- Полвека с Центральной догмой молекулярной биологии

Содержание
- 2. Доклад на Международной научной конференции «Чарльз Дарвин и современная биология» 21-23 сентября 2009 г., Санкт-Петербург
- 3. Памяти Френсиса Крика и Джона Мейнарда Смита: Francis Harry Compton Crick June 8, 1916 - July
- 4. Википедия Центральная догма молекулярной биологии (ЦДМБ) — обобщающее наблюдаемое в природе правило (?) реализации генетической информации:
- 5. Wikipedia. Поток инфомации в биологических системах
- 6. «Общепринятая» формулировка ЦДМБ «Центральная догма в ее общепринятой форме описывает матричные процессы: репликацию, транскрипцию, трансляцию, обладающие
- 7. Наиболее распространенные интерпретации Центральной догмы: ДНК делает РНК делает Белок ДНК делает РНК делает Белки делают
- 8. Google: Картинки по запросу central dogma of molecular biology
- 9. Google: Картинки по запросу central dogma of molecular biology
- 10. Google: Видео по запросу central dogma of molecular biology
- 11. Идиосинкразия Центральная догма молекулярной биологии (ЦДМБ) почему-то вызывает у некоторых авторов желание низвергнуть ее или, как
- 12. Долой! «Смерть Центральной догмы» «Центральная догма – миф» «Центральная догма неверна» «Центральная догма ошибочна» «Центральная догма
- 13. Thieffry D., Sarkar S. Forty years under the central dogma. Trends in biochemical sciences, 1998, vol.
- 14. Вред или бред? Есть, например, мнение, что ЦДМБ «принесла биологии больше вреда, чем лысенковщина» (Чайковский, 2006;
- 15. SYMPOSIA OF THE SOCIETY FOR EXPERIMENTAL BIOLOGY NUMBER XII THE BIOLOGICAL REPLICATION OF MACROMOLECULES CAMBRIDGE: AT
- 16. Crick F. H. C. (1958) On protein synthesis. Symp. Soc. Exp. Biol. 12: 138-163 Несколько лет
- 17. В чем проблема? Как могут синтезироваться белки? Молекулы белка характеризуются исключительной специфичностью, которая несомненно зависит от
- 18. Существо проблемы Химическая реакция, с помощью которой любые две аминокислоты (или активированные аминокислоты) соединяются вместе, скорее
- 19. Общие принципы Мои собственные размышления (и размышления многих моих коллег) основаны на двух общих принципах, которые
- 20. Центральная Догма Как только «информация» переходит в белок, она не может выйти обратно (появиться снова). Более
- 21. Crick F. H. C. (1970) Central Dogma of Molecular Biology. Nature 227, 561-563 Abstract: Центральная догма
- 22. Crick F. H. C. (1970) Центральная догма ничего не говорит о том, из чего сделана машинерия
- 23. Crick F. H. C. (1970) Совершенно немыслимо, исходя их стереохимических соображений, что перенос белок → белок
- 24. Сейчас, когда мы достаточно осведомлены о невероятной сложности внутриклеточных белок-синтезирующих фабрик, то невозможность ни репликации, ни
- 25. Итак Я предположил, что «информация», попав в белок, уже не может передаться оттуда куда-то еще», добавив,
- 26. Открытия, интерпретируемые как исключения из ЦДМБ Обратная транскрипция Рибозимы Редактирование РНК Сплайсинг Эпигенетические явления (Геномный импринтинг)
- 27. В чем же дело? Почему же ЦДМБ вызывавет такое недопонимание и неприятие? Одной из причин подобных
- 28. Какая информация передается от белков к НК? Сейчас известно, что, редупликация ДНК практически невозможна без ДНК-полимеразы.
- 29. Правило принуждения в молекулярной биологии В живых (биологических) системах на молекулярном уровне очень часто происходят процессы,
- 30. Правило принуждения в молекулярной биологии Как правило, природные молекулярно-биологические процессы осуществляются по принуждению. В этом состоит
- 31. Maynard Smith J. (1989) Evolutionary Genetics. Oxford University press Неправда, что ДНК может реплицироваться без белков,
- 32. Lewontin Richard C. (2003), The DNA Era GeneWatch 16(4) http://www.gene-watch.org/pages/genewatch.html ДНК не является самореплицирующейся субстанцией, подобно
- 33. Молекулярная машина Drexler K. Eric. (1999) Building Molecular Machine Systems Trends Biotechnol. 17(1), 5-7 http://www.imm.org/Reports/index.html Выражение
- 34. Schneider T. D. (1991). Theory of molecular machines. I. Channel capacity of molecular machines. J. Theor.
- 36. Аристотель. О душе Дабы не быть тавтологией, «Определение [предмета] должно вскрывать не только то, что [он]
- 37. «Причинная» формулировка ЦДМБ For any protein sequence its replication, transcription and (reverse) translation are prohibited because
- 38. Операционализм Научные понятия должны быть определены в терминах выполняемых операций. Например, понятие длины тогда определено, когда
- 39. Операциональная формулировка ЦДМБ Если ввести в клетку белок нового типа, то это не приведет к синтезу
- 40. Если в клетке присутствует белок с новой аминокислотной последовательностью, то это не может вызвать образование молекулы
- 41. Чужеродный белок Назовем белок чужеродным для данной клетки, если в ней отсутствует ген для этого белка,
- 42. Постулат Крика (ЦДМБ) – молекулярный аналог постулата Вейсмана По существу ЦДМБ есть молекулярный аналог постулата Вейсмана
- 43. Крик Ф. Безумный поиск: Личный взгляд на научное открытие. – Москва-Ижевск: Ин-т компьютерных исследований, 2004. –
- 44. Crick F. H. C. (1970) Центральная догма сегодня столько же важна как и тогда, когда она
- 45. Maynard Smith J. (1975-2000) В настоящее время нет оснований сомневаться в справедливости центральной догмы. Центральная догма
- 46. Значение ЦДМБ В 60-е годы ЦДМБ помогла отсечь из рассмотрения неправдоподобные процессы типа обратной трансляции белков
