- Производство аминокислот

Содержание
- 2. Сферы практического применения препаратов аминокислот 1) в пищевой промышленности (около 30 % производимых аминокислот): Цистеин -
- 3. 3) в химической промышленности аминокислоты используют как предшественники : в производстве детергенов, в производстве полиаминокислот, в
- 4. Получение аминокислот гидролизом природного белоксодержащего сырья В качестве источников для гидролиза используют: отходы мясоперерабатывающей промышленности (отходы
- 5. Для выделения отдельных аминокислот из гидролизата проводят сложную многостадийную очистку, включающую в себя: нейтрализацию среды выделения,
- 6. Получение аминокислот химическим синтезом Химический синтез аминокислот занимает второе место по объему производства (около 30%). Основные
- 7. 2. Производство аминокислот связано с использованием дорогостоящего оборудования и агрессивных токсических соединений в качестве исходного сырья.
- 8. Микробиологический способ производства аминокислот Более 60 % всех производимых чистых препаратов аминокислот получают путем микробиологического синтеза.
- 9. При одноступенчатом синтезе в промышленных культиваторах выращивают ауксотрофные регуляторные мутанты, являющиеся сверхпродуцентами аминокислот. После завершения рабочего
- 10. В процессе двухступенчатого синтеза аминокислоты: получают предшественника аминокислоты (часто более дешевым химическим синтезом), с помощью ферментов,
- 11. Продуценты аминокислот. В качестве продуцентов аминокислот используют генетически измененные штаммы, обладающие способностью к сверхсинтезу аминокислот. Лучшими
- 12. Для производства аминокислот бактерии стали использовать с начала 50-х гг., при этом штаммы бактерий постоянно улучшали
- 13. Условия накопления глутамата: Полное или частичное подавление активности α-кетоглутаратдегидрогеназы, Добавление β-лактамных антибиотиков, Добавление ПАВ и жирных
- 14. Ауксотрофные мутанты применяются, когда необходимо синтезировать соединения являющиеся конечными продуктами разветвленных цепей метаболических реакций: Например: L-аспартат
- 15. Ауксотрофные мутанты способны накапливать конечные продукты неразветвленных путей биосинтеза. В таких случаях приходится отбирать мутанты с
- 16. Для увеличения выходааминокислот можно воспользоваться как ауксотрофией, так и дефектами регуляции одновременно. Например: у Corynebacterium glutamicum
- 17. L-глутаминовая кислота (α-аминоглутаровая): – первая аминокислота, полученная путем промышленного микробиологического синтеза; важнейшая аминокислота растительных и животных
- 18. Продуценты глутаминовой кислоты: - дрожжи, - микроскопические грибы, - бактерии (обеспечивают наибольший выход). Бактерии - продуценты:
- 19. Стадии производства глутаминовой кислоты : Приготовление питательной среды. - Питательная среда содержит 5 % сахарозы, 1
- 20. - Интенсивность аэрации – 40–45 мг О2 л/мин, рН 7,8–8,0, температура – 30 °С. - Ферментация
- 21. Постферментационная стадия (получение высокоочищенных препаратов) : 1. в культуральную жидкость добавляют: - негашеную известь или известковое
- 22. Для получения глутамата натрия - кристаллы глутаминовой кислоты обрабатывают гидроокисью натрия: 1. влажные кристаллы растворяют в
- 23. Лизин – алифатическая незаменимая аминокислота: - структурные элемент белка, - является предшественником карнитина и оксилизина, -
- 24. Лизин – алифатическая незаменимая аминокислота: - структурные элемент белка, - является предшественником карнитина и оксилизина, -
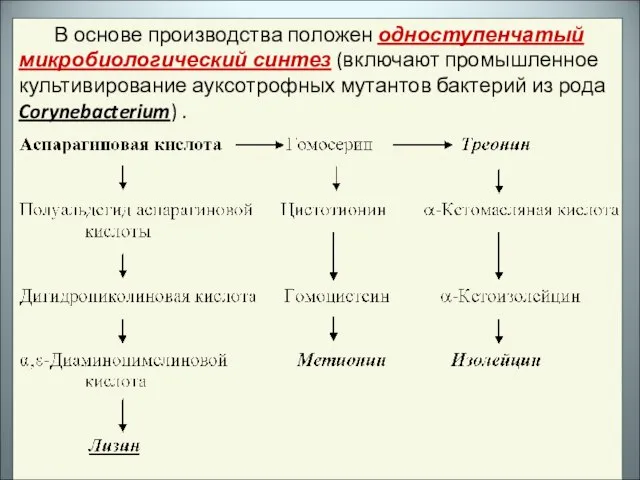
- 25. В основе производства положен одноступенчатый микробиологический синтез (включают промышленное культивирование ауксотрофных мутантов бактерий из рода Corynebacterium)
- 26. Мутантные клетки, не образующие гомосерин-дегидрогеназы, при культивировании на искусственной питательной среде обеспечивают высокий выход лизина. Дефицитные
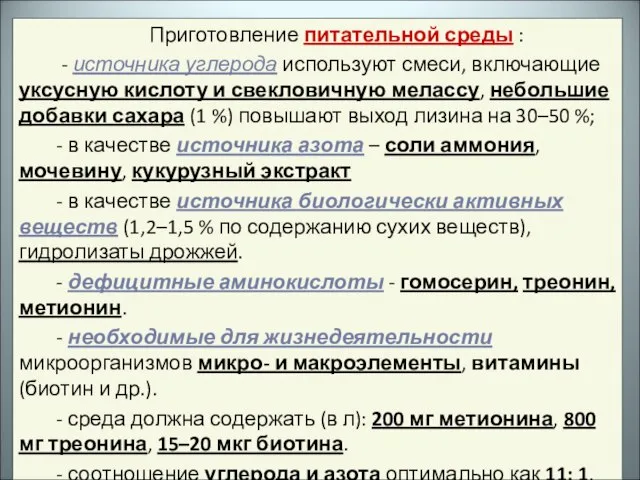
- 27. Приготовление питательной среды : - источника углерода используют смеси, включающие уксусную кислоту и свекловичную мелассу, небольшие
- 28. Посевной материал - вначале выращивается в посевных аппаратах при 28–32 °C, рН 7–7,2 в течение 18–24
- 29. Производство нескольких видов продукции: жидкий концентрат лизина (ЖКЛ), сухой кормовой концентрат лизина (ККЛ), высококонцентрированные кормовые и
- 30. 3. Для получения очищенного высококонцентрированного препарата лизина культуральную жидкость после фильтрования подкисляют соляной кислотой до рН
- 31. Отсутствие или дефицит триптофана в организме приводит к ряду тяжелых заболеваний (диабет, туберкулез, пеллагра). Для производства
- 33. Продуценты - ауксотрофные мутанты рода Bacillus subtilis с нарушенным синтезом фенилаланина и тирозина. Все технологические процессы
- 34. Биохимическое превращение антраниловой кислоты в триптофан происходит в 3 этапа: - на I этапе из антраниловой
- 36. Скачать презентацию
Слайд 2Сферы практического применения препаратов аминокислот
1) в пищевой промышленности (около 30 %
Сферы практического применения препаратов аминокислот
1) в пищевой промышленности (около 30 %

