірусами
1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси)
2) Активація онкогена ретровірусною
Механізми клітинної трансформації ретровірусами
1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси)
2) Активація онкогена ретровірусною

Слайд 29 R U5 GAG POL ENV U3 R
Деякі Ретровіруси мають додатковий ген
“типовий
R U5 GAG POL ENV U3 R
Деякі Ретровіруси мають додатковий ген
“типовий

Слайд 30Вірус саркоми птахів (FSV)
R U5 dGAG FMS dENV U3 R
Вірус мієлоцитозу
Вірус саркоми птахів (FSV)
R U5 dGAG FMS dENV U3 R
Вірус мієлоцитозу

Слайд 31Клітинний ген має гомологію з геном, знайденим у трансформуюго вірусу
Клітинні онкогени можуть
Клітинний ген має гомологію з геном, знайденим у трансформуюго вірусу
Клітинні онкогени можуть

Слайд 32
вірус саркоми Рауса є результатом рекомбінацій між c-src і древнім стандартним ретровірусом
вірус саркоми Рауса є результатом рекомбінацій між c-src і древнім стандартним ретровірусом

Слайд 33Структура дефектного РВ
Структура дефектного РВ

Слайд 34Гостро трансформуючі ретровіруси
src Rous sarcoma virus Chicken
myc Avian myelocytomatosis virus Chicken
erb A, erb B Avian
Гостро трансформуючі ретровіруси
src Rous sarcoma virus Chicken
myc Avian myelocytomatosis virus Chicken
erb A, erb B Avian

Слайд 35Трансдукуючі ретровіруси
Включення клітинного прото-онкогену в вірусний геном замість кодуючих вірусних послідовностей (виняток
Трансдукуючі ретровіруси
Включення клітинного прото-онкогену в вірусний геном замість кодуючих вірусних послідовностей (виняток
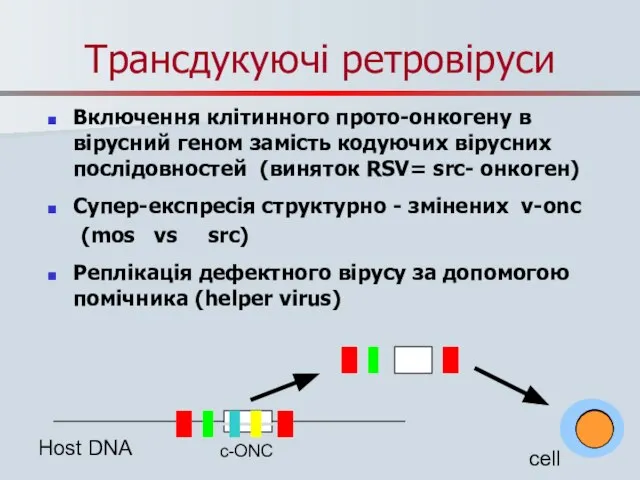
Слайд 36Наслідки ретровірусної трансдукції
“Простий удар” канцерогенезу (одна подія)
Поліклональна : пухлинний ріст починається в
Наслідки ретровірусної трансдукції
“Простий удар” канцерогенезу (одна подія)
Поліклональна : пухлинний ріст починається в