- 47. Запрет Крика Центральная догма – столь же важный запрет, как принцип (запрет) Паули, как запреты на
- 48. Треугольник Крика Центральная догма молекулярной биологии есть вооборажаемо домысливаемое ОТСУТСТВИЕ стрелок на «треугольнике Крика» (слева). Нагляднее
- 49. Crick F. Ideas on Protein Synthesis (Oct. 1956) http://profiles.nlm.nih.gov/SC/B/F/T/_/scbbft.pdf
- 50. Смысл стрелок на треугольнике Крика Стрелки указывают не столько направление, сколько саму возможность копирования, переписывания (целиком
- 51. Homo faber Только Человек способен обойти запреты Центральной догмы и осуществить «репликацию», «транскрипцию» и «обратную трансляцию»
- 52. Инге-Вечтомов С.Г., Миронова Л.Н. (2003) Бешеные коровы, дрожжи и "белковая наследственность" Наука в России № 3
- 53. Треугольник Инге-Вечтомова Увы, здесь смысл стрелок для «переноса информации» от Белка к Белку совершенно иной, нежели
- 54. Прионы Прионы – это взбесившиеся белки, которые берутся за руки, чтоб не пропасть поодиночке Прионы –
- 55. Лобашев Михаил Ефимович 11 ноября1907 – 4 января1971
- 57. Сигнальная наследственность — передача поведенческих навыков потомству посредством условного рефлекса подражания. По Павлову – это вторая
- 58. Контакт и подражание или заразительность Это интереснейшее явление происходит в результате так называемого биологического контакта поколений,
- 59. Is Lamarck back? (Ю.О. Чернов, 2001) В динамике сигнальной наследственности, как и в прионизации, много общего
- 60. Сравнение (аналогия) не есть доказательство Прионизация есть прионизация. Это уникальное, не имеющее аналогов явление, о чем
- 61. Аристотель Признак образованного человека: при любом типе исследования добиваться такой степени точности, какую только дозволяет предмет
- 62. Андрей Николаевич Колмогоров Писать математическую статью надо так, будто находишься под гильотиной, и одно неточное слово
- 63. Итак 1. Центральная догма молекулярной биологии (ЦДМБ) есть запрет на репликацию, транскрипцию и трансляцию белков (запрет
- 64. 5. ЦДМБ – отнюдь не то же самое, что гипотеза последовательности, с которой ее часто путают
- 65. Ученый должен быть скептиком 9. Распространенное или общепринятое не означает правильное. Подвергайте сомнению интепретации. Штудируйте первоисточники.
- 66. ПРИЛОЖЕНИЕ Цитаты на языке оригинала Дополнительные иллюстрации Литература
- 67. General principles My own thinking (and that of many my colleagues) is based on two general
- 68. The Central Dogma This states that once ‘information’ has passed into protein it cannot get out
- 69. Crick F. H. C. (1970) Central Dogma of Molecular Biology. Nature 227, 561-563 Abstract: The central
- 70. Crick F. H. C. (1970) Central dogma says nothing about what the machinery of transfer is
- 71. Crick F. H. C. (1970) It was most unlikely, for stereochemical reasons, that protein → protein
- 72. John Maynard Smith If a new kind of protein is introduced into a cell, this cannot
- 73. Термины или метафоры? Кибернетические и технические выражения типа «информация» и «матрица» являются (по крайней мере в
- 74. «Информация», «код», «матрица» суть метафоры Метафоры повсеместны в науке. Но, когда они служат лишь суррогатами для
- 75. Информация Информация есть информация, не вещество и не энергия. Норберт Винер, Кибернетика, 1962. Приходится сожалеть, что
- 76. Stephen William Hawking 8 Jan 1942
- 77. Черные дыры и «Информационный парадокс»
- 78. Полисемия понятия «информация» Информация как сообщение Информация как коммуникация Информация как представление Информация как знание Информация
- 79. Три типа информации в молекулярной биологии Необходимо различать по меньшей мере три типа молекулярно-биологической информации (или
- 80. Какие еще обсуждаются типы информации в молекулярной биологии Структурная Функциональная Связанная (bond information) Свободная (Ryan J.
- 81. Lionel Sharples Penrose (1898–1972) Genetics, Vol. 150, 1333-1340, December 1998
- 83. Музыкальный палиндром
- 84. Sir Roger Penrose 8 Aug 1931
- 85. A Self-reproducing Analogue L. S. Penrose & R. Penrose Nature 4571:1183, 1957 Исходное расположение A.B.B.A.B.A.A.A.B.A.B.B. После
- 86. L. S. Penrose, "Self-Reproducing Machines", Scientific American, 200:105-114 (June 1959). В нашем воображении процесс механического самовоспроизведения
- 87. Есть ген – есть белок
- 88. Есть мутация – есть мутант
- 89. Есть модификация – нет мутанта
- 90. Нет гена – нет белка
- 91. Прион внутри
- 92. Прион извне
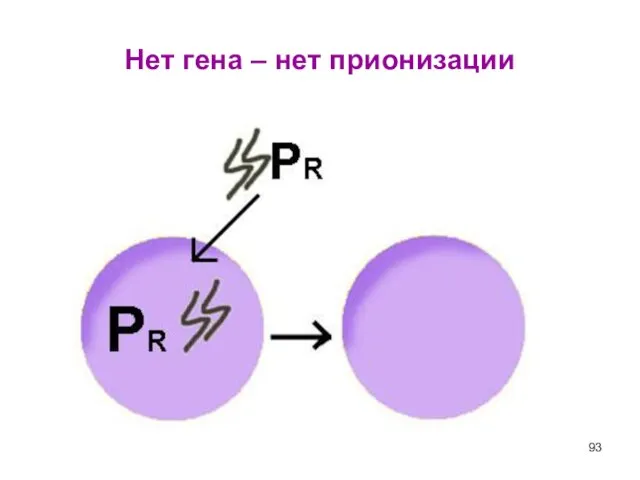
- 93. Нет гена – нет прионизации
- 94. Литература: информация Ryan J. P. F. J. (1972) Information, entropy and various systems. J. Theor. Biol.
- 95. Литература: молекулярные машины Schneider T. D. (1991). Theory of molecular machines. I. Channel capacity of molecular
- 96. Литература: Maynard Smith Maynard Smith J. (1969) The status of neo-Darwinism. In: Towards a Theoretical Biology,
- 97. Литература: прионы и ЦДМБ Keyes, M.E. (1999) The Prion Challenge to the ‘Central Dogma’ of Molecular
- 98. Литература: прионы и ЦДМБ Muzumdar M. (2002) Prions: Proteinaceous Predators. Harvard Science Review 37: 37-39. Инге-Вечтомов
- 99. Признательность Я глубоко признателен Юрию Олеговичу Чернову и Сергею Георгиевичу Инге-Вечтомову, редкие минуты общения с которыми
- 101. Скачать презентацию
Слайд 2Доклад на
Международной научной конференции
«Чарльз Дарвин и современная биология»
21-23 сентября 2009 г.,
Доклад на
Международной научной конференции
«Чарльз Дарвин и современная биология»
21-23 сентября 2009 г.,