Слайд 3 3) в химической промышленности аминокислоты используют как предшественники :
в производстве
3) в химической промышленности аминокислоты используют как предшественники :
в производстве

Слайд 4Получение аминокислот гидролизом природного белоксодержащего сырья
В качестве источников для гидролиза используют:
отходы
Получение аминокислот гидролизом природного белоксодержащего сырья
В качестве источников для гидролиза используют:
отходы

Слайд 5Для выделения отдельных аминокислот из гидролизата проводят сложную многостадийную очистку, включающую в
Для выделения отдельных аминокислот из гидролизата проводят сложную многостадийную очистку, включающую в

Слайд 6Получение аминокислот химическим синтезом
Химический синтез аминокислот занимает второе место по объему
Получение аминокислот химическим синтезом
Химический синтез аминокислот занимает второе место по объему

Слайд 72. Производство аминокислот связано с использованием дорогостоящего оборудования и агрессивных токсических соединений

Слайд 8Микробиологический способ производства аминокислот
Более 60 % всех производимых чистых препаратов аминокислот
Микробиологический способ производства аминокислот
Более 60 % всех производимых чистых препаратов аминокислот

Слайд 9
При одноступенчатом синтезе в промышленных культиваторах выращивают ауксотрофные регуляторные мутанты, являющиеся
При одноступенчатом синтезе в промышленных культиваторах выращивают ауксотрофные регуляторные мутанты, являющиеся