Слайд 37Retroviruses and cancer
Retroviruses and cancer
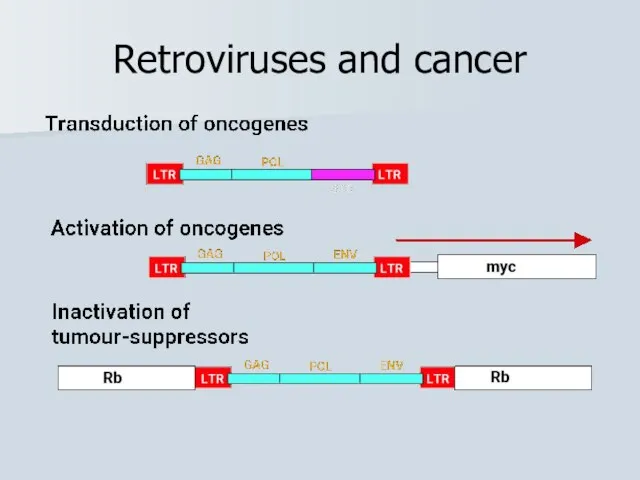
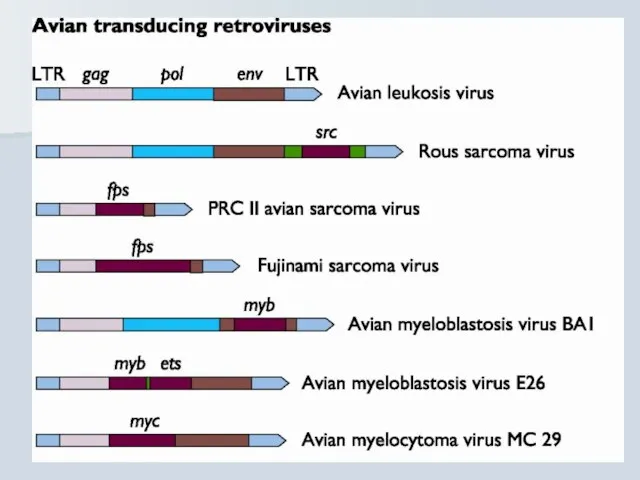
Слайд 39Білкові продукти прото-онкогенів та онкогенів:
~100 різних онкогенів ідентифіковано.
Всі відомі прото-онкогени залучені в
Білкові продукти прото-онкогенів та онкогенів:
~100 різних онкогенів ідентифіковано.
Всі відомі прото-онкогени залучені в

Слайд 40Характеристика вірусних онкогенів (Virology 2000)
Характеристика вірусних онкогенів (Virology 2000)
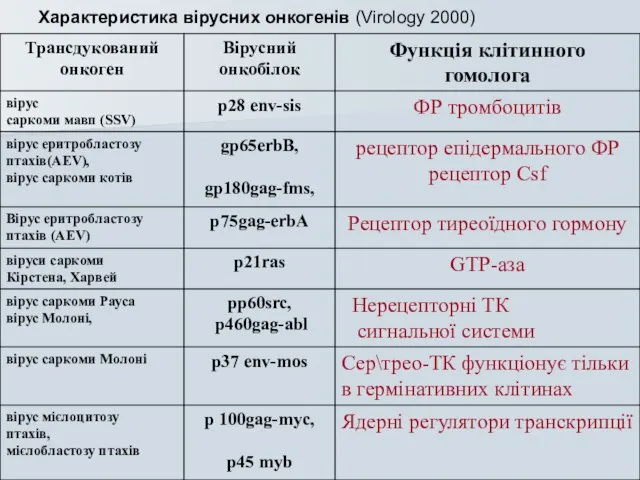
Слайд 41 Трансдуковні гени – це гени сигнального шляху
Зовнішні сигнальні молекули або фактори
Трансдуковні гени – це гени сигнального шляху
Зовнішні сигнальні молекули або фактори