Слайд 3Памяти
Френсиса Крика и Джона Мейнарда Смита:
Francis Harry Compton Crick
June 8,
Памяти
Френсиса Крика и Джона Мейнарда Смита:
Francis Harry Compton Crick June 8,

Слайд 4Википедия
Центральная догма молекулярной биологии (ЦДМБ) —
обобщающее наблюдаемое в природе правило (?) реализации
Википедия
Центральная догма молекулярной биологии (ЦДМБ) —
обобщающее наблюдаемое в природе правило (?) реализации

Слайд 5Wikipedia. Поток инфомации в биологических системах
Wikipedia. Поток инфомации в биологических системах

Слайд 6«Общепринятая» формулировка ЦДМБ
«Центральная догма в ее общепринятой форме описывает матричные процессы: репликацию,
«Общепринятая» формулировка ЦДМБ
«Центральная догма в ее общепринятой форме описывает матричные процессы: репликацию,

Слайд 7Наиболее распространенные интерпретации Центральной догмы:
ДНК делает РНК делает Белок
ДНК делает РНК делает
Наиболее распространенные интерпретации Центральной догмы:
ДНК делает РНК делает Белок
ДНК делает РНК делает

Слайд 8Google: Картинки по запросу central dogma of molecular biology
Google: Картинки по запросу central dogma of molecular biology

Слайд 9Google: Картинки по запросу central dogma of molecular biology
Google: Картинки по запросу central dogma of molecular biology

Слайд 10Google: Видео по запросу central dogma of molecular biology
Google: Видео по запросу central dogma of molecular biology
Слайд 11Идиосинкразия
Центральная догма молекулярной биологии (ЦДМБ)
почему-то вызывает у некоторых авторов желание низвергнуть
Идиосинкразия
Центральная догма молекулярной биологии (ЦДМБ)
почему-то вызывает у некоторых авторов желание низвергнуть

Слайд 12Долой!
«Смерть Центральной догмы»
«Центральная догма – миф»
«Центральная догма неверна»
«Центральная догма ошибочна»
«Центральная догма
Долой!
«Смерть Центральной догмы»
«Центральная догма – миф»
«Центральная догма неверна»
«Центральная догма ошибочна»
«Центральная догма

Слайд 13Thieffry D., Sarkar S.
Forty years under the central dogma.
Trends in biochemical sciences,
Thieffry D., Sarkar S. Forty years under the central dogma. Trends in biochemical sciences,

Слайд 14Вред или бред?
Есть, например, мнение, что ЦДМБ «принесла биологии больше вреда, чем
Вред или бред?
Есть, например, мнение, что ЦДМБ «принесла биологии больше вреда, чем

Слайд 15SYMPOSIA OF THE
SOCIETY FOR EXPERIMENTAL BIOLOGY
NUMBER XII
THE
BIOLOGICAL REPLICATION
OF MACROMOLECULES
CAMBRIDGE: AT THE
SYMPOSIA OF THE
SOCIETY FOR EXPERIMENTAL BIOLOGY
NUMBER XII
THE
BIOLOGICAL REPLICATION
OF MACROMOLECULES
CAMBRIDGE: AT THE

Слайд 16Crick F. H. C. (1958)
On protein synthesis.
Symp. Soc. Exp. Biol.
Crick F. H. C. (1958) On protein synthesis. Symp. Soc. Exp. Biol.

Слайд 17В чем проблема?
Как могут синтезироваться белки?
Молекулы белка характеризуются исключительной специфичностью, которая несомненно
В чем проблема?
Как могут синтезироваться белки?
Молекулы белка характеризуются исключительной специфичностью, которая несомненно

Слайд 18Существо проблемы
Химическая реакция, с помощью которой любые две аминокислоты (или активированные аминокислоты)
Существо проблемы
Химическая реакция, с помощью которой любые две аминокислоты (или активированные аминокислоты)

Слайд 19 Общие принципы
Мои собственные размышления
(и размышления многих моих коллег)
основаны на
Общие принципы
Мои собственные размышления
(и размышления многих моих коллег)
основаны на

Слайд 20Центральная Догма
Как только «информация» переходит в белок, она не может выйти обратно
Центральная Догма
Как только «информация» переходит в белок, она не может выйти обратно

Слайд 21Crick F. H. C. (1970) Central Dogma of Molecular Biology. Nature 227,
Crick F. H. C. (1970) Central Dogma of Molecular Biology. Nature 227,

Слайд 22Crick F. H. C. (1970)
Центральная догма ничего не говорит о том, из
Crick F. H. C. (1970)
Центральная догма ничего не говорит о том, из

Слайд 23Crick F. H. C. (1970)
Совершенно немыслимо, исходя их стереохимических соображений, что перенос
Crick F. H. C. (1970)
Совершенно немыслимо, исходя их стереохимических соображений, что перенос