Слайд 10
В процессе двухступенчатого синтеза аминокислоты:
получают предшественника аминокислоты (часто более
В процессе двухступенчатого синтеза аминокислоты:
получают предшественника аминокислоты (часто более

Слайд 11 Продуценты аминокислот.
В качестве продуцентов аминокислот используют генетически измененные штаммы, обладающие
Продуценты аминокислот.
В качестве продуцентов аминокислот используют генетически измененные штаммы, обладающие

Слайд 12 Для производства аминокислот бактерии стали использовать с начала 50-х гг., при
Для производства аминокислот бактерии стали использовать с начала 50-х гг., при

Слайд 13Условия накопления глутамата:
Полное или частичное подавление активности α-кетоглутаратдегидрогеназы,
Добавление β-лактамных антибиотиков,
Добавление
Условия накопления глутамата:
Полное или частичное подавление активности α-кетоглутаратдегидрогеназы,
Добавление β-лактамных антибиотиков,
Добавление

Слайд 14Ауксотрофные мутанты применяются, когда необходимо синтезировать соединения являющиеся конечными продуктами разветвленных цепей
Ауксотрофные мутанты применяются, когда необходимо синтезировать соединения являющиеся конечными продуктами разветвленных цепей

Слайд 15 Ауксотрофные мутанты способны накапливать конечные продукты неразветвленных путей биосинтеза.
В
Ауксотрофные мутанты способны накапливать конечные продукты неразветвленных путей биосинтеза.
В

Слайд 16 Для увеличения выходааминокислот можно воспользоваться как ауксотрофией, так и дефектами регуляции
Для увеличения выходааминокислот можно воспользоваться как ауксотрофией, так и дефектами регуляции

Слайд 17 L-глутаминовая кислота (α-аминоглутаровая):
– первая аминокислота, полученная путем промышленного микробиологического синтеза;
L-глутаминовая кислота (α-аминоглутаровая):
– первая аминокислота, полученная путем промышленного микробиологического синтеза;

Слайд 18 Продуценты глутаминовой кислоты:
- дрожжи,
- микроскопические грибы,
- бактерии (обеспечивают наибольший
Продуценты глутаминовой кислоты:
- дрожжи,
- микроскопические грибы,
- бактерии (обеспечивают наибольший

Слайд 19 Стадии производства глутаминовой кислоты :
Приготовление питательной среды.
- Питательная среда
Стадии производства глутаминовой кислоты :
Приготовление питательной среды.
- Питательная среда

Слайд 20
- Интенсивность аэрации – 40–45 мг О2 л/мин, рН 7,8–8,0,
- Интенсивность аэрации – 40–45 мг О2 л/мин, рН 7,8–8,0,

Слайд 21Постферментационная стадия
(получение высокоочищенных препаратов) :
1. в культуральную жидкость добавляют:
-
Постферментационная стадия
(получение высокоочищенных препаратов) :
1. в культуральную жидкость добавляют:
-

Слайд 22Для получения глутамата натрия -
кристаллы глутаминовой кислоты обрабатывают гидроокисью натрия:
1. влажные
Для получения глутамата натрия -
кристаллы глутаминовой кислоты обрабатывают гидроокисью натрия:
1. влажные

Слайд 23 Лизин – алифатическая незаменимая аминокислота:
- структурные элемент белка,
- является предшественником
Лизин – алифатическая незаменимая аминокислота:
- структурные элемент белка,
- является предшественником

Слайд 24 Лизин – алифатическая незаменимая аминокислота:
- структурные элемент белка,
- является предшественником
Лизин – алифатическая незаменимая аминокислота:
- структурные элемент белка,
- является предшественником

Слайд 25 В основе производства положен одноступенчатый микробиологический синтез (включают промышленное культивирование ауксотрофных
В основе производства положен одноступенчатый микробиологический синтез (включают промышленное культивирование ауксотрофных
Слайд 26 Мутантные клетки, не образующие гомосерин-дегидрогеназы, при культивировании на искусственной питательной среде
Мутантные клетки, не образующие гомосерин-дегидрогеназы, при культивировании на искусственной питательной среде

Слайд 27Приготовление питательной среды :
- источника углерода используют смеси, включающие уксусную кислоту
Приготовление питательной среды :
- источника углерода используют смеси, включающие уксусную кислоту
Слайд 28Посевной материал - вначале выращивается в посевных аппаратах при 28–32 °C, рН
Посевной материал - вначале выращивается в посевных аппаратах при 28–32 °C, рН

