- Рост и развитие растений Основные фитогормоны

Содержание
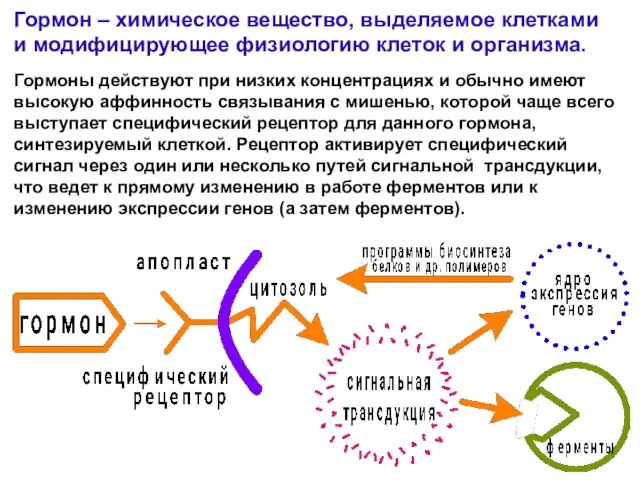
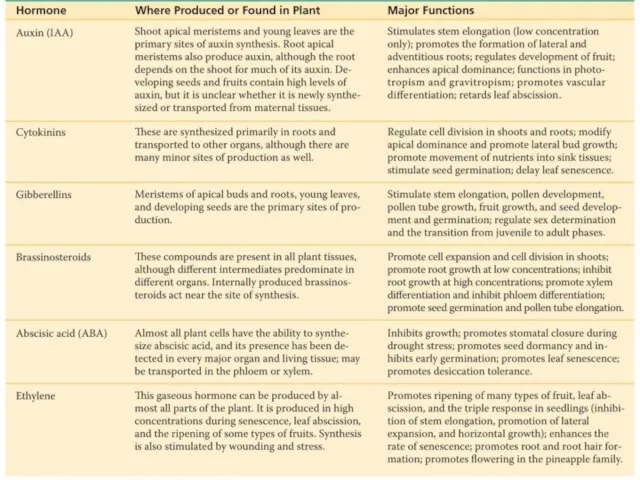
- 2. Гормон – химическое вещество, выделяемое клетками и модифицирующее физиологию клеток и организма. Гормоны действуют при низких
- 3. Историю открытия и изучения фитогормонов – по учебнику! В 19 веке, Чарльз Дарвин и его сын
- 4. Результаты Контроль Свет Освещенная сторона колеоптиля Затененная сторона колеоптиля
- 5. Результаты (конец 19-го века) Свет Кончик удален Дарвин и Дарвин: фототропический ответ возникает только когда кончик
- 6. Свет Кончик отделен желатиновой септой, которая проницаема для растворов Кончик отделен непроницаемой пластинкой 1913 год, Питер
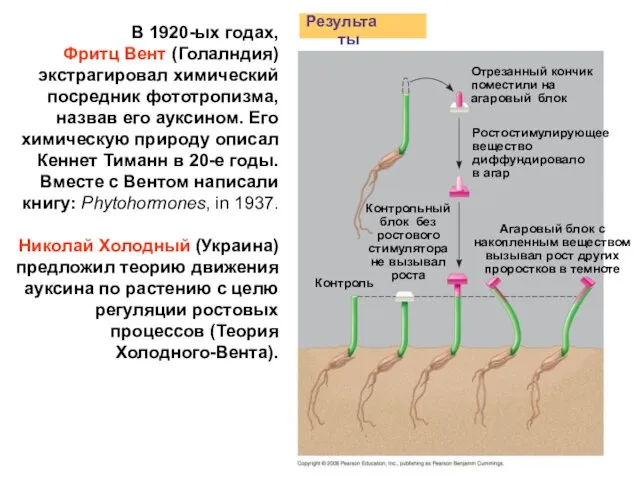
- 7. В 1920-ых годах, Фритц Вент (Голалндия) экстрагировал химический посредник фототропизма, назвав его ауксином. Его химическую природу
- 9. Ауксин (греч. auxein - «расти/увеличиваться») Любой химический агент, стимулитующий растяжение колеоптилей (классификация по физиологической активности!). Индолил-3-уксусная
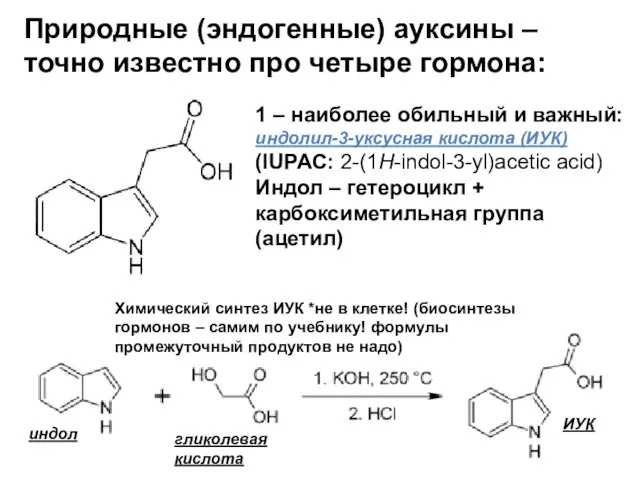
- 10. Природные (эндогенные) ауксины – точно известно про четыре гормона: 1 – наиболее обильный и важный: индолил-3-уксусная
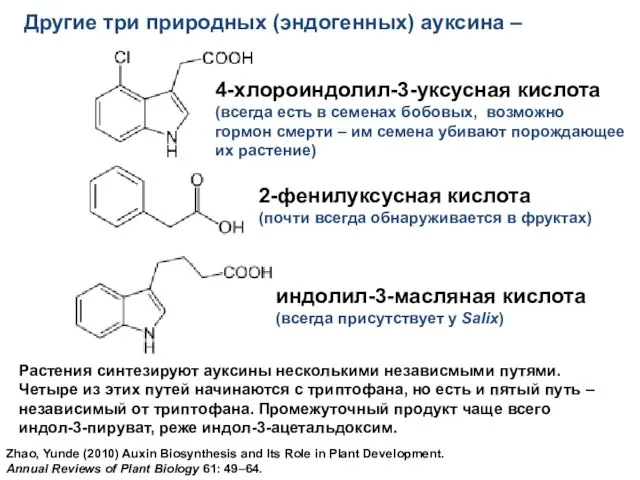
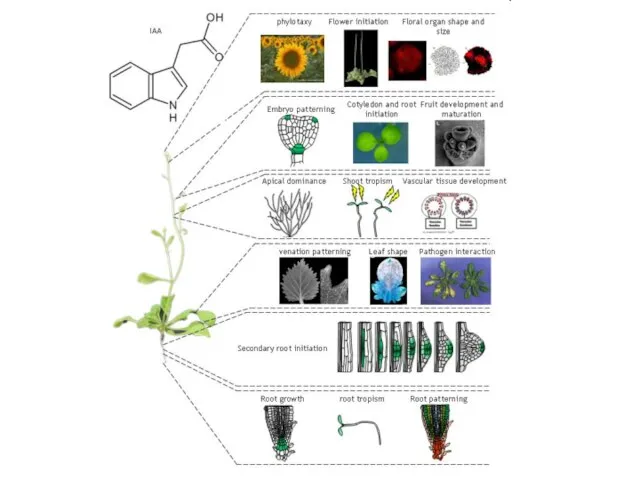
- 11. Другие три природных (эндогенных) ауксина – Растения синтезируют ауксины несколькими независмыми путями. Четыре из этих путей
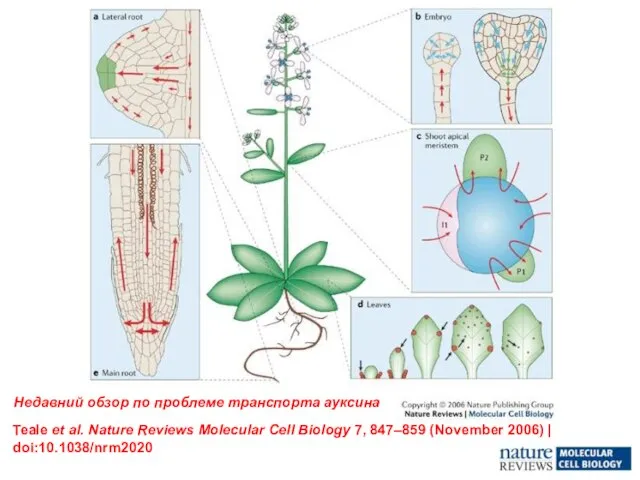
- 13. Teale et al. Nature Reviews Molecular Cell Biology 7, 847–859 (November 2006) | doi:10.1038/nrm2020 Недавний обзор
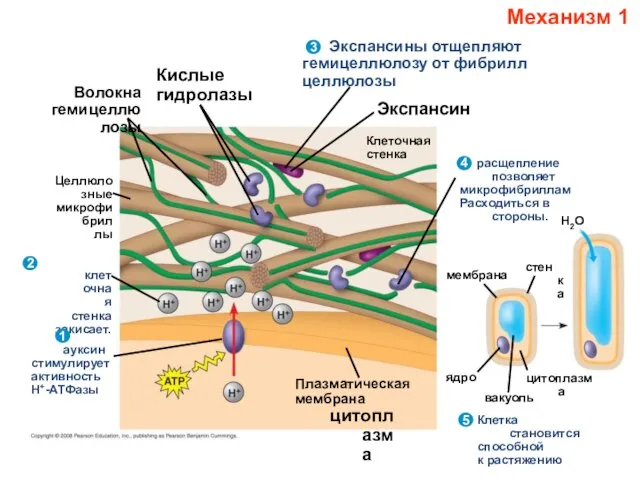
- 15. Как ауксины стимулируют рост? - Н+-АТФазы, - экспансины Ауксин стимулирует протонную АТФазу; выделяемые этим ферментом Н+