Слайд 24Сейчас, когда мы достаточно осведомлены о невероятной сложности внутриклеточных белок-синтезирующих фабрик,
то
Сейчас, когда мы достаточно осведомлены о невероятной сложности внутриклеточных белок-синтезирующих фабрик,
то

Слайд 25Итак
Я предположил, что «информация», попав в белок, уже не может передаться оттуда
Итак
Я предположил, что «информация», попав в белок, уже не может передаться оттуда

Слайд 26Открытия, интерпретируемые как исключения из ЦДМБ
Обратная транскрипция
Рибозимы
Редактирование РНК
Сплайсинг
Эпигенетические явления (Геномный импринтинг)
РНК-интерференция
Прионизация
Открытия, интерпретируемые как исключения из ЦДМБ
Обратная транскрипция
Рибозимы
Редактирование РНК
Сплайсинг
Эпигенетические явления (Геномный импринтинг)
РНК-интерференция
Прионизация

Слайд 27В чем же дело?
Почему же ЦДМБ вызывавет такое недопонимание и неприятие?
Одной из
В чем же дело?
Почему же ЦДМБ вызывавет такое недопонимание и неприятие?
Одной из

Слайд 28Какая информация передается от белков к НК?
Сейчас известно, что, редупликация ДНК практически
Какая информация передается от белков к НК?
Сейчас известно, что, редупликация ДНК практически

Слайд 29Правило принуждения в молекулярной биологии
В живых (биологических) системах на молекулярном уровне очень
Правило принуждения в молекулярной биологии
В живых (биологических) системах на молекулярном уровне очень

Слайд 30Правило принуждения
в молекулярной биологии
Как правило, природные молекулярно-биологические процессы осуществляются по принуждению.
Правило принуждения
в молекулярной биологии
Как правило, природные молекулярно-биологические процессы осуществляются по принуждению.

Слайд 31Maynard Smith J. (1989) Evolutionary Genetics. Oxford University press
Неправда, что ДНК может
Maynard Smith J. (1989) Evolutionary Genetics. Oxford University press
Неправда, что ДНК может

Слайд 32Lewontin Richard C. (2003), The DNA Era
GeneWatch 16(4)
http://www.gene-watch.org/pages/genewatch.html
ДНК не является
Lewontin Richard C. (2003), The DNA Era
GeneWatch 16(4)
http://www.gene-watch.org/pages/genewatch.html
ДНК не является

Слайд 33Молекулярная машина
Drexler K. Eric. (1999) Building Molecular Machine Systems
Trends Biotechnol. 17(1),
Молекулярная машина Drexler K. Eric. (1999) Building Molecular Machine Systems Trends Biotechnol. 17(1),

Слайд 34Schneider T. D. (1991). Theory of molecular machines. I. Channel capacity of
Schneider T. D. (1991). Theory of molecular machines. I. Channel capacity of

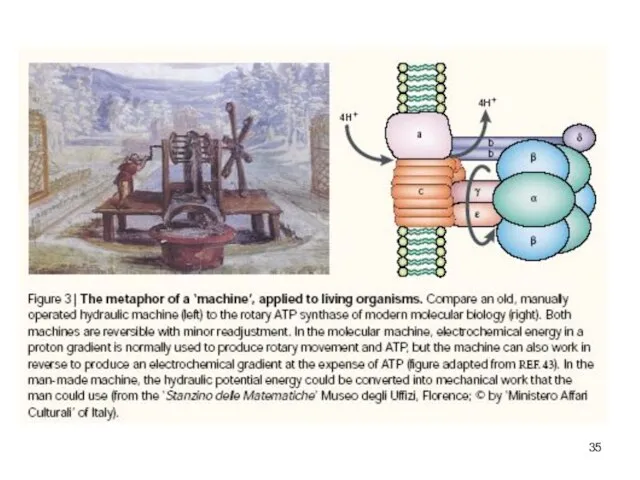
Слайд 36Аристотель. О душе
Дабы не быть тавтологией,
«Определение [предмета] должно вскрывать не только то,
Аристотель. О душе
Дабы не быть тавтологией,
«Определение [предмета] должно вскрывать не только то,
![Аристотель. О душе Дабы не быть тавтологией, «Определение [предмета] должно вскрывать не](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/471236/slide-35.jpg)
Слайд 37«Причинная» формулировка ЦДМБ
For any protein sequence its replication, transcription and (reverse) translation
«Причинная» формулировка ЦДМБ
For any protein sequence its replication, transcription and (reverse) translation

Слайд 38Операционализм
Научные понятия должны быть определены в терминах выполняемых операций.
Например, понятие длины тогда
Операционализм
Научные понятия должны быть определены в терминах выполняемых операций.
Например, понятие длины тогда

Слайд 39Операциональная формулировка ЦДМБ
Если ввести в клетку белок нового типа, то это не
Операциональная формулировка ЦДМБ
Если ввести в клетку белок нового типа, то это не

Слайд 40Если в клетке присутствует белок с новой аминокислотной последовательностью, то это не
Если в клетке присутствует белок с новой аминокислотной последовательностью, то это не

Слайд 41Чужеродный белок
Назовем белок чужеродным для данной клетки, если в ней отсутствует ген
Чужеродный белок
Назовем белок чужеродным для данной клетки, если в ней отсутствует ген

Слайд 42Постулат Крика (ЦДМБ) –
молекулярный аналог постулата Вейсмана
По существу ЦДМБ есть молекулярный
Постулат Крика (ЦДМБ) –
молекулярный аналог постулата Вейсмана
По существу ЦДМБ есть молекулярный

Слайд 43Крик Ф. Безумный поиск: Личный взгляд на научное открытие. – Москва-Ижевск: Ин-т
Крик Ф. Безумный поиск: Личный взгляд на научное открытие. – Москва-Ижевск: Ин-т

Слайд 44Crick F. H. C. (1970)
Центральная догма сегодня столько же важна как и
Crick F. H. C. (1970)
Центральная догма сегодня столько же важна как и