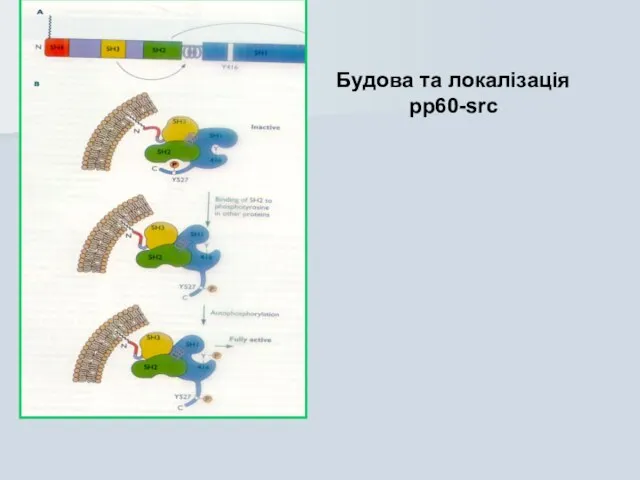
Слайд 42Будова та локалізація pp60-src
Будова та локалізація pp60-src
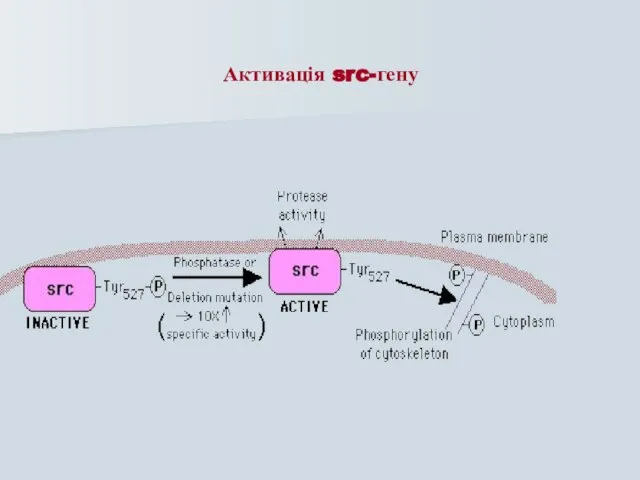
Слайд 43Активація src-гену
Активація src-гену
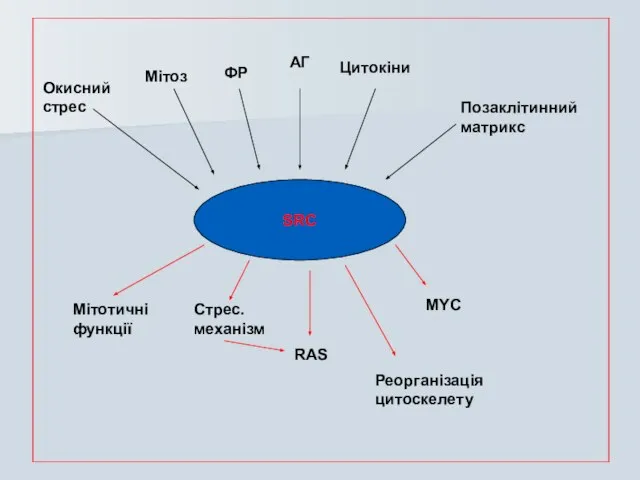
Слайд 44SRC
Окисний
стрес
Мітоз
ФР
АГ
Цитокіни
Позаклітинний
матрикс
Мітотичні
функції
Стрес.
механізм
RAS
MYC
Реорганізація
цитоскелету
SRC
Окисний
стрес
Мітоз
ФР
АГ
Цитокіни
Позаклітинний
матрикс
Мітотичні
функції
Стрес.
механізм
RAS
MYC
Реорганізація
цитоскелету
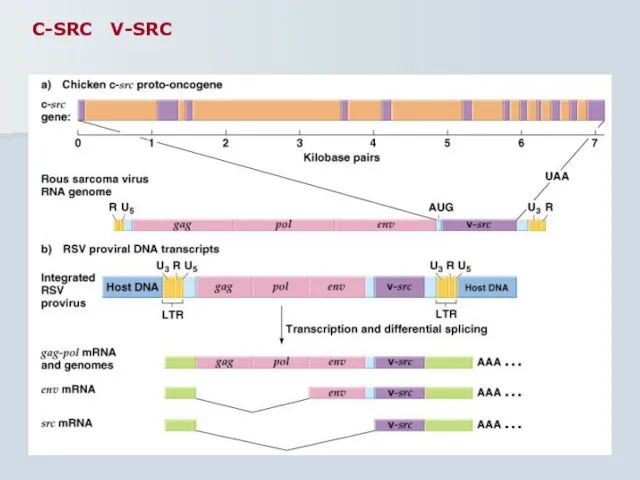
Слайд 45C-SRC V-SRC
C-SRC V-SRC
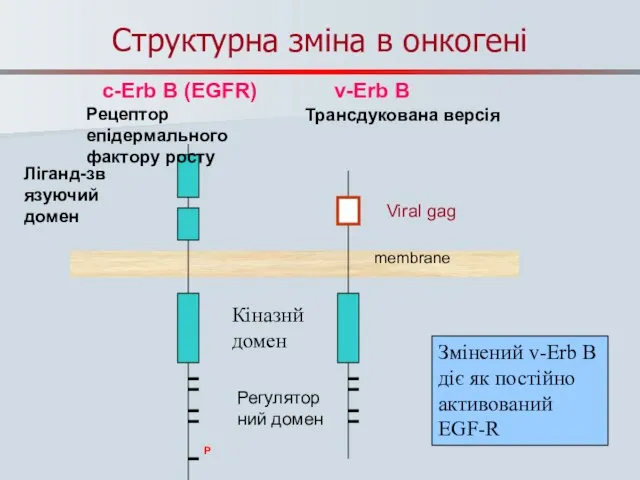
Слайд 46Структурна зміна в онкогені
PP
PP
P
c-Erb B (EGFR) v-Erb B
Змінений v-Erb B діє як
Структурна зміна в онкогені
PP
PP
P
c-Erb B (EGFR) v-Erb B
Змінений v-Erb B діє як
Слайд 47 Мутації протоонкогенів Ras
характерні для пухлин людини
Мутації в кодонах 12, 13,
Мутації протоонкогенів Ras
характерні для пухлин людини
Мутації в кодонах 12, 13,