- 16. Волокна гемицеллюлозы Целлюлозные микрофибриллы клеточная стенка закисает. 2 1 ауксин стимулирует активность Н+-АТФазы Кислые гидролазы Экспансин
- 17. Уже для вас известный механизм (механизм 2): 1 - H+-ATФаза гиперполяризует мембрану, выкачивая положительно заряженные H+
- 18. Механизм 2
- 19. Цитокинины стимулируют цитокинез(ис) – деление клеток Зеатин (первый описанный цитокинин из кукурузы) Контролируют клеточное деление и
- 20. Цитокинины, ауксин и ряд других факторов взаимодействуют для контроля апикального доминирования. В апик. домин. терминальная (верхушечная)
- 21. Можно выделить 2 типа цитокининов по их структуре: 1 – адениновый тип: кинетин, зеатин и 6-бензиламинопурин
- 22. Гиббереллины (классификация по общей структуре!) также как и все гормоны имеют плеотропное действие: - Удлинение побегов
- 23. Гиббереллины Гиббереловая кислота У растений, грибов и бактерий обнаружено около 150 веществ, относимых к группе гиббереллинов.
- 24. ГК - открыта японским ученым Ейчи Куросавой в 1926 г. Как продукт патогенного гриба Gibberella fujikuroi,
- 25. Ауксин и гиббереллины регулируют созревание фруктов и закладку семян. Gibberellins are used in spraying of Thompson
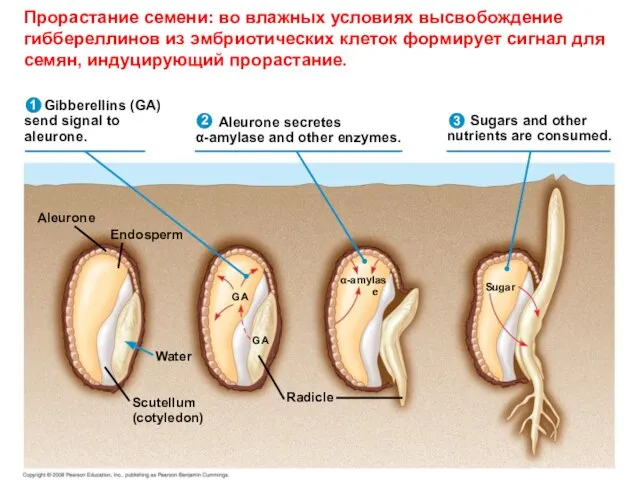
- 26. Gibberellins (GA) send signal to aleurone. Aleurone secretes α-amylase and other enzymes. Sugars and other nutrients
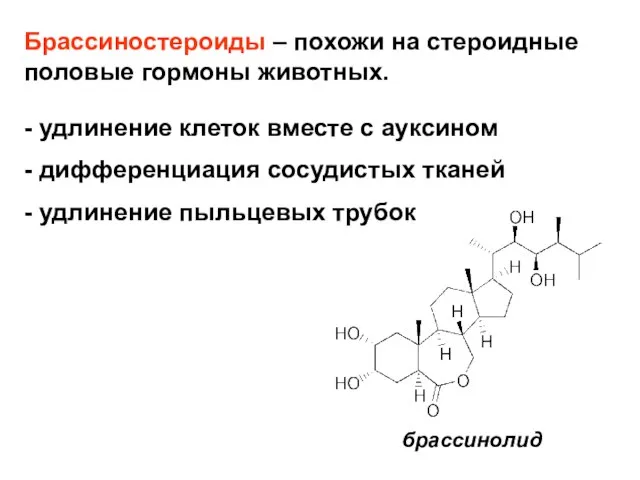
- 27. Брассиностероиды – похожи на стероидные половые гормоны животных. - удлинение клеток вместе с ауксином - дифференциация
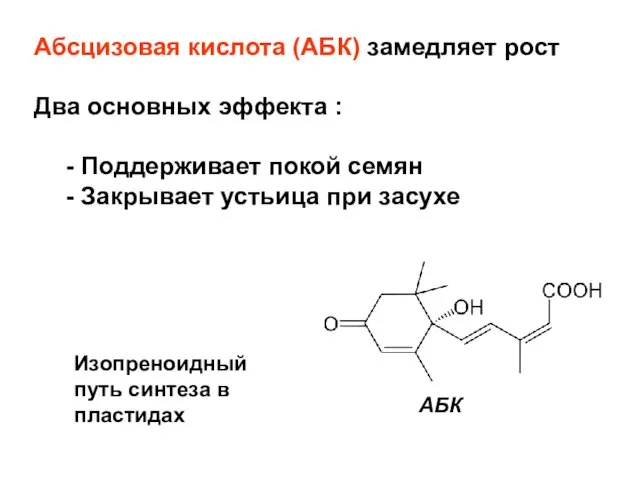
- 28. Абсцизовая кислота (AБК) замедляет рост Два основных эффекта : - Поддерживает покой семян - Закрывает устьица
- 29. Early germination in red mangrove Early germination in maize mutant Coleoptile
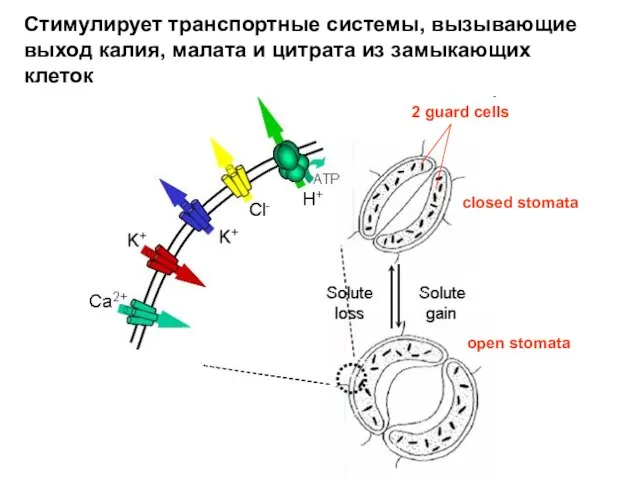