Слайд 45Maynard Smith J. (1975-2000)
В настоящее время нет оснований сомневаться в справедливости центральной
Maynard Smith J. (1975-2000)
В настоящее время нет оснований сомневаться в справедливости центральной

Слайд 46Значение ЦДМБ
В 60-е годы ЦДМБ помогла отсечь из рассмотрения неправдоподобные процессы типа
Значение ЦДМБ
В 60-е годы ЦДМБ помогла отсечь из рассмотрения неправдоподобные процессы типа

Слайд 47Запрет Крика
Центральная догма – столь же важный запрет,
как принцип (запрет) Паули,
как
Запрет Крика
Центральная догма – столь же важный запрет,
как принцип (запрет) Паули,
как

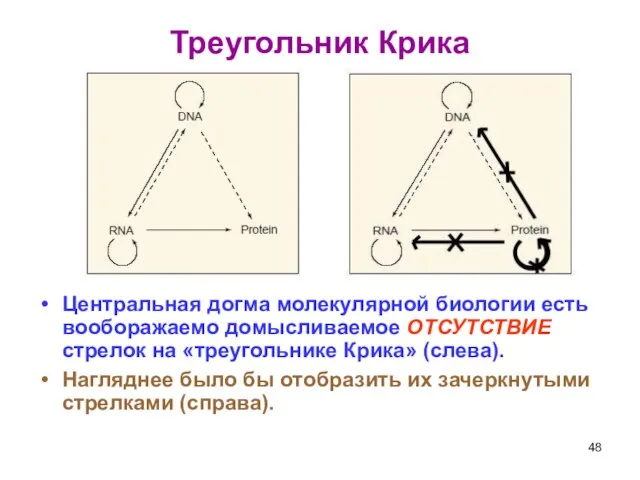
Слайд 48Треугольник Крика
Центральная догма молекулярной биологии есть вооборажаемо домысливаемое ОТСУТСТВИЕ стрелок на «треугольнике
Треугольник Крика
Центральная догма молекулярной биологии есть вооборажаемо домысливаемое ОТСУТСТВИЕ стрелок на «треугольнике
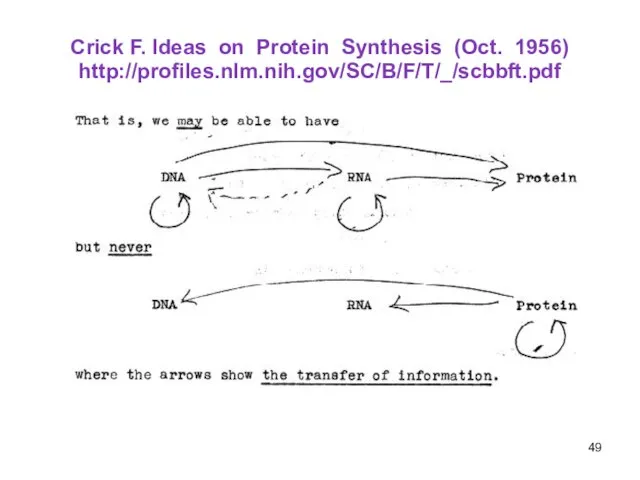
Слайд 49Crick F. Ideas on Protein Synthesis (Oct. 1956)
http://profiles.nlm.nih.gov/SC/B/F/T/_/scbbft.pdf
Crick F. Ideas on Protein Synthesis (Oct. 1956)
http://profiles.nlm.nih.gov/SC/B/F/T/_/scbbft.pdf
Слайд 50Смысл стрелок на треугольнике Крика
Стрелки указывают не столько направление, сколько саму возможность
Смысл стрелок на треугольнике Крика
Стрелки указывают не столько направление, сколько саму возможность

Слайд 51Homo faber
Только Человек способен обойти запреты Центральной догмы и осуществить «репликацию», «транскрипцию»
Homo faber
Только Человек способен обойти запреты Центральной догмы и осуществить «репликацию», «транскрипцию»

Слайд 52Инге-Вечтомов С.Г., Миронова Л.Н. (2003) Бешеные коровы, дрожжи и "белковая наследственность" Наука
Инге-Вечтомов С.Г., Миронова Л.Н. (2003) Бешеные коровы, дрожжи и "белковая наследственность" Наука
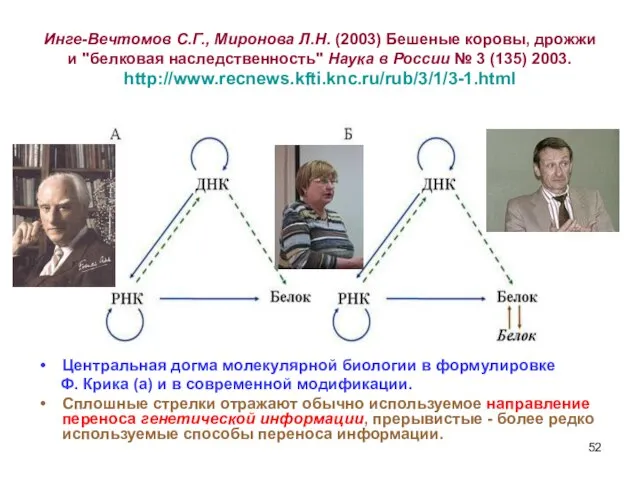
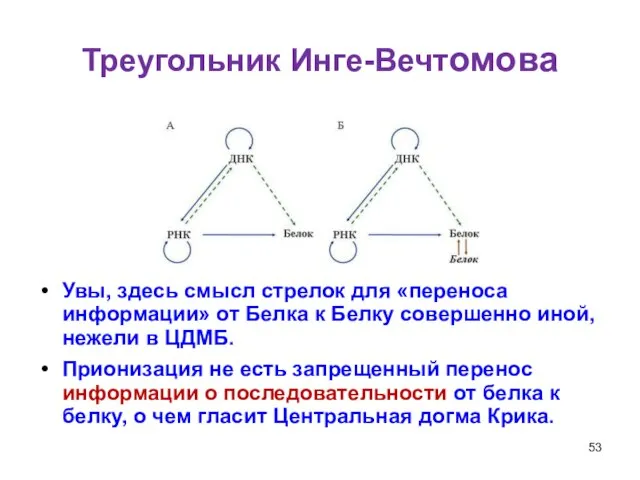
Слайд 53Треугольник Инге-Вечтомова
Увы, здесь смысл стрелок для «переноса информации» от Белка к Белку
Треугольник Инге-Вечтомова
Увы, здесь смысл стрелок для «переноса информации» от Белка к Белку
Слайд 54Прионы
Прионы – это взбесившиеся белки, которые берутся за руки, чтоб не пропасть
Прионы
Прионы – это взбесившиеся белки, которые берутся за руки, чтоб не пропасть