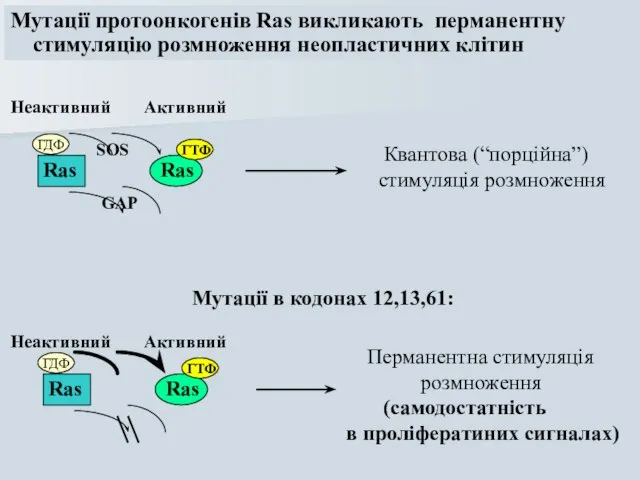
Слайд 48Мутації протоонкогенів Ras викликають перманентну
стимуляцію розмноження неопластичних клітин
Ras
Ras
ГДФ
ГТФ
SOS
GAP
Ras
Ras
ГДФ
ГТФ
Мутації в кодонах 12,13,61:
Мутації протоонкогенів Ras викликають перманентну
стимуляцію розмноження неопластичних клітин
Ras
Ras
ГДФ
ГТФ
SOS
GAP
Ras
Ras
ГДФ
ГТФ
Мутації в кодонах 12,13,61:
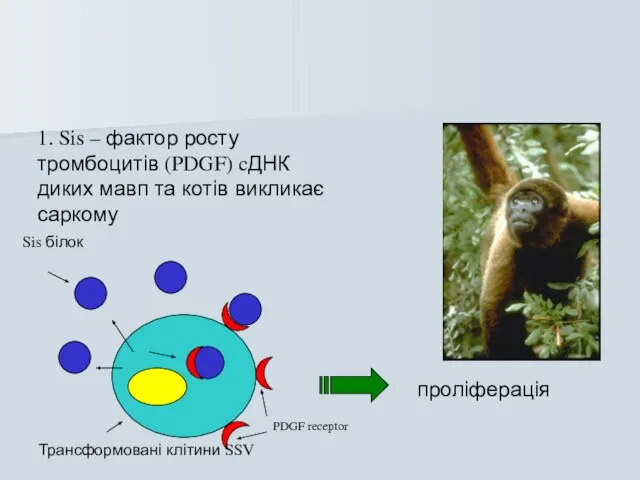
Слайд 491. Sis – фактор росту тромбоцитів (PDGF) cДНК диких мавп та котів
1. Sis – фактор росту тромбоцитів (PDGF) cДНК диких мавп та котів
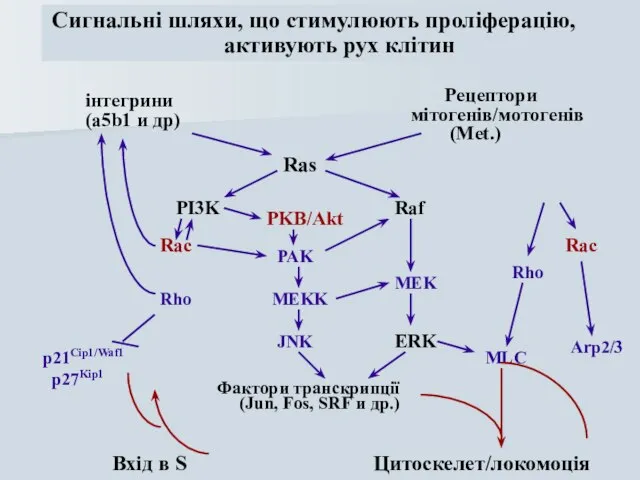
Слайд 50інтегрини
(a5b1 и др)
Рецептори
мітогенів/мотогенів
(Met.)
PI3K
Raf
ERK
Rho
Ras
MEK
Rac
PAK
MEKK
JNK
Фактори транскрипції
(Jun, Fos, SRF и др.)
p21Cip1/Waf1
інтегрини
(a5b1 и др)
Рецептори
мітогенів/мотогенів
(Met.)
PI3K
Raf
ERK
Rho
Ras
MEK
Rac
PAK
MEKK
JNK
Фактори транскрипції
(Jun, Fos, SRF и др.)
p21Cip1/Waf1
Слайд 51За функціональною активністю онкогени можна розділити на групи
1.а) з тирозинкіназною активністю :
За функціональною активністю онкогени можна розділити на групи
1.а) з тирозинкіназною активністю :