- 30. Стимулирует транспортные системы, вызывающие выход калия, малата и цитрата из замыкающих клеток closed stomata open stomata
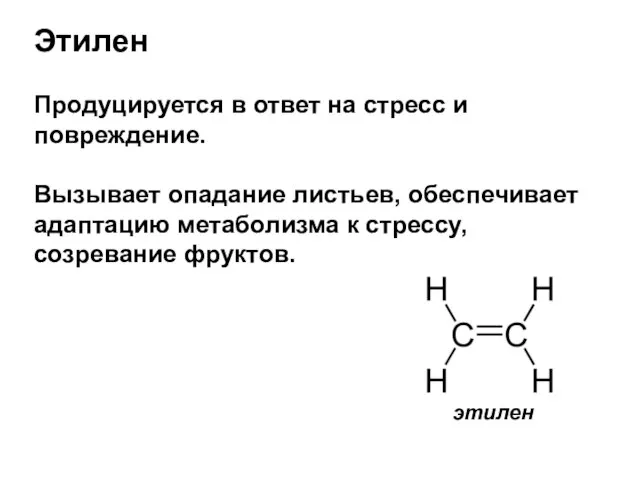
- 31. Этилен Продуцируется в ответ на стресс и повреждение. Вызывает опадание листьев, обеспечивает адаптацию метаболизма к стрессу,
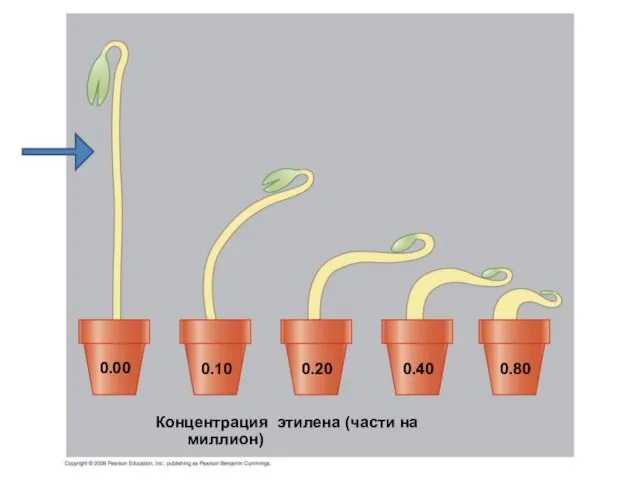
- 32. Концентрация этилена (части на миллион) 0.10 0.00 0.20 0.40 0.80
- 33. Защитынй слой Стебель Слой отделения Черешок Уменьшение уровня ауксина и повышение уровня этилена приводит к образованию
- 34. Учить не надо
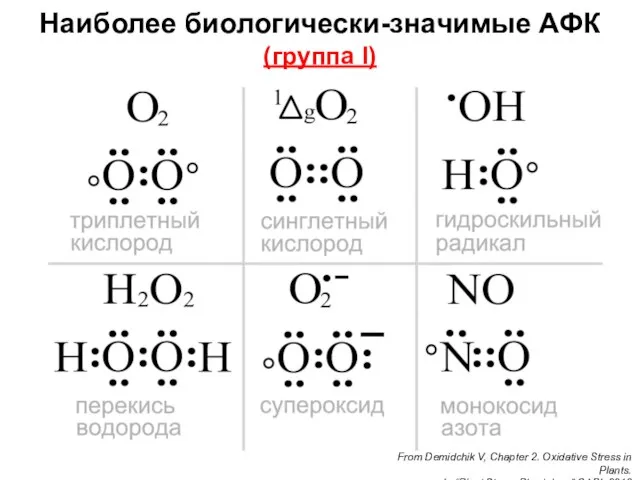
- 35. Наиболее биологически-значимые АФК (группа I) From Demidchik V, Chapter 2. Oxidative Stress in Plants. In “Plant
- 36. Другие АФК, участвующие в оксидативном стрессе и физиологических реакциях (группа II) Гидропероксильный (гидропероксидный) радикал HO2• Пероскильный
- 37. Потенциальные рецепторы АФК в клетках растений: (A) двухкомпонентные гистидин-киназы (two-component histidine kinases); (Б) редокс-чувствительные факторы транскрипции:
- 38. Почему так важен Са2+? - повышение уровня Са2+ в цитоплазме (4-мерная кальциевая волна - XYZt) –
- 39. Цитоплазматические сенсоры Са2+: Кальций-связывающие белки “Ca2+-binding proteins” (CBPs). Примерно 250 генов на геном (высшие растения). «Обратимо
- 40. Другие сигнально-регуляторные механизмы, индуцируемые АФК: MAP-киназы и другие серин-треониновые киназы, MAPK-фосфатазы и другие ферменты активируются под
- 42. Скачать презентацию
Слайд 2Гормон – химическое вещество, выделяемое клетками и модифицирующее физиологию клеток и организма.
Гормоны
Гормон – химическое вещество, выделяемое клетками и модифицирующее физиологию клеток и организма.
Гормоны
Слайд 3Историю открытия и изучения фитогормонов – по учебнику!
В 19 веке, Чарльз Дарвин
Историю открытия и изучения фитогормонов – по учебнику!
В 19 веке, Чарльз Дарвин