Слайд 55Лобашев Михаил Ефимович
11 ноября1907 – 4 января1971
Лобашев Михаил Ефимович
11 ноября1907 – 4 января1971


Слайд 57Сигнальная наследственность
— передача поведенческих навыков потомству посредством условного рефлекса подражания.
По Павлову
Сигнальная наследственность
— передача поведенческих навыков потомству посредством условного рефлекса подражания.
По Павлову

Слайд 58Контакт и подражание или заразительность
Это интереснейшее явление происходит в результате так называемого
Контакт и подражание или заразительность
Это интереснейшее явление происходит в результате так называемого

Слайд 59Is Lamarck back? (Ю.О. Чернов, 2001)
В динамике сигнальной наследственности, как и в
Is Lamarck back? (Ю.О. Чернов, 2001)
В динамике сигнальной наследственности, как и в

Слайд 60Сравнение (аналогия) не есть доказательство
Прионизация есть прионизация.
Это уникальное, не имеющее аналогов явление,
Сравнение (аналогия) не есть доказательство
Прионизация есть прионизация.
Это уникальное, не имеющее аналогов явление,

Слайд 61Аристотель
Признак образованного человека: при любом типе исследования добиваться такой степени точности, какую
Аристотель
Признак образованного человека: при любом типе исследования добиваться такой степени точности, какую

Слайд 62Андрей Николаевич Колмогоров
Писать математическую статью надо так, будто находишься под гильотиной, и
Андрей Николаевич Колмогоров
Писать математическую статью надо так, будто находишься под гильотиной, и

Слайд 63Итак
1. Центральная догма молекулярной биологии (ЦДМБ) есть запрет на репликацию, транскрипцию и
Итак
1. Центральная догма молекулярной биологии (ЦДМБ) есть запрет на репликацию, транскрипцию и

Слайд 645. ЦДМБ – отнюдь не то же самое, что гипотеза последовательности, с
5. ЦДМБ – отнюдь не то же самое, что гипотеза последовательности, с

Слайд 65Ученый должен быть скептиком
9. Распространенное или общепринятое не означает правильное.
Подвергайте сомнению интепретации.
Штудируйте
Ученый должен быть скептиком
9. Распространенное или общепринятое не означает правильное.
Подвергайте сомнению интепретации.
Штудируйте

Слайд 66ПРИЛОЖЕНИЕ
Цитаты на языке оригинала
Дополнительные иллюстрации
Литература
ПРИЛОЖЕНИЕ
Цитаты на языке оригинала
Дополнительные иллюстрации
Литература

Слайд 67General principles
My own thinking (and that of many my colleagues) is based
General principles
My own thinking (and that of many my colleagues) is based

Слайд 68The Central Dogma
This states that once ‘information’ has passed into protein it
The Central Dogma
This states that once ‘information’ has passed into protein it

Слайд 69Crick F. H. C. (1970) Central Dogma of Molecular Biology. Nature 227,
Crick F. H. C. (1970) Central Dogma of Molecular Biology. Nature 227,

Слайд 70Crick F. H. C. (1970)
Central dogma says nothing about what the machinery
Crick F. H. C. (1970)
Central dogma says nothing about what the machinery

Слайд 71Crick F. H. C. (1970)
It was most unlikely, for stereochemical reasons, that
Crick F. H. C. (1970)
It was most unlikely, for stereochemical reasons, that

Слайд 72John Maynard Smith
If a new kind of protein is introduced into a
John Maynard Smith
If a new kind of protein is introduced into a

Слайд 73Термины или метафоры?
Кибернетические и технические выражения типа «информация» и «матрица» являются (по
Термины или метафоры?
Кибернетические и технические выражения типа «информация» и «матрица» являются (по

Слайд 74 «Информация», «код», «матрица» суть метафоры
Метафоры повсеместны в науке. Но, когда они
«Информация», «код», «матрица» суть метафоры
Метафоры повсеместны в науке. Но, когда они

Слайд 75Информация
Информация есть информация, не вещество и не энергия.
Норберт Винер, Кибернетика,
Информация
Информация есть информация, не вещество и не энергия.
Норберт Винер, Кибернетика,

Слайд 76Stephen William Hawking
8 Jan 1942
Stephen William Hawking
8 Jan 1942

Слайд 77Черные дыры и «Информационный парадокс»
Черные дыры и «Информационный парадокс»

Слайд 78Полисемия понятия «информация»
Информация как сообщение
Информация как коммуникация
Информация как представление
Информация как знание
Информация
Полисемия понятия «информация»
Информация как сообщение
Информация как коммуникация
Информация как представление
Информация как знание
Информация

Слайд 79Три типа информации в молекулярной биологии
Необходимо различать по меньшей мере три типа
Три типа информации в молекулярной биологии
Необходимо различать по меньшей мере три типа

Слайд 80Какие еще обсуждаются типы информации в молекулярной биологии
Структурная
Функциональная
Связанная (bond information)
Свободная (Ryan J.
Какие еще обсуждаются типы информации в молекулярной биологии
Структурная
Функциональная
Связанная (bond information)
Свободная (Ryan J.