Слайд 52
Онкогени як сигнали трансдукції
Фактор росту
v-sis, int-1, int-2, hst, fgf-5
Рецептори фактору росту
v-erb-B, v-fms,
Онкогени як сигнали трансдукції
Фактор росту
v-sis, int-1, int-2, hst, fgf-5
Рецептори фактору росту
v-erb-B, v-fms,

Слайд 53Існує три гіпотези щодо можливих механізмів неопластичного перетворення клітин за допомогою онкогенів:
-«кількісна»
Існує три гіпотези щодо можливих механізмів неопластичного перетворення клітин за допомогою онкогенів:
-«кількісна»


Слайд 55Механізми клітинної трансформації ретровірусами
1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси)
2) Активація онкогена ретровірусною
Механізми клітинної трансформації ретровірусами
1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси)
2) Активація онкогена ретровірусною

Слайд 56Cis-активуючі ретровіруси
Не мають онкогена
Зберігають всі вірусні гени
Компетентні за реплікацією
Cis-активуючі ретровіруси
Не мають онкогена
Зберігають всі вірусні гени
Компетентні за реплікацією

Слайд 57Повільно трансформуючі (хронічно) ретровіруси викликають пухлини протягом тривалого латентного періоду
Немає онкогена! –
Повільно трансформуючі (хронічно) ретровіруси викликають пухлини протягом тривалого латентного періоду
Немає онкогена! –

Слайд 58Пухлина походить з однієї клітини - моноклональна
ALV може інтегруватись в геном клітини
Пухлина походить з однієї клітини - моноклональна
ALV може інтегруватись в геном клітини

Слайд 59Чи специфічні ці сайти?
Вірус Myelocytoma виділений від різних птахів завжди знаходять
Чи специфічні ці сайти?
Вірус Myelocytoma виділений від різних птахів завжди знаходять

Слайд 60Як C-onc залучаються в не вірусний канцерогенез?
Як C-onc залучаються в не вірусний канцерогенез?