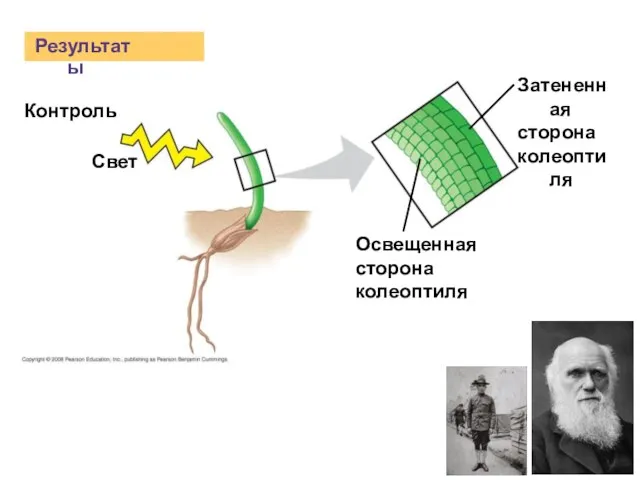
Слайд 4Результаты
Контроль
Свет
Освещенная
сторона
колеоптиля
Затененная
сторона
колеоптиля
Результаты
Контроль
Свет
Освещенная
сторона
колеоптиля
Затененная
сторона
колеоптиля
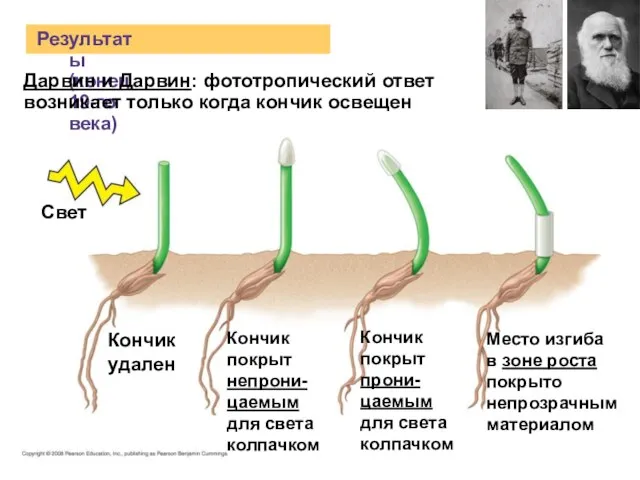
Слайд 5Результаты (конец 19-го века)
Свет
Кончик
удален
Дарвин и Дарвин: фототропический ответ
возникает только когда кончик
Результаты (конец 19-го века)
Свет
Кончик
удален
Дарвин и Дарвин: фототропический ответ
возникает только когда кончик
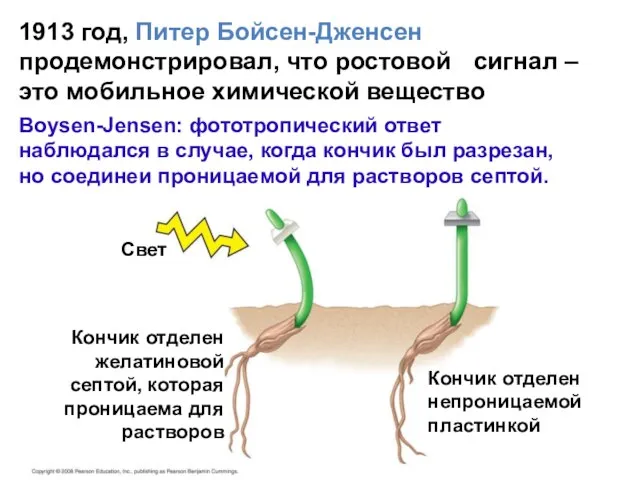
Слайд 6Свет
Кончик отделен
желатиновой
септой, которая
проницаема для
растворов
Кончик отделен
непроницаемой
пластинкой
1913 год,
Свет
Кончик отделен
желатиновой
септой, которая
проницаема для
растворов
Кончик отделен
непроницаемой
пластинкой
1913 год,
Слайд 7В 1920-ых годах,
Фритц Вент (Голалндия) экстрагировал химический посредник фототропизма, назвав его
В 1920-ых годах,
Фритц Вент (Голалндия) экстрагировал химический посредник фототропизма, назвав его
Слайд 9Ауксин (греч. auxein - «расти/увеличиваться»)
Любой химический агент, стимулитующий растяжение колеоптилей (классификация по физиологической
Ауксин (греч. auxein - «расти/увеличиваться»)
Любой химический агент, стимулитующий растяжение колеоптилей (классификация по физиологической