Слайд 81Lionel Sharples Penrose
(1898–1972)
Genetics, Vol. 150, 1333-1340, December 1998
Lionel Sharples Penrose
(1898–1972)
Genetics, Vol. 150, 1333-1340, December 1998


Слайд 83Музыкальный палиндром
Музыкальный палиндром

Слайд 84Sir Roger Penrose
8 Aug 1931
Sir Roger Penrose
8 Aug 1931

Слайд 85A Self-reproducing Analogue
L. S. Penrose & R. Penrose
Nature 4571:1183, 1957
Исходное расположение
A.B.B.A.B.A.A.A.B.A.B.B.
После добавления
A Self-reproducing Analogue
L. S. Penrose & R. Penrose
Nature 4571:1183, 1957
Исходное расположение
A.B.B.A.B.A.A.A.B.A.B.B.
После добавления

Слайд 86L. S. Penrose, "Self-Reproducing Machines", Scientific American, 200:105-114 (June 1959).
В нашем
L. S. Penrose, "Self-Reproducing Machines", Scientific American, 200:105-114 (June 1959).
В нашем

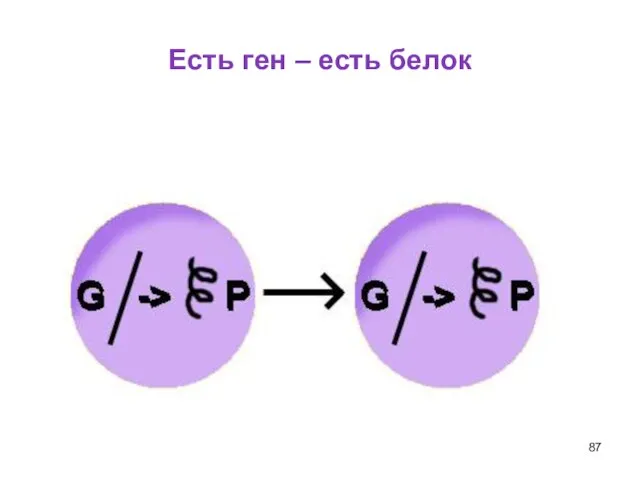
Слайд 87Есть ген – есть белок
Есть ген – есть белок
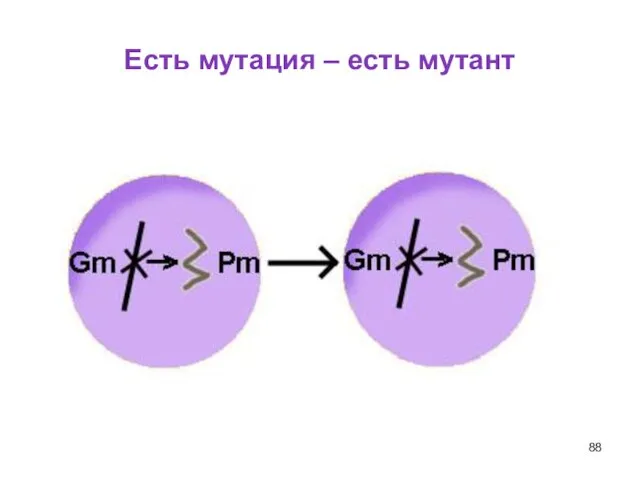
Слайд 88Есть мутация – есть мутант
Есть мутация – есть мутант
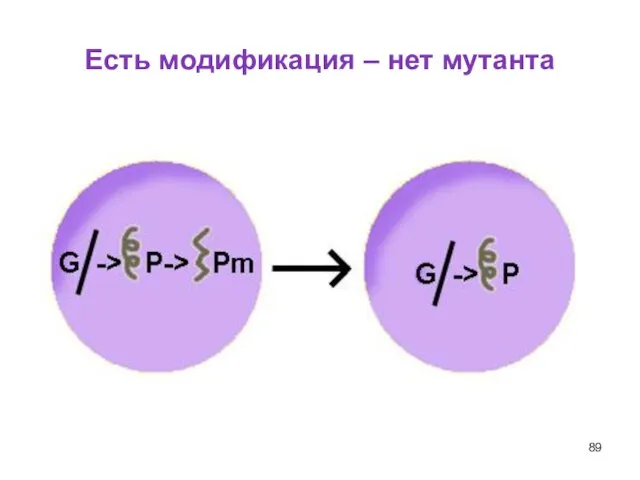
Слайд 89Есть модификация – нет мутанта
Есть модификация – нет мутанта
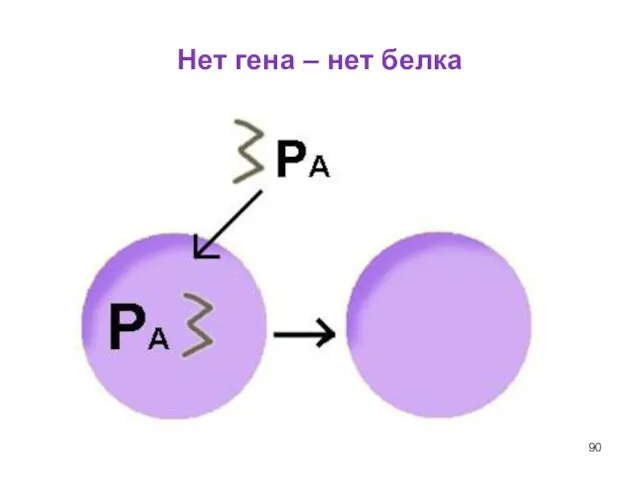
Слайд 90Нет гена – нет белка
Нет гена – нет белка
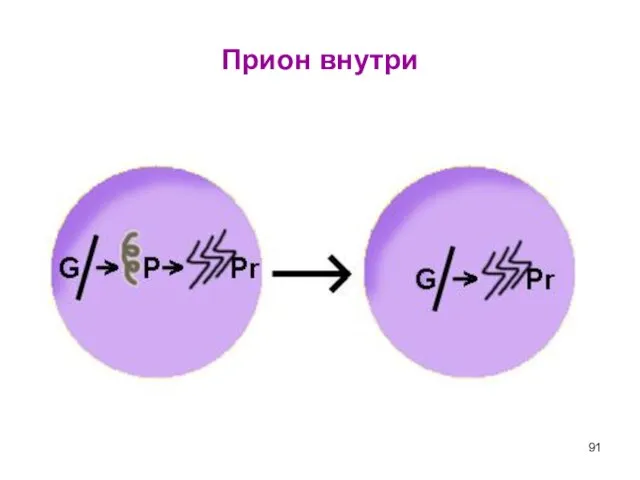
Слайд 91Прион внутри
Прион внутри
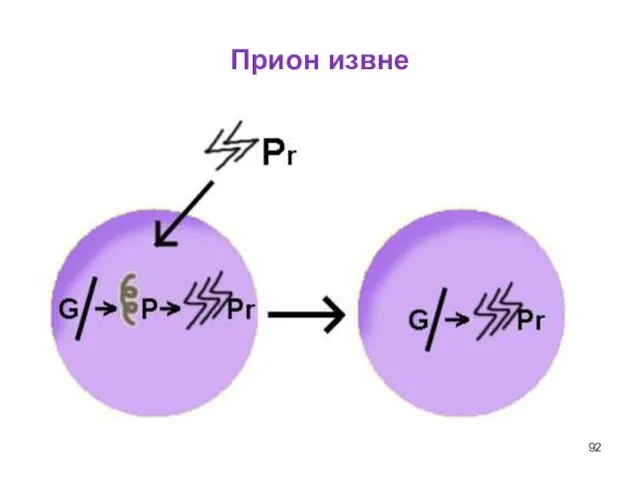
Слайд 92Прион извне
Прион извне
Слайд 93Нет гена – нет прионизации
Нет гена – нет прионизации
Слайд 94Литература: информация
Ryan J. P. F. J. (1972) Information, entropy and various systems.
Литература: информация
Ryan J. P. F. J. (1972) Information, entropy and various systems.