Слайд 61From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006 Pearson
From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006 Pearson
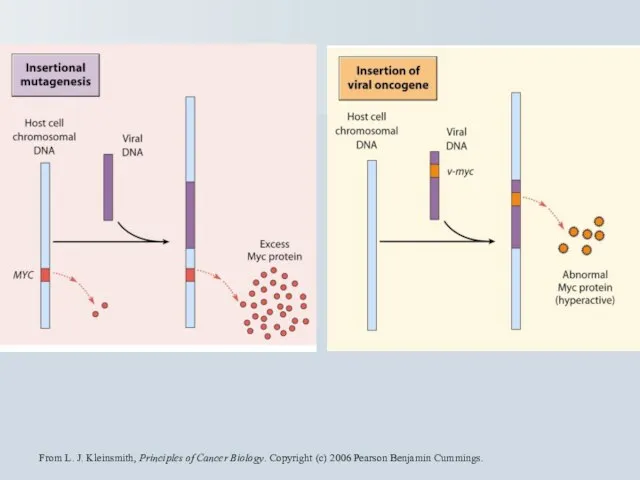
Слайд 62Протоонкогени та перебудова хромомсом
Лімфома Беркітта t(8;14), t(8;22), t(2;8) - мішень c-myc.
інсерція
Протоонкогени та перебудова хромомсом
Лімфома Беркітта t(8;14), t(8;22), t(2;8) - мішень c-myc.
інсерція

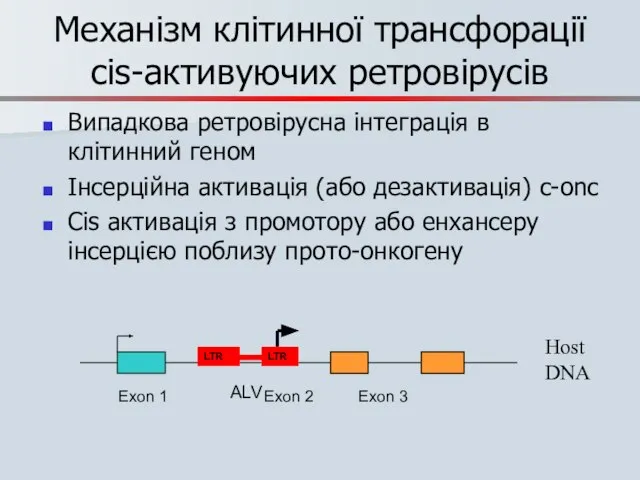
Слайд 63Механізм клітинної трансфорації cis-активуючих ретровірусів
Випадкова ретровірусна інтеграція в клітинний геном
Інсерційна активація (або
Механізм клітинної трансфорації cis-активуючих ретровірусів
Випадкова ретровірусна інтеграція в клітинний геном
Інсерційна активація (або
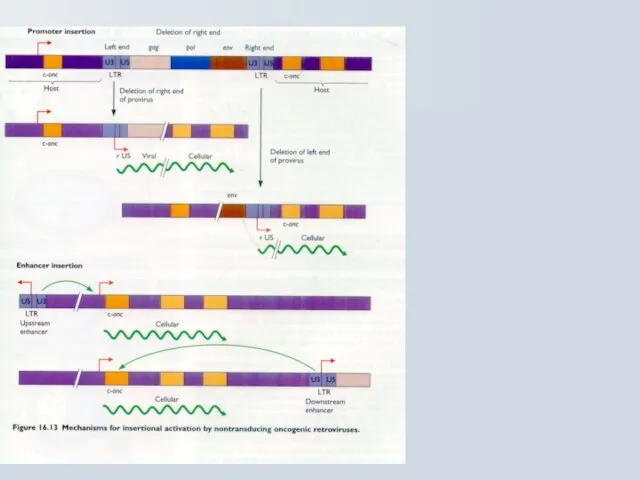
Слайд 65Наслідки активації онкогену при інсерції Ретровірусів
Клітинна трансформація відбувається завдяки інсерції
Наслідки активації онкогену при інсерції Ретровірусів
Клітинна трансформація відбувається завдяки інсерції