Слайд 10Природные (эндогенные) ауксины – точно известно про четыре гормона:
1 – наиболее обильный
Природные (эндогенные) ауксины – точно известно про четыре гормона:
1 – наиболее обильный
Слайд 11Другие три природных (эндогенных) ауксина –
Растения синтезируют ауксины несколькими независмыми путями.
Другие три природных (эндогенных) ауксина –
Растения синтезируют ауксины несколькими независмыми путями.
Слайд 13Teale et al. Nature Reviews Molecular Cell Biology 7, 847–859 (November 2006)
Teale et al. Nature Reviews Molecular Cell Biology 7, 847–859 (November 2006)
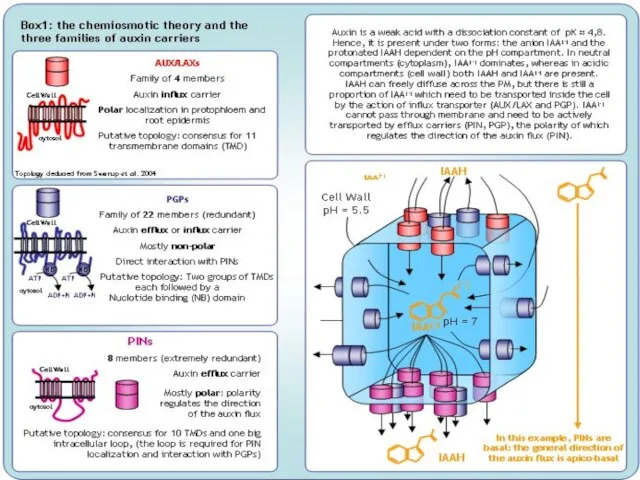
Слайд 15Как ауксины стимулируют рост?
- Н+-АТФазы, - экспансины
Ауксин стимулирует протонную АТФазу; выделяемые этим
Как ауксины стимулируют рост?
- Н+-АТФазы, - экспансины
Ауксин стимулирует протонную АТФазу; выделяемые этим