Слайд 95Литература: молекулярные машины
Schneider T. D. (1991). Theory of molecular machines. I. Channel
Литература: молекулярные машины
Schneider T. D. (1991). Theory of molecular machines. I. Channel

Слайд 96Литература: Maynard Smith
Maynard Smith J. (1969) The status of neo-Darwinism. In: Towards
Литература: Maynard Smith
Maynard Smith J. (1969) The status of neo-Darwinism. In: Towards

Слайд 97Литература: прионы и ЦДМБ
Keyes, M.E. (1999) The Prion Challenge to the ‘Central
Литература: прионы и ЦДМБ
Keyes, M.E. (1999) The Prion Challenge to the ‘Central

Слайд 98Литература: прионы и ЦДМБ
Muzumdar M. (2002) Prions: Proteinaceous Predators. Harvard Science Review
Литература: прионы и ЦДМБ
Muzumdar M. (2002) Prions: Proteinaceous Predators. Harvard Science Review

Слайд 99Признательность
Я глубоко признателен Юрию Олеговичу Чернову и Сергею Георгиевичу Инге-Вечтомову, редкие минуты
Признательность
Я глубоко признателен Юрию Олеговичу Чернову и Сергею Георгиевичу Инге-Вечтомову, редкие минуты

 Деревья в стихах
Деревья в стихах In a Soviet classroom
In a Soviet classroom  Presentation Title
Presentation Title  УНИВЕСАЛЬНОЕ УСТРОЙСТВО для работы с информацией
УНИВЕСАЛЬНОЕ УСТРОЙСТВО для работы с информацией  Презентация на тему Вычисление массы и объема тела (7 класс)
Презентация на тему Вычисление массы и объема тела (7 класс) Маркетинговые коммуникации в Интернете. Интернет-сайт как инструмент маркетинговых коммуникаций Ольга Галяткина, руководител
Маркетинговые коммуникации в Интернете. Интернет-сайт как инструмент маркетинговых коммуникаций Ольга Галяткина, руководител Урок русского языка и литературы
Урок русского языка и литературы Правила защиты от мошенников
Правила защиты от мошенников Викторина Правда или неправда? Шахматы
Викторина Правда или неправда? Шахматы 20140110_urok_3
20140110_urok_3 Гидродинамическое моделирование распределения запасов УВС по продуктивному пласту по результатам ГДИС
Гидродинамическое моделирование распределения запасов УВС по продуктивному пласту по результатам ГДИС Учиться нужно весело, чтоб хорошо учиться.
Учиться нужно весело, чтоб хорошо учиться. архитектура пк
архитектура пк Иоганн Вольфганг Фон Гете
Иоганн Вольфганг Фон Гете Германо - скандинавская мифология. Боги
Германо - скандинавская мифология. Боги Авторская педагогическая разработка адаптационная факультативный курс «Деловой английский для школьников» по УМК Дворецкой О.
Авторская педагогическая разработка адаптационная факультативный курс «Деловой английский для школьников» по УМК Дворецкой О. Компания INCO-Veritas занимается выпуском удобрений с 1976г. и в настоящее время является одним из крупнейших производителей данных товар
Компания INCO-Veritas занимается выпуском удобрений с 1976г. и в настоящее время является одним из крупнейших производителей данных товар Итоги квартала. Шаблон
Итоги квартала. Шаблон История создания гимна
История создания гимна Шкала электромагнитных излучений
Шкала электромагнитных излучений Пятнистый олень
Пятнистый олень Мордовская вышивка
Мордовская вышивка Управління якістю логістичних операцій на підприємстві
Управління якістю логістичних операцій на підприємстві Холе́рик. Что такое темперамент и какие есть виды?
Холе́рик. Что такое темперамент и какие есть виды? Игрушки царской семьи
Игрушки царской семьи ПРЕЗЕНТАЦИЯ ОСВ г Волжский 2022
ПРЕЗЕНТАЦИЯ ОСВ г Волжский 2022 Сергей Иванович Ожегов
Сергей Иванович Ожегов Элективный курсВОДА — САМОЕ УДИВИТЕЛЬНОЕ ВЕЩЕСТВОЗанятие по теме:ВОДА И ЧЕЛОВЕК
Элективный курсВОДА — САМОЕ УДИВИТЕЛЬНОЕ ВЕЩЕСТВОЗанятие по теме:ВОДА И ЧЕЛОВЕК