Слайд 66Механізми клітинної трансформації ретровірусами
1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси)
2) Активація онкогена ретровірусною
Механізми клітинної трансформації ретровірусами
1) Ретровірусна трансдукція онкогену (трансдукуючі ретровіруси)
2) Активація онкогена ретровірусною

Слайд 67Human T cell Leukemia Virus type I (HTLV-I)
Асоційований з 2 фатальними хворобами
Human T cell Leukemia Virus type I (HTLV-I)
Асоційований з 2 фатальними хворобами

Слайд 68HTLV-I та ATL
1980 Галло ізолював тип C retrovirus (HTLV1) від хворого
HTLV-I та ATL
1980 Галло ізолював тип C retrovirus (HTLV1) від хворого

Слайд 69Міжклітинний контакт
Статевий (60% від чоловіка до жінки проти 1% від жінки до
Міжклітинний контакт
Статевий (60% від чоловіка до жінки проти 1% від жінки до

Слайд 70Онкогенез, опосередкований важливим ретровірусним білком
Виключення з парадігми ретровірусного онкогенезу (HTLV-1)
HTLV-1 не
Онкогенез, опосередкований важливим ретровірусним білком
Виключення з парадігми ретровірусного онкогенезу (HTLV-1)
HTLV-1 не

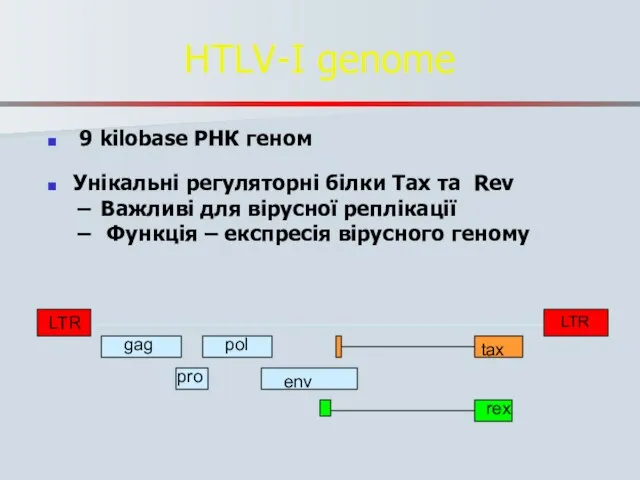
Слайд 71HTLV-I genome
9 kilobase РНК геном
Унікальні регуляторні білки Tax та Rev
Важливі
HTLV-I genome
9 kilobase РНК геном
Унікальні регуляторні білки Tax та Rev
Важливі
Слайд 72Tax тa Oнкогенез
Tax необхідний для вірусної реплікації
40kda фосфопротеїн
Tранскрипційний активатор геному HTLV-I
Взаємодія з
Tax тa Oнкогенез
Tax необхідний для вірусної реплікації
40kda фосфопротеїн
Tранскрипційний активатор геному HTLV-I
Взаємодія з

Слайд 73Tax – множинний трансактиватор
Зв”язування з клітинними транскрипційними факторами посилює їх взаємодію з
Tax – множинний трансактиватор
Зв”язування з клітинними транскрипційними факторами посилює їх взаємодію з

Слайд 74Білки регулятори клітинного циклу – мішені дії Tax
Iнактивація p53 (G1/S контроль рестрикції)
Білки регулятори клітинного циклу – мішені дії Tax
Iнактивація p53 (G1/S контроль рестрикції)

Слайд 75Tax репресує DНК -репарацію
Репресія DНК pol β , яка залучена в репарацію
Tax репресує DНК -репарацію
Репресія DНК pol β , яка залучена в репарацію
Слайд 76Механізми клітинної трансформації ретровірусами
Механізми клітинної трансформації ретровірусами