Слайд 16Волокна
гемицеллюлозы
Целлюлозные
микрофибриллы
клеточная
стенка
закисает.
2
1
ауксин
стимулирует
активность
Н+-АТФазы
Кислые
гидролазы
Экспансин
Экспансины
Волокна
гемицеллюлозы
Целлюлозные
микрофибриллы
клеточная
стенка
закисает.
2
1
ауксин
стимулирует
активность
Н+-АТФазы
Кислые
гидролазы
Экспансин
Экспансины
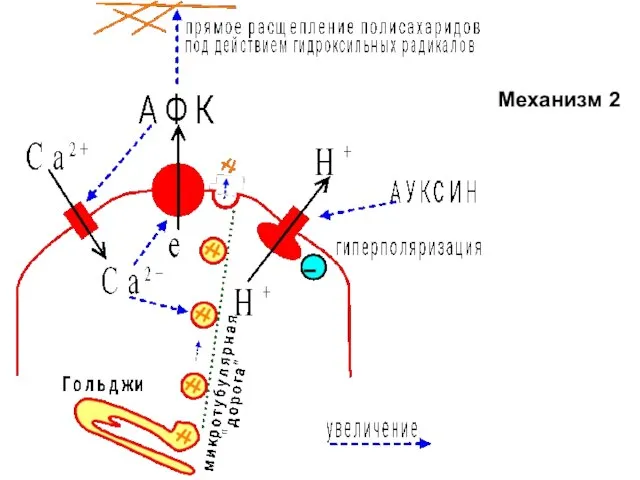
Слайд 17Уже для вас известный механизм (механизм 2):
1 - H+-ATФаза гиперполяризует мембрану,
Уже для вас известный механизм (механизм 2):
1 - H+-ATФаза гиперполяризует мембрану,

Слайд 18Механизм 2
Механизм 2
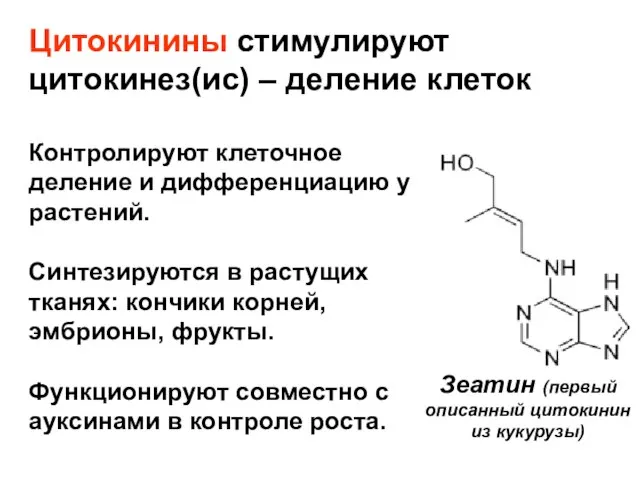
Слайд 19Цитокинины стимулируют цитокинез(ис) – деление клеток
Зеатин (первый описанный цитокинин из кукурузы)
Контролируют клеточное
Цитокинины стимулируют цитокинез(ис) – деление клеток
Зеатин (первый описанный цитокинин из кукурузы)
Контролируют клеточное
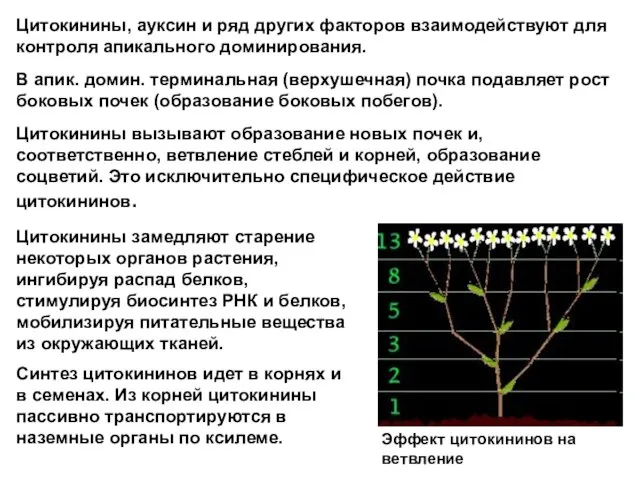
Слайд 20Цитокинины, ауксин и ряд других факторов взаимодействуют для контроля апикального доминирования.
В апик.
Цитокинины, ауксин и ряд других факторов взаимодействуют для контроля апикального доминирования.
В апик.
Слайд 21Можно выделить 2 типа цитокининов по их структуре:
1 – адениновый тип: кинетин,
Можно выделить 2 типа цитокининов по их структуре:
1 – адениновый тип: кинетин,