Слайд 29Производство нескольких видов продукции: жидкий концентрат лизина (ЖКЛ), сухой кормовой концентрат лизина
Производство нескольких видов продукции: жидкий концентрат лизина (ЖКЛ), сухой кормовой концентрат лизина

Слайд 303. Для получения очищенного высококонцентрированного препарата лизина культуральную жидкость после фильтрования подкисляют
3. Для получения очищенного высококонцентрированного препарата лизина культуральную жидкость после фильтрования подкисляют

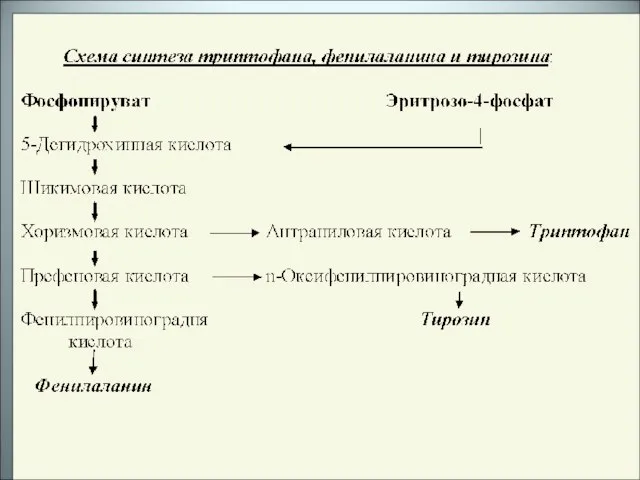
Слайд 31Отсутствие или дефицит триптофана в организме приводит к ряду тяжелых заболеваний (диабет,
Отсутствие или дефицит триптофана в организме приводит к ряду тяжелых заболеваний (диабет,

Слайд 33Продуценты - ауксотрофные мутанты рода Bacillus subtilis с нарушенным синтезом фенилаланина и
Продуценты - ауксотрофные мутанты рода Bacillus subtilis с нарушенным синтезом фенилаланина и

Слайд 34Биохимическое превращение антраниловой кислоты в триптофан происходит в 3 этапа:
- на
Биохимическое превращение антраниловой кислоты в триптофан происходит в 3 этапа:
- на

 Зачемсовременному человекууметь рисовать на компьютере?
Зачемсовременному человекууметь рисовать на компьютере? Собрание организаторов теневой экономики
Собрание организаторов теневой экономики ОАО "ЛВЗ "Висант"
ОАО "ЛВЗ "Висант" Интеллектуальная игра День Конституции
Интеллектуальная игра День Конституции Человек в политической жизни
Человек в политической жизни Презентация на тему Булгаков "Мастер и Маргарита". Понтий Пилат и Иешуа Га-Ноцри
Презентация на тему Булгаков "Мастер и Маргарита". Понтий Пилат и Иешуа Га-Ноцри  Решения Microsoft для образования
Решения Microsoft для образования Оформление букв
Оформление букв Х. Колумб – один из величайших мореплавателей
Х. Колумб – один из величайших мореплавателей Корни с чередованием
Корни с чередованием Рукодельная привычка. Игра-викторина
Рукодельная привычка. Игра-викторина Обязанности лиц суточного наряда
Обязанности лиц суточного наряда  Звёзды и созвездия
Звёзды и созвездия Презентация на тему Без праздника не бывает детства
Презентация на тему Без праздника не бывает детства Как стать офицером
Как стать офицером Качество речи 11 класс
Качество речи 11 класс RCR
RCR Урок развития речи. Описание картины Е.М.Широкова Друзья
Урок развития речи. Описание картины Е.М.Широкова Друзья Wonders of the World
Wonders of the World Основы химической кинетики
Основы химической кинетики Разложение разности квадратов на множители
Разложение разности квадратов на множители PureMVС в картинках - для ленивых – Ростислав Сирык http://flash-ripper.com/
PureMVС в картинках - для ленивых – Ростислав Сирык http://flash-ripper.com/ Снеговик
Снеговик Игра. Профессиональные стандарты в подготовке должностных лиц таможенных органов
Игра. Профессиональные стандарты в подготовке должностных лиц таможенных органов Стадии развития эмбриона
Стадии развития эмбриона Лихтенштейн
Лихтенштейн Презентация на тему Архитектура Персонального компьютера
Презентация на тему Архитектура Персонального компьютера  Aaaa
Aaaa