Слайд 77Онкогенні
РНК- віруси
Онкогенні
РНК- віруси
 Гестационнный сахарный диабет
Гестационнный сахарный диабет Будова Атома
Будова Атома  Презентация на тему Алтай
Презентация на тему Алтай Понятие и формы реализации права
Понятие и формы реализации права Самоуправление
Самоуправление Наш Пушкин
Наш Пушкин Обзор программ CCNA v.4.0
Обзор программ CCNA v.4.0 Переборные алгоритмы
Переборные алгоритмы Конструирование из строительного материала в младшей группе
Конструирование из строительного материала в младшей группе  Фонари
Фонари Омонимы и омографы
Омонимы и омографы Соломинки. Делаем своими руками
Соломинки. Делаем своими руками Инструкция по оформлению витрин ППО рекламной продукцией A1
Инструкция по оформлению витрин ППО рекламной продукцией A1 Выбирай банк смолоду
Выбирай банк смолоду Расчет электрических цепей
Расчет электрических цепей РОЗНИЧНАЯ ТОРГОВЛЯ
РОЗНИЧНАЯ ТОРГОВЛЯ мужчины и женщины - такие похожие и такие разные?
мужчины и женщины - такие похожие и такие разные? Игрушки и сувениры из Сергиева Посада и англоязычных стран
Игрушки и сувениры из Сергиева Посада и англоязычных стран Потому что знает:выйти из нее можно только одним способом.И нормальному человеку он не годится.
Потому что знает:выйти из нее можно только одним способом.И нормальному человеку он не годится. Выдающиеся деятели культуры. Генри Мур (1898-1986)
Выдающиеся деятели культуры. Генри Мур (1898-1986) И.Бунин Современная политическая ситуация в России: основные проблемы
И.Бунин Современная политическая ситуация в России: основные проблемы Видеопрезентация рисунков учеников 2а класса
Видеопрезентация рисунков учеников 2а класса Субъекты и участники обязательного медицинского страхования: понятие и различие правового статуса
Субъекты и участники обязательного медицинского страхования: понятие и различие правового статуса ЧТО ИЗУЧАЮТ СИНТАКСИС И ПУНКТУАЦИЯ
ЧТО ИЗУЧАЮТ СИНТАКСИС И ПУНКТУАЦИЯ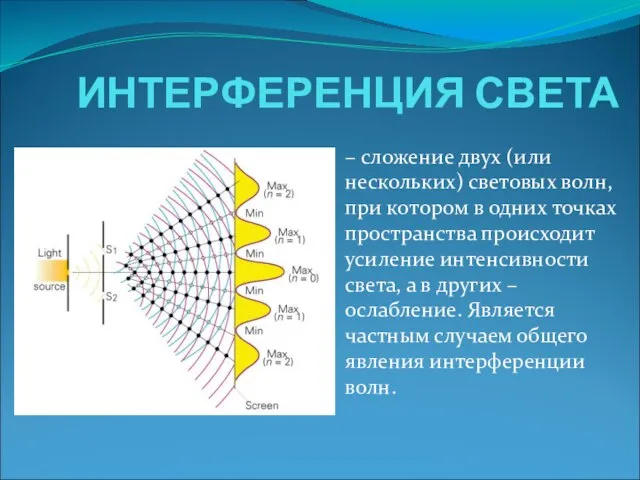 Интерференция света
Интерференция света  Единый государственный экзамен: проблемы содержания и организации
Единый государственный экзамен: проблемы содержания и организации Теория организации. Классические виды организационных структур. (Тема 2)
Теория организации. Классические виды организационных структур. (Тема 2) КООРДИНАЦИЯ МЕЖДУНАРОДНОГО СОТРУДНИЧЕСТВА МЕЖДУ РОССИЙСКОЙ ТП И ЕВРОПЕЙСКИМИ СТИ В ОБЛАСТИ АЭРОНАВТИКИ
КООРДИНАЦИЯ МЕЖДУНАРОДНОГО СОТРУДНИЧЕСТВА МЕЖДУ РОССИЙСКОЙ ТП И ЕВРОПЕЙСКИМИ СТИ В ОБЛАСТИ АЭРОНАВТИКИ