Слайд 22Гиббереллины (классификация по общей структуре!) также как и все гормоны имеют плеотропное
Гиббереллины (классификация по общей структуре!) также как и все гормоны имеют плеотропное
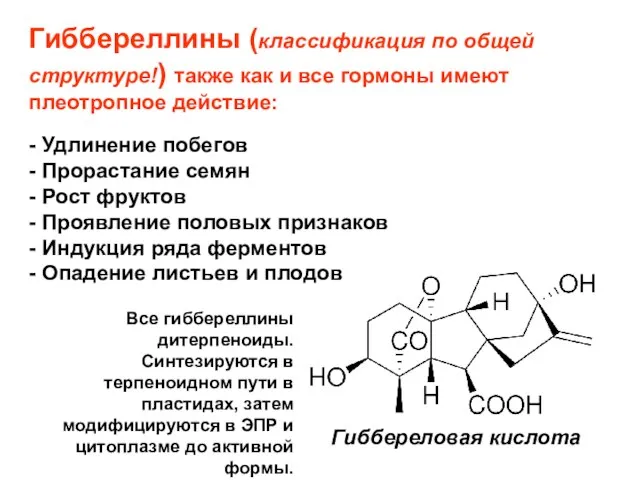
Слайд 23Гиббереллины
Гиббереловая кислота
У растений, грибов и бактерий обнаружено около 150 веществ, относимых к
Гиббереллины
Гиббереловая кислота
У растений, грибов и бактерий обнаружено около 150 веществ, относимых к

Слайд 24ГК - открыта японским ученым Ейчи Куросавой в 1926 г. Как продукт
ГК - открыта японским ученым Ейчи Куросавой в 1926 г. Как продукт

Слайд 25Ауксин и гиббереллины регулируют созревание фруктов и закладку семян. Gibberellins are used
Ауксин и гиббереллины регулируют созревание фруктов и закладку семян. Gibberellins are used

Слайд 26 Gibberellins (GA)
send signal to
aleurone.
Aleurone secretes
α-amylase and other enzymes.
Sugars
Gibberellins (GA)
send signal to
aleurone.
Aleurone secretes
α-amylase and other enzymes.
Sugars
Слайд 27Брассиностероиды – похожи на стероидные половые гормоны животных.
- удлинение клеток вместе
Брассиностероиды – похожи на стероидные половые гормоны животных.
- удлинение клеток вместе
Слайд 28Абсцизовая кислота (AБК) замедляет рост
Два основных эффекта :
- Поддерживает покой семян
- Закрывает
Абсцизовая кислота (AБК) замедляет рост
Два основных эффекта :
- Поддерживает покой семян
- Закрывает
Слайд 29Early germination
in red mangrove
Early germination
in maize mutant
Coleoptile
Early germination
in red mangrove
Early germination
in maize mutant
Coleoptile

Слайд 30Стимулирует транспортные системы, вызывающие выход калия, малата и цитрата из замыкающих клеток
Стимулирует транспортные системы, вызывающие выход калия, малата и цитрата из замыкающих клеток
Слайд 31Этилен
Продуцируется в ответ на стресс и повреждение.
Вызывает опадание листьев, обеспечивает адаптацию метаболизма
Этилен
Продуцируется в ответ на стресс и повреждение.
Вызывает опадание листьев, обеспечивает адаптацию метаболизма
Слайд 32Концентрация этилена (части на миллион)
0.10
0.00
0.20
0.40
0.80
Концентрация этилена (части на миллион)
0.10
0.00
0.20
0.40
0.80
Слайд 33Защитынй слой
Стебель
Слой отделения
Черешок
Уменьшение уровня ауксина и повышение уровня этилена приводит к образованию
Защитынй слой
Стебель
Слой отделения
Черешок
Уменьшение уровня ауксина и повышение уровня этилена приводит к образованию

Слайд 34Учить не надо
Учить не надо
Слайд 35Наиболее биологически-значимые АФК (группа I)
From Demidchik V, Chapter 2. Oxidative Stress in
Наиболее биологически-значимые АФК (группа I)
From Demidchik V, Chapter 2. Oxidative Stress in
Слайд 36Другие АФК, участвующие в оксидативном стрессе и физиологических реакциях (группа II)
Гидропероксильный (гидропероксидный)
Другие АФК, участвующие в оксидативном стрессе и физиологических реакциях (группа II)
Гидропероксильный (гидропероксидный)

Слайд 37Потенциальные рецепторы АФК в клетках растений:
(A) двухкомпонентные гистидин-киназы (two-component histidine kinases);
Потенциальные рецепторы АФК в клетках растений:
(A) двухкомпонентные гистидин-киназы (two-component histidine kinases);

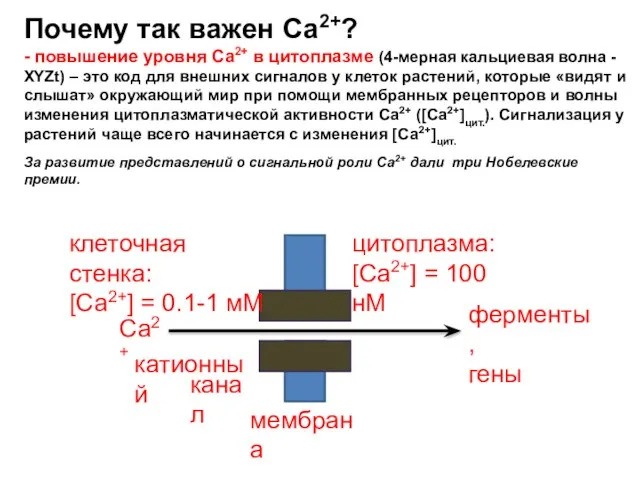
Слайд 38Почему так важен Са2+?
- повышение уровня Са2+ в цитоплазме (4-мерная кальциевая волна
Почему так важен Са2+?
- повышение уровня Са2+ в цитоплазме (4-мерная кальциевая волна
Слайд 39Цитоплазматические сенсоры Са2+:
Кальций-связывающие белки “Ca2+-binding proteins” (CBPs). Примерно 250 генов на
Цитоплазматические сенсоры Са2+:
Кальций-связывающие белки “Ca2+-binding proteins” (CBPs). Примерно 250 генов на

Слайд 40Другие сигнально-регуляторные механизмы, индуцируемые АФК:
MAP-киназы и другие серин-треониновые киназы, MAPK-фосфатазы и
Другие сигнально-регуляторные механизмы, индуцируемые АФК:
MAP-киназы и другие серин-треониновые киназы, MAPK-фосфатазы и

 Сочитание обуви и брюк
Сочитание обуви и брюк Образ одуванчика средствами живописи (ИЗО, 1 класс)
Образ одуванчика средствами живописи (ИЗО, 1 класс) Работа ПМПк МБДОУ детского сада общеразвивающего вида с приорететным осущетсвлением деятельности по физическому развитию детей
Работа ПМПк МБДОУ детского сада общеразвивающего вида с приорететным осущетсвлением деятельности по физическому развитию детей Past_Tenses
Past_Tenses Главный секретарь
Главный секретарь КИБЕРНЕТИКА
КИБЕРНЕТИКА Ёлку украшаем. Ёлкым сорастарена
Ёлку украшаем. Ёлкым сорастарена Инновационные подходы в организации событийных мероприятий в учреждениях культуры
Инновационные подходы в организации событийных мероприятий в учреждениях культуры Презентация на тему Русская литература второй половины 19 века
Презентация на тему Русская литература второй половины 19 века Комнатные растения в интерьере 7 класс
Комнатные растения в интерьере 7 класс Алтайский край. Получение выплаты от 3 до 7 лет в новых условиях
Алтайский край. Получение выплаты от 3 до 7 лет в новых условиях Словесная живопись рассказов М.А.Шолохова.
Словесная живопись рассказов М.А.Шолохова. Bonjour mes amis. 5 класс
Bonjour mes amis. 5 класс Пуантилизм. Открытый урок. Изобразительное искусство. 6 класс
Пуантилизм. Открытый урок. Изобразительное искусство. 6 класс St. Valentine's Day
St. Valentine's Day edf82d9d26bc44b09aecd98f1a92e47e
edf82d9d26bc44b09aecd98f1a92e47e Эконива. Прописные истины рекламы и маркетинга
Эконива. Прописные истины рекламы и маркетинга Федеральный государственный образовательный стандарт начального общего образования по литературному чтению
Федеральный государственный образовательный стандарт начального общего образования по литературному чтению резервное копирование и восстановление данных
резервное копирование и восстановление данных Мышцы
Мышцы Системы счисления, история и современность
Системы счисления, история и современность Деятельность школы как центра творческого развития личности школьника в пространстве дополнительного образования
Деятельность школы как центра творческого развития личности школьника в пространстве дополнительного образования Неклеточные формы жизни. Вирусы и бактериофаги
Неклеточные формы жизни. Вирусы и бактериофаги Автоматы и пулеметы
Автоматы и пулеметы Правоотношения по пенсионному обеспечению
Правоотношения по пенсионному обеспечению Презентация на тему Проверка безударной гласной в корне слова
Презентация на тему Проверка безударной гласной в корне слова  Концепция бенчмаркинга
Концепция бенчмаркинга «Мониторинг социально-трудовой сферы. Результаты социологического исследования рынка труда»
«Мониторинг социально-трудовой сферы. Результаты социологического исследования рынка